
Abstract
Background:
The major confirmed genetic risk factor for late-onset, sporadic Alzheimer’s disease (AD) is variant ɛ4 of apolipoprotein E gene (APOE). It is proposed that ApoE, a protein involved in transport of cholesterol to neurons can cause neurodegeneration in AD through interaction with metals. Previous studies mostly associated copper, iron, zinc, and calcium with ApoE4-mediated toxicity.
Objective:
To test the association of essential metals with APOE genotype.
Methods:
We compared plasma and cerebrospinal fluid (CSF) levels of copper, zinc, iron, sodium, magnesium, calcium, cobalt, molybdenum, manganese, boron, and chromium, and CSF ferritin levels among AD, mild cognitive impairment (MCI) patients, and healthy controls (HC) with different APOE genotype.
Results:
Sodium, copper, and magnesium levels were increased in carriers of ɛ4 allele. Additionally, the increase in sodium, calcium and cobalt plasma levels was observed in carriers of ɛ4/ɛx genotype. The decrease in boron plasma levels was observed in carriers of ɛ4 allele and ɛ4/ɛ4 genotype. Additionally, CSF zinc levels as well as plasma sodium levels were increased in AD patients compared to HC.
Conclusion:
These results indicate that the molecular underpinnings of association of essential metals and metalloids with APOE should be further tested and clarified in vivo and in vitro.
INTRODUCTION
Homeostasis of essential metals is altered in Alzheimer’s disease (AD) [1–3], and it has been proposed that such changes are directly related to AD pathology [1]. This primarily refers to essential metals that are normally present in organism and are crucial for normal functioning of many proteins and enzymes. Altered metal homeostasis in the AD brain has mainly been related to copper, zinc, and iron [1].
Apolipoprotein E (ApoE) is a protein involved in transport of cholesterol to neurons, and is mainly produced by astrocytes [4]. The apolipoprotein E gene (APOE) variant ɛ4 is the major confirmed genetic risk factor for late-onset, sporadic AD that comprises over 99%of all AD cases [5, 6]. There are three common ApoE variants (ApoE2, ApoE3, and ApoE4). Two single nucleotide polymorphisms (SNPs) present in APOE gene determine which amino acid will be present at the protein level at positions 112 and 158. Thus, ApoE2 variant has Cys112 and Cys158, ApoE3 has Cys112 and Arg158, and ApoE4 has Arg112 and Arg158 [7]. APOE ɛ4 heterozygotes have a 5 times increased risk, while APOE ɛ4 homozygotes have a 20 times increased risk for developing AD [8]. ApoE2 is considered to have protective effect in AD [9]. Association of metals with ApoE was observed in AD. Xu and collaborators proposed three mechanisms through which metal ions may interact with ApoE: 1) copper, zinc, and iron accumulate in amyloid plaques (AP); AP cause metal dyshomeostasis that leads to decrease in ApoE levels in AD; 2) metal dyshomeostasis in AD decreases APOE transcription and translation, which may promote toxicity of amyloid-β (Aβ) as ApoE promotes Aβ clearance; and 3) ApoE proteolysis that occurs in AD is more prominent for ApoE4 isoforms whose fragments disrupt mitochondrial and cytoskeletal functions and lead to neurodegeneration [10]. Stability of ApoE isoforms could be mediated by metal binding [11] and as metals stabilize ApoE isoforms in the order ɛ2 > ɛ3 > ɛ4, this could be the cause of higher vulnerability of the ApoE4 isoform for proteolysis. However, as these authors emphasized, these assumptions should be further tested [10].
Previous studies associated copper, iron, zinc, and calcium with ApoE4-mediated toxicity [12–15]. In the current study we further analyzed the association of other essential metals like sodium, magnesium, cobalt, molybdenum, manganese, chromium, and the metalloid boron with ApoE. We also measured cerebrospinal fluid (CSF) ferritin levels as it likely reflects the levels of iron in the brain. The scope of this study was to compare plasma and CSF levels of Cu, Zn, Fe, Na, Mg, Ca, Co, Mo, Mn, B, and Cr, and CSF levels of ferritin in AD, mild cognitive impairment (MCI) patients and healthy controls (HC) with different APOE genotype. The investigation of a possible association between essential metals and APOE permitted us to assess the existence of altered metal homeostasis in AD.
MATERIALS AND METHODS
Cerebrospinal fluid and blood collection
This study included 197 patients recruited at the University Hospital Centre, Zagreb and General Hospital Varaždin of whom 126 fulfilled NINCDS-ADRDA criteria for AD, 52 suffered from MCI [16, 17], and 19 were HC. Patients were neurologically tested, as described previously [18]. The examination included the Mini-Mental State Examination (MMSE), VDRL testing for syphilis, complete blood tests including thyroid function, albumin levels, and levels of vitamin B12 and electrolytes. None of the subjects included in this study suffered from renal diseases. It should be however noted that a limitation of this study is that although information on medication regimens was available for the majority of the included patients (summarized in Supplementary Table 1), it did not include information on use of supplements (as the levels of boron, copper, and magnesium may be affected by supplements). CSF was obtained between the L3/L4 or L4/L5 intervertebral spaces by lumbar puncture, always performed in the morning between 9 a.m. and 11 a.m. After centrifugation for 10 min at 2000 g, samples were aliquoted and stored at –80°C. Venous blood samples (4 ml) were collected into plastic syringes with 1 ml of acid citrate dextrose as an anticoagulant. Blood samples were consistently obtained in the morning on an empty stomach. Thrombocyte-free plasma samples were collected by centrifugation (1,100 g for 3 min and then 5,087 g for 15 min), and stored at –20°C. Ferritin levels in CSF were determined by electrochemiluminescence (ECL) using a Roche Cobas E601 instrument (Roche, Basel, Switzerland). All procedures were implemented in accordance with the approval of the Central Ethical Committee of the University of Zagreb Medical School (case no. 380-59-10106-18-111/126, class 641-01/18-02/01 from June 20, 2018) and Ethical Committee of the Clinical Hospital Centre Zagreb (case no. 02/21 AG, class 8.1-18/82-2 from April 24, 2018).
Genotyping
Genomic DNA was extracted from peripheral blood using the salting-out method [19]. APOE polymorphisms (rs7412 and rs429358) were determined in 122 AD and 52 MCI patients and 15 HC by ABI Prism 7300 Real Time PCR System apparatus (Applied Biosystems, Foster City, CA, USA) using primers and probes purchased from Applied Biosystems as TaqMan® SNP Genotyping Assay (C_904973_10 ND C_3084793_20). All genotyping procedures were done by a researcher who was blind to all clinical data according to the procedures described by Applied Biosystems. Out of 189, 54 samples (29%) were genotyped again as a quality control for genotyping analyses. The three common variants of APOE (ɛ2, ɛ3, and ɛ4) were determined by two SNPs (rs429358 and rs7412).
Analysis of metals by inductively coupled plasma mass spectroscopy (ICP-MS)
CSF and plasma levels of Cu, Zn, Fe, Na, Mg, Ca, Co, Mo, Mn, B, and Cr were determined using inductively coupled plasma mass spectroscopy (ICP-MS) on Agilent 7500cx (Agilent Technologies, Tokyo, Japan) (Table 1). Before analysis CSF and plasma samples were prepared by dilution (1:10 for CSF and 1:20 for plasma) with solution containing 0.7 mM ammonia, 0.01 mM EDTA, 0.07%(v/v) Triton X-100 and 2μg/l of internal standards (Ge, Rh, Tb, Lu, and Ir) in ultrapure water. MicroMist nebulizer combined with a Peltier standard quartz spray chamber (Scott–type) cooled at 2°C and a quartz torch with a 2.5 mm diameter injector with a Shield Plate system and Ni sampler and skimmer cones were used. Tune solution of 1μg/l 7Li, 59Co, 89Y, 140Ce, and 205Tl was used for daily optimization of ICP-MS working conditions. Samples preparation and analysis were done in a laboratory with HVAC system (Heating, Ventilating, and Air Conditioning) combined with HEPA filters. Standard addition method (i.e., matrix-matched calibration) was used for the quantification of elements concentration in CSF and plasma samples. To confirm the accuracy of the measurements, commercially available reference materials were used: ClinChek Plasma Controls (Level I and II) and ClinChek Serum Controls (Level I and II) from RECIPE (Munich, Germany); Seronorm Trace Elements Serum (Level I and II) (Sero AS, Billingstad, Norway). Since there is a possibility for contamination of the samples with chromium from the needles used for sample collection, chromium was removed from statistical analysis.
Levels of Cu, Zn, Fe, Na, Mg, Ca, Co, Mo, Mn, and B measured in CSF and plasma of AD, MCI patients, and HC
AD, Alzheimer’s disease; CSF, cerebrospinal fluid; HC, healthy control; MCI, mild cognitive impairment; SD, standard deviation.
Statistical analysis
CSF and plasma Cu, Zn, Fe, Na, Mg, Ca, Co, Mo, Mn, and B levels were compared between two groups using Mann-Whitney U tests, while non-parametric Kruskal-Wallis tests were used for comparison across three or more groups. A post-hoc non-parametric Dunn test to correct p values was used for pairwise comparisons. Statistical analysis was also done after introduction of age and sex as covariates. When analyzing sodium plasma and CSF levels, presence of hypertension and cardiovascular diseases were also introduced as additional covariates (Table 2). Covariate analysis was performed using non-parametric Quade’s ANCOVA. The genotype and allele distributions were determined by a χ2 test. Statistical analyses were performed with SPSS 19.0.1 (SPSS, Chicago, IL, USA), with the α value set at 0.05 for statistical significance.
Demographic data and information on the presence of hypertension and cardiovascular diseases in AD and MCI patients and HC
AD, Alzheimer’s disease; F, female; HC, healthy controls; M, male; MCI, mild cognitive impairment.
RESULTS
Levels of metals in plasma and CSF of AD, MCI patients, and HC
Significant difference in the levels of zinc measured in CSF was observed among AD, MCI patients, and HC (H test = 9.317, df = 2, p = 0.009; Fig. 1A). This association was preserved after correction for age and sex (F(2,190) = 3.766, p = 0.025). The levels of zinc were significantly increased in CSF of AD patients compared to HC (p = 0.027). The levels of sodium measured in plasma were significantly different between AD, MCI patients, and HC (H test = 10.567, df = 2, p = 0.005; Fig. 1B). Sodium levels were significantly increased in plasma of AD patients compared to HC (p = 0.004). This association was preserved after correction for age and sex (F(2,141) = 6.228, p = 0.003). When introducing hypertension in addition to age and sex as a covariate, the levels of sodium plasma levels remained significantly different among the groups (F(2,113) = 4.724, p = 0.011). However, when introducing existence of cardiovascular diseases as covariate in addition to age and sex, the significance in sodium plasma levels among the groups was lost (F(2,110) = 2.384, p = 0.097). Also, introduction of hypertension and cardiovascular diseases as covariates in addition to age and sex, resulted in loss of significance in plasma sodium levels among the groups (F(2,110) = 2.288, p = 0.106). There was no significant difference in the levels of zinc measured in plasma, sodium measured in CSF, and copper, iron, calcium, magnesium, cobalt, molybdenum, manganese and boron measured in both plasma and CSF among AD, MCI patients and HC (Table 1).
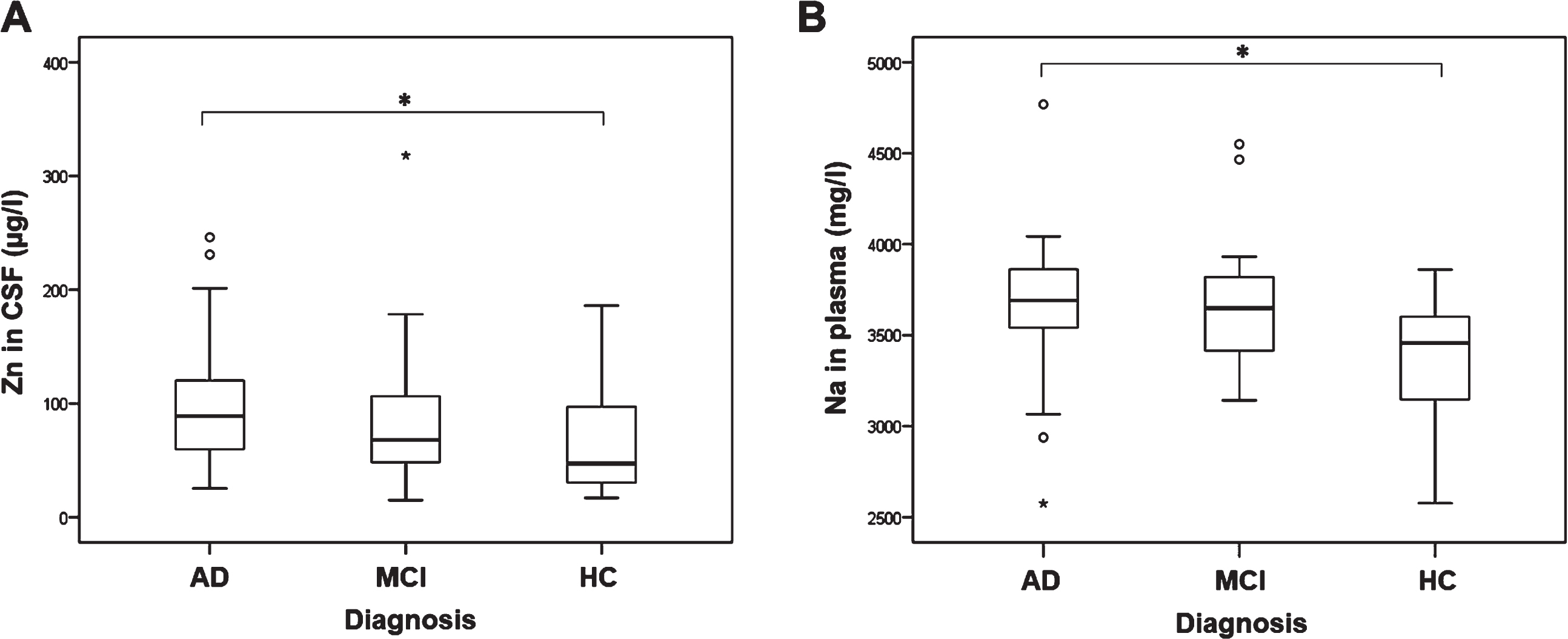
Levels of A) zinc measured in CSF (1p = 0.027*, 2p = 0.051) and B) sodium measured in plasma of AD, MCI patients and HC (1p = 0.004*, 2p = 0.002*, 3p = 0.013*, 4p = 0.097, 5p = 0.106). 1Uncorrected. 2Corrected for age and sex. 3Corrected for hypertension, age, and sex. 4Corrected for cardiovascular diseases, age, and sex. 5Corrected for hypertension, cardiovascular diseases, age, and sex. (*p ≤ 0.05).
Levels of metals in AD, MCI patients, and HC with different APOE genotypes
No significant difference in distribution of APOE genotypes and alleles was observed among AD, MCI patients, and HC (Table 3).
Count (N) and frequencies (%) of APOE gene polymorphism (rs7412 and rs429358) genotypes and alleles in AD and MCI patients and HCs
AD, Alzheimer’s disease; APOE, apolipoprotein E; HC, healthy control; MCI, mild cognitive impairment.
A significant increase in sodium plasma levels was observed in AD, MCI patients, and HC carriers of ɛ4 APOE genotype (ɛ4/ɛ4+ɛ4/ɛx) (F(1,90) = 4.354, p = 0.040; Fig. 2; Table 4). Sodium plasma levels were also significantly increased in AD, MCI patients, and HC carriers of ɛ4/ɛx APOE genotype compared to carriers of ɛx/ɛx APOE genotype (F(2,90) = 3.414, p = 0.037; p = 0.037; Fig. 2; Table 4). Additionally, sodium plasma levels were significantly increased in carriers of ɛ4/ɛ3 APOE genotype compared to carriers of ɛ3/ɛ3 APOE genotype in AD, MCI patients, and HC (H test = 12.530, df = 4, p = 0.014; p = 0.020) and AD and MCI patients (H test = 10.427, df = 4, p = 0.034; p = 0.050; Fig. 2G-H), but this association was lost after introduction of hypertension and cardiovascular diseases as covariates (Table 4).
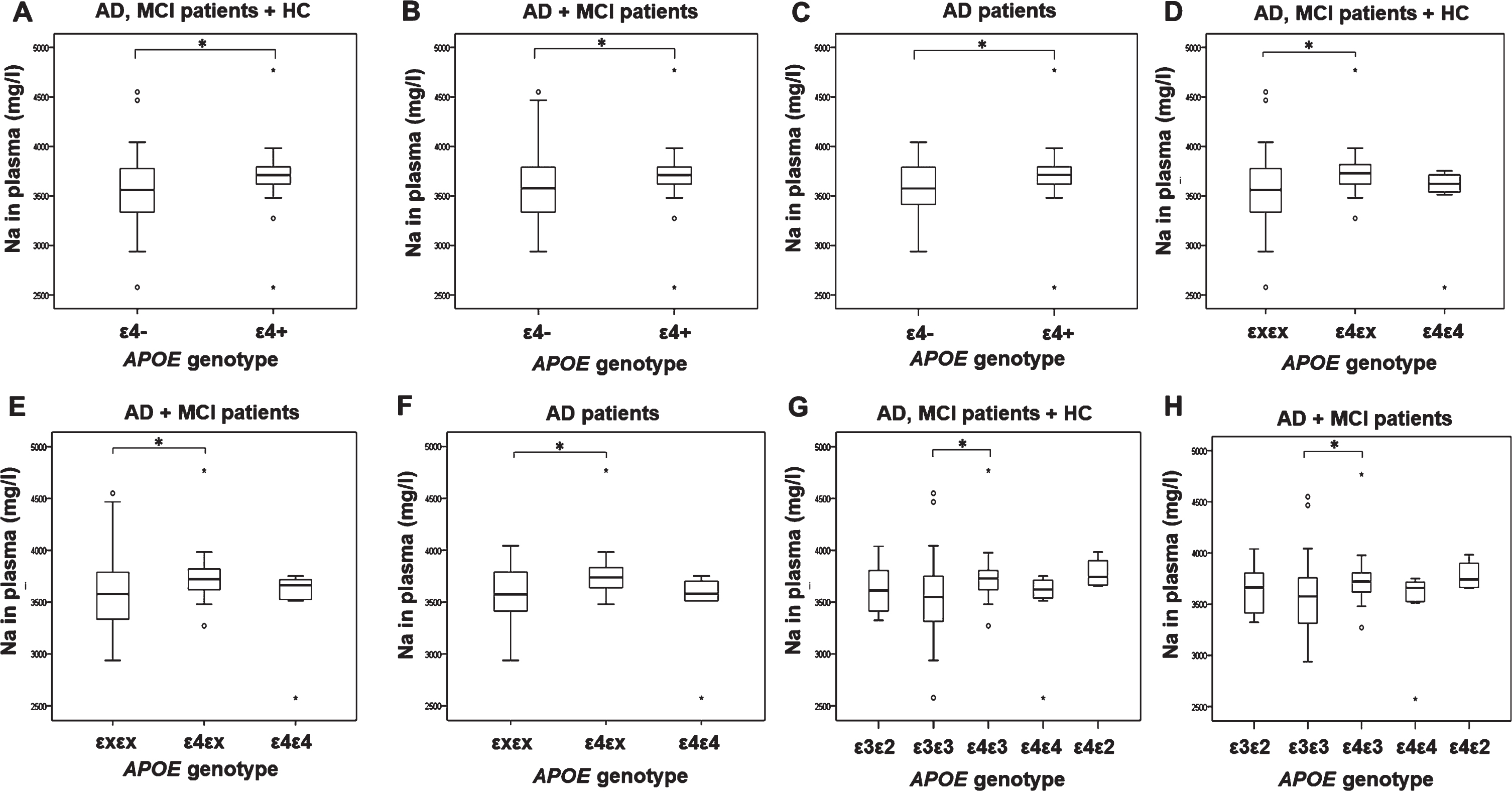
Levels of sodium measured in plasma of AD, MCI patients and HC with different APOE genotypes. A) 1p = 0.002*, 2p = 0.002*, 3p = 0.042*, 4p = 0.038*, 5p = 0.040*, B) 1p = 0.006*, 2p = 0.009*, 3p = 0.078, 4p = 0.071, 5p = 0.074, C) 1p = 0.010*, 2p = 0.050*, 3p = 0.352, 4p = 0.398, 5p = 0.398, D) 1p = 0.002*, 2p = 0.002*, 3p = 0.040*, 4p = 0.034*, 5p = 0.037*, E) 1p = 0.006*, 2p = 0.008*, 3p = 0.099, 4p = 0.089, 5p = 0.094, F) 1p = 0.014*, 2p = 0.026*, 3p = 0.171, 4p = 0.139, 5p = 0.137, G) 1p = 0.020*, 2p = 0.009*, 3p = 0.069, 4p = 0.068, 5p = 0.066, H) 1p = 0.050*, 2p = 0.038*, 3p = 0.105, 4p = 0.102, 5p = 0.097. 1Uncorrected. 2Corrected for age and sex. 3Corrected for hypertension, age, and sex. 4Corrected for cardiovascular diseases, age, and sex. 5Corrected for hypertension, cardiovascular diseases, age, and sex. (*p ≤ 0.05).
Analysis of sodium levels between patients with different APOE genotype with covariate analysis (with age, sex, hypertension and cardiovascular diseases as covariates)
AD, Alzheimer’s disease; APOE, apolipoprotein E; HC, healthy control; MCI, mild cognitive impairment. *p ≤ 0.05.
Plasma boron levels were significantly decreased in AD and MCI patients carrying ɛ4/ɛ4 APOE genotype compared to patients carrying ɛx/ɛx APOE genotype (F(2,105) = 3.998, p = 0.021; p = 0.020; Fig. 3A). Decrease in boron plasma levels was also observed in AD and MCI patients carriers of ɛ4 allele (F(1,105) =4.077, p = 0.046; Fig. 3).
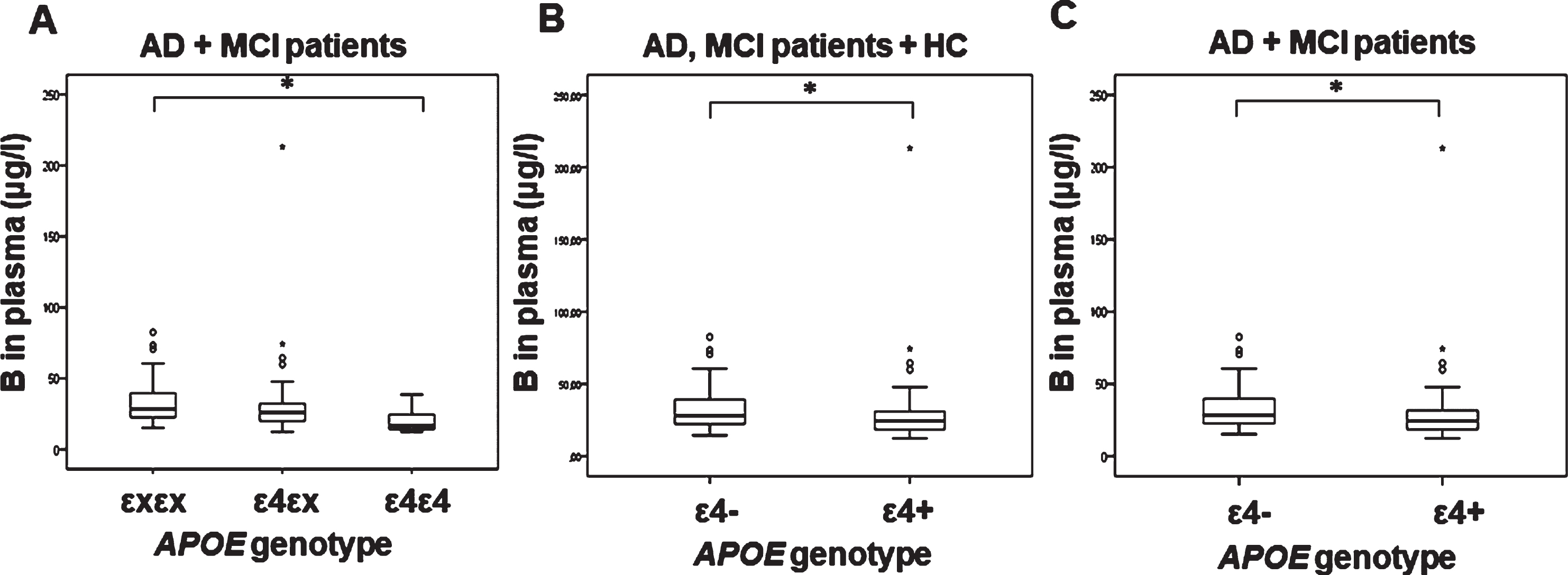
Levels of boron measured in plasma of AD, MCI patients, and HC with different APOE genotypes. A) 1p = 0.012*, 2p = 0.020*, B) 1p = 0.045*, 2p = 0.053, C) 1p = 0.043*, 2p = 0.046*. 1Uncorrected. 2Corrected for age and sex. (*p ≤ 0.05).
Copper plasma levels (U = 456, Z = –1.987, p = 0.047; Fig. 4A) and magnesium plasma levels (F(1,69) = 6.041, p = 0.016; Fig. 4B) were increased in AD patients carrying ɛ4 allele (for copper, significance was lost after introduction of covariantes age and sex [F(2,68) = 3.665, p = 0.060]). Additionally, zinc plasma levels (F(2,68) = 3.556, p = 0.034; Fig. 5A), calcium plasma levels (F(2,69) = 4.155, p = 0.020; p = 0.028; Fig. 5B), and cobalt plasma levels (F(2,116) = 3.069, p = 0.050; p = 0.040; Fig. 5C) were increased in patients carrying ɛ4/ɛx APOE genotype compared to patients carrying ɛ4/ɛ4 APOE genotype.
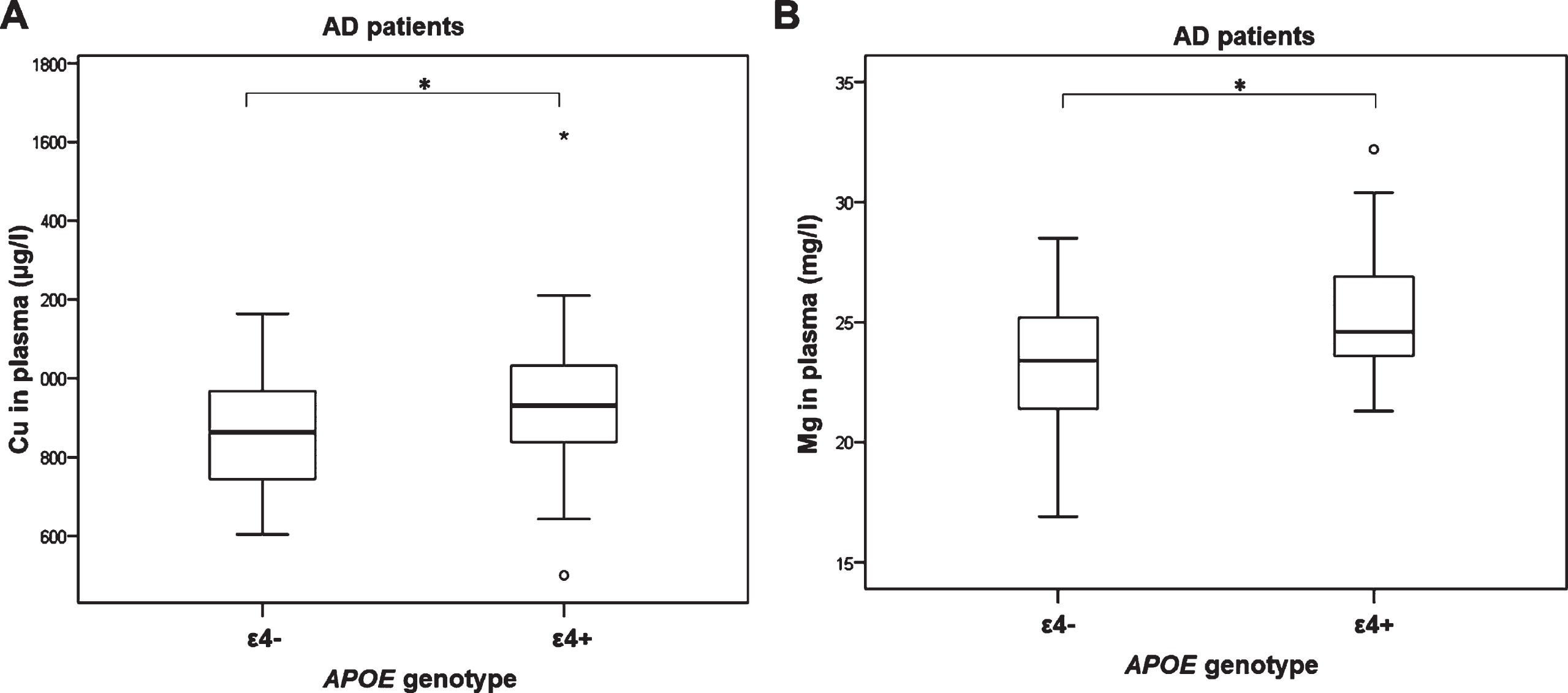
Levels of A) copper (1p = 0.047*, 2p = 0.060) and B) magnesium (1p = 0.010*, 2p = 0.016*) measured in plasma of AD patients with different APOE genotypes. 1Uncorrected. 2Corrected for age and sex. (*p ≤ 0.05).
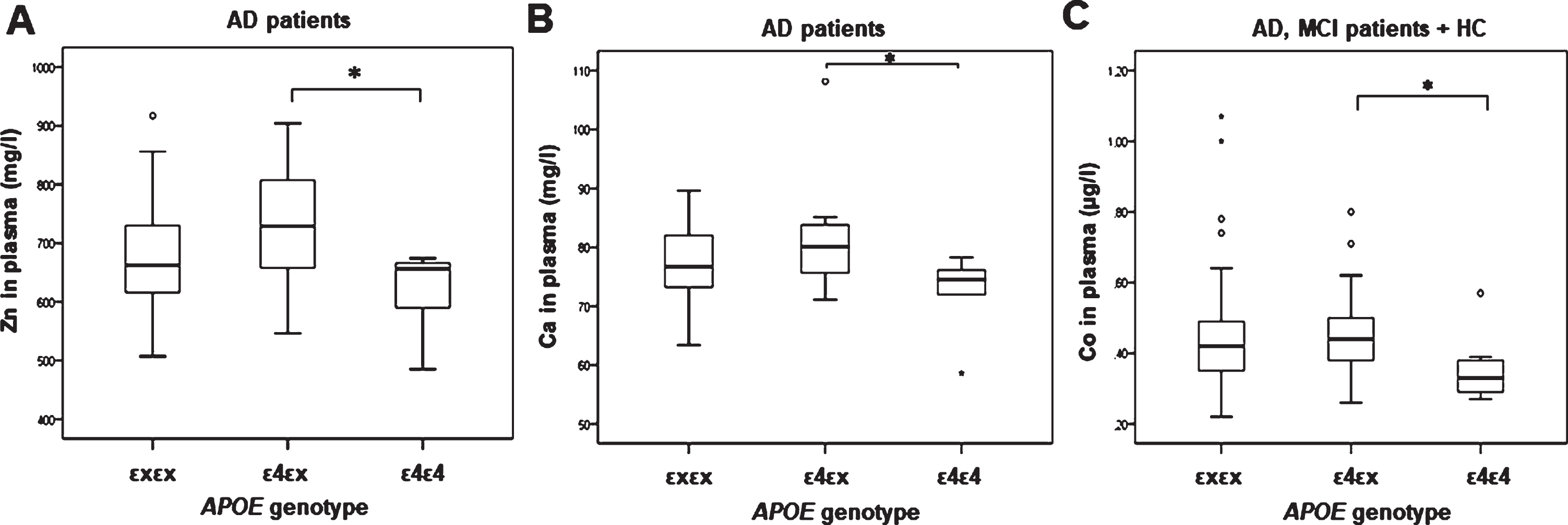
Levels of A) zinc (1p = 0.045*, 2p = 0.034*) and B) calcium (1p = 0.044*, 2p = 0.028*) measured in plasma of AD patients and C) cobalt (1p = 0.033*, 2p = 0.040*) measured in plasma of AD, MCI patients and HC with different APOE genotypes. 1Uncorrected. 2Corrected for age and sex. (*p ≤ 0.05).
Plasma levels of iron, molybdenum, and manganese, and CSF levels of copper, zinc, iron, sodium, magnesium, calcium, cobalt, molybdenum, manganese, and boron did not differ significantly among patients carrying different APOE genotype. No significant difference in the levels of CSF ferritin was observed among patients with different APOE genotype.
DISCUSSION
In this study we demonstrate increases in plasma levels of sodium, copper, and magnesium in carriers of ɛ4 allele of APOE. Additionally, sodium, zinc, calcium and cobalt plasma levels were increased in carriers of the ɛ4/ɛx genotype. Conversely, boron plasma levels were decreased in carriers of the ɛ4 allele and ɛ4/ɛ4 genotype. We also observed an increase in CSF zinc levels and plasma sodium levels in AD patients compared to HC.
Squitti and collaborators were among the first to observe an increase of serum copper in APOE ɛ4 carriers compared to APOE ɛ4 non-carriers [12, 21]. Zappasodi et al. tested the correlation of “free” serum copper and electroencephalographic (EEG) activity in AD and reported stronger correlation between serum copper and temporal α1 EEG activity in APOE ɛ4 carriers compared to APOE ɛ4 non-carriers [22]. Gonzalez et al. reported higher levels of serum zinc and copper in AD patients APOE ɛ4 carriers compared to non-carriers [13]. Miyata and Smith proposed that ApoE antioxidative activity might be mediated by sequestration of copper in an isoform-dependent manner [11]. In fact, copper can even affect transcription of the APOE gene [23]. Additionally, patients carrying the APOE ɛ4 genotype have earlier onset of symptoms of Wilson disease [24], a rare genetic disorder characterized by copper overload [25]. These results support our observation of increased copper plasma levels in AD patients carrying APOE ɛ4 allele.
Increased risk for AD in APOE ɛ4 carriers could be at least in part due to zinc dyshomeostasis in AD brains. When testing ApoE proteolysis in the presence of zinc, the ApoE4 isoform is the most sensitive to proteolysis compared to ApoE2 and ApoE3 isoforms [10]. In addition to the fact that metal-induced aggregation of Aβ (either by Zn or Cu) is highest in the presence of ApoE4 [26], Oh et al. showed that zinc promotes ApoE and Aβ aggregation into larger ApoE/Aβ complexes, making Aβ more resistant to Aβ-degrading proteases [27]. In turn, ApoE can regulate zinc homeostasis, as reduction in synaptic zinc levels and reduced expression of ZnT3 (a zinc transporter required for accumulation of zinc in synaptic vesicles) was observed in APOE knockout mice [28]. Because synaptic zinc is important for long-term potentiation [29], decrease in its levels can lead to cognitive impairment. A study of APOE-targeted gene replacement mice showed no difference in the levels of zinc, copper, and iron measured in the liver of mice with different APOE genotypes [30]. However, a study in humans showed higher levels of serum zinc in APOE ɛ4 carriers with AD compared to non-carriers [13]. The present study shows significantly elevated zinc plasma levels in ɛ4/ɛx heterozygotes compared to ɛ4/ɛ4 homozygotes and a significant increase in zinc CSF levels in AD patients compared to HC, while there was no difference in zinc plasma levels in AD versus MCI, and MCI versus HC groups.
A neuroimaging study [15] revealed that cortical iron (measured by quantitative susceptibility mapping magnetic resonance imaging [MRI]) and APOE ɛ4 allele synergistically interact with the default mode network (DMN) activity. DMN function is altered early in AD [31–33]. Kagerer et al. proposed that APOE ɛ4 allele could accelerate the accumulation of iron in the brain that could contribute to DMN dysfunction [15]. Moreover, Van Bergen et al. observed increased brain iron levels (measured by quantitative susceptibility mapping) in carriers of the APOE ɛ4 allele [34]. Ayton and collaborators also reported an increase in CSF ferritin levels in carriers of the APOE ɛ4 allele [35, 36]. As CSF ferritin levels likely reflect the levels of iron in the brain, they proposed that elevation of the brain iron could be a possible mechanism through which APOE ɛ4 allele contributes to the increased risk for AD [35]. Additionally, Tisato et al. observed that presence of certain variants in genes responsible for iron metabolism (such as HFE 282Y allele) can reduce the APOE ɛ4-associated risk for AD [37]. It has been shown that ApoE binds iron [11] and another study revealed that iron upregulates ApoE levels in cultured neurons and astrocytes [38]. However, no changes were found in brain iron levels, measured postmortem in gray matter in carriers of the APOE ɛ4 allele [39]. Also, as mentioned above, no difference in the levels of zinc, copper, and iron measured in the liver of mice with different APOE genotypes [30]. These two studies support our results as we did not observe the difference in CSF and plasma iron levels in patients with different APOE genotypes. We also did not observe a difference in CSF levels of ferritin in patients with different APOE genotypes.
In addition, the present investigation reveals a significant increase in sodium plasma levels in carriers of the APOE ɛ4 allele and in patients carrying the ɛ4/ɛx and ɛ4/ɛ3 APOE genotypes. Also, sodium plasma levels were significantly increased in AD patients compared to HC. However, an increase in sodium CSF levels was observed in hypertensive patients with history of familial AD [40]. Also, quantitative sodium imaging using ultrahigh-field MRI revealed increase in tissue sodium concentrations in many AD brain regions [41, 42]. Moreover, an increase in sodium levels in frontal and parietal cortex was detected in postmortem AD brains [43]. These authors did not observe any changes in sodium CSF levels, as in our study. They also showed that treatment of astrocytes with Aβ leads to an increase in intracellular levels of sodium, suggesting that imbalance in cell ion homeostasis in AD brain can be triggered by Aβ and thus could contribute to the pathophysiology of AD [43]. Some studies indicate that higher dietary sodium intake might be associated with impaired cognitive function, although with mixed results (for a systematic review, see [44]). When analyzing sodium levels among groups, we introduced hypertension and cardiovascular diseases as additional covariates. Sodium levels show a positive association with blood pressure [45], while hypertension is a major risk factor for development of cardiovascular diseases [46, 47]. In addition, various cardiovascular pathologies are observed in AD (cerebral amyloid angiopathy, cerebral arteriosclerosis, small blood vessel disease, microvascular degeneration and dysfunction of blood-brain barrier) [48, 49]. By introducing covariates, we wanted to exclude the possibility that differences in sodium levels between the groups with different APOE genotypes and diagnoses is the consequence of cardiovascular pathology rather than AD pathology. Although after introduction of these covariates, statistical significance was lost in some groups, but when including all cases (AD, MCI patients, and HC), statistical significance remained. Thus, we concluded that the observed alterations in sodium levels between patients with different APOE genotype and patients with different diagnoses is the consequence of AD pathology, not cardiovascular pathology.
Boron deprivation can affect cognitive performance and lead to poorer performance on short-term memory tasks [50–52]. The results of our study support these finding as we observed a significant decrease in boron plasma levels in carriers of ɛ4 allele and ɛ4/ɛ4 APOE genotypes.
Impaired calcium signaling is a hallmark of many neurodegenerative disorders, including AD [53]. There is ample evidence of ApoE interaction with calcium. A study in primary neurons collected in APOE wild-type and knockout mice showed that after a mechanical injury, rates of apoptosis and intercellular calcium levels were higher in ApoE4 neurons [14]. These authors hypothesized that APOE polymorphisms can influence calcium levels. Our results support this view. Compared to carriers of the ɛ4/ɛ4 genotype, we observed increased calcium plasma levels in patients carrying the ɛ4/ɛx APOE genotype. In embryonic rat hippocampal neurons, Tolar and collaborators showed that ApoE and truncated ApoE peptide lead to increased intracellular calcium levels and neuronal death [54]. Also, treatment of primary cerebral cortical neurons isolated from APOE knockout mice with ApoE4 lead to calcium overload through N-methyl-
Treatment of streptozotocin-induced rat models of sporadic AD with magnesium sulfate decreased tau protein phosphorylation and had positive effect on cognitive functions and synaptic plasticity [61]. Magnesium deficiency in diet was also associated with impaired memory [62], while magnesium supplementation improved memory [63–65]. Decreased levels of magnesium were detected in the brain and blood cells of AD patients [66, 67]. Zhu et al. recently showed that optimal dietary magnesium intake improves cognitive function at least in part through modification of APOE methylation [68]. However, in apparent contrast to these data, we also observed increased levels of plasma magnesium in carriers of the ɛ4 APOE allele. Interestingly, we observed a significant increase in cobalt plasma levels in patients carrying the ɛ4/ɛx APOE genotype compared to carriers of ɛ4/ɛ4 genotype. Increased levels of cobalt have been reported in brains of AD patients, especially in nucleus basalis of Meynert [69].
CONCLUSION
In conclusion, our study reveals a strong association between copper, zinc, sodium, magnesium, calcium, and cobalt, as well as the metalloid boron, with APOE genotype in AD and MCI patients. As previous studies addressed mostly the association of calcium, copper, iron and zinc with ApoE4-mediated toxicity, our findings indicate that additional in vivo and in vitro studies into the molecular basis of the association of other essential metals and metalloids with ApoE-dependent mechanisms are warranted. The most notable finding of this study is the increase of sodium plasma levels and decrease in boron plasma levels in carriers of risk alleles in APOE gene that to our knowledge had not been previously observed. Additionally, variation in the plasma levels of magnesium and cobalt in patients with different APOE genotype should be further tested on larger cohorts.
Footnotes
ACKNOWLEDGMENTS
This work was funded by The Croatian Science Foundation grant IP-2019-04-3584 (“Role of blood-brain barrier, innate immunity, and tau protein oligomerization in the pathogenesis of Alzheimer’s disease”) to GŠ and by the Scientific Centre of Excellence for Basic, Clinical and Translational Neuroscience CoRE-NEURO (“Experimental and clinical research of hypoxic-ischemic damage in perinatal and adult brain”; GA KK01.1.1.01.0007 funded by the European Union through the European Regional Development Fund), and in part by NIH grants P50 AG005138 and P30 AG066514 to PRH.