
Abstract
Background:
The medicinal herb Centella asiatica has been long been used for its neuroprotective and cognitive enhancing effects. We have previously shown that two weeks of treatment with a water extract of Centella asiatica (CAW) improves cognition and activates the endogenous antioxidant response pathway without altering amyloid-β (Aβ) plaque burden.
Objective:
Here, we assess the effect of long-term treatment of CAW in the 5xFAD mouse model of Aβ accumulation.
Methods:
Four-month-old 5xFAD mice were treated with CAW in their drinking water (2 g/L) for three months at which point they underwent cognitive testing as well as analysis of Aβ plaque levels and antioxidant and synaptic gene expression. In order to confirm the involvement of the antioxidant regulatory transcription factor NRF2 on the effects of CAW on synaptic plasticity, neurons isolated from 5xFAD mice were also treated with CAW and the targeted inhibitor ML385.
Results:
Three months of treatment with CAW improved spatial and contextual memory as well as executive function in 5xFAD mice. This improvement was accompanied by increased antioxidant gene expression and a decrease in Aβ plaque burden relative to untreated 5xFAD animals. In isolated neurons, treatment with ML385 blocked the effects of CAW on dendritic arborization and synaptic gene expression.
Conclusion:
These results suggest that prolonged CAW exposure could be beneficial in Alzheimer’s disease and that these effects likely involve NRF2 activation. Moreover, these findings suggest that targeting NRF2 itself may be a relevant therapeutic strategy for improving synaptic plasticity and cognitive function in Alzheimer’s disease.
INTRODUCTION
Alzheimer’s disease (AD) is an irreversible progressive form of dementia that affects 44 million people worldwide [1]. Currently there are no disease modifying therapies available owing in part to the incomplete understanding of the biology underlying AD. The pathological hallmarks of AD are the formation of amyloid-β (Aβ) plaques and neurofibrillary tangles which lead to synaptic degeneration, neuronal loss, and severe cognitive impairments, including profound deficits in memory and executive function.
Increased oxidative stress is also widespread in the AD brain [2] and considered to be an early event contributing to cognitive decline [3]. Oxidative stress occurs when there is an excess of free radicals wh-ich can cause oxidative damage to cellular macromolecules. Markers of increased oxidative stress are evident in AD patients [4–6] as well as in experimental models of AD where increased oxidative stress results in increased Aβ production, synaptic dysfunction, neural death, and cognitive impairment [7–11]. The endogenous antioxidant response pathway is regulated by the transcription factor NRF2 (nuclear factor erythroid 2-related factor 2, also called NFE2L2) and activation of NRF2 has been shown mitigate oxidative damage and cognitive deficits in models of AD and other neurological diseases [12–16].
Centella asiatica (L.) Urban (Apiaceae) is an herb used in Ayurvedic and Chinese traditional medicine to boost memory and enhance cognitive function [17]. This traditional usage has been supported by several human trials demonstrating cognitive enhancing effects in both healthy and impaired populations with no reported adverse events [18, 19]. This excellent safety profile supports the classification of Centella asiatica as a Class 1 herb (one that can be safely consumed when used appropriately) by the Botanical Safety Handbook [20] and has made it an attractive candidate for wider clinical use. Extracts of Centella asiatica have been shown to have neuroprotective effects in models of neurodegenerative conditions including AD [21, 22]. We and other have shown that the water extract of Centella asiatica (CAW) can protect against Aβ cytotoxity, activate NRF2 and reduce oxidative damage as well as improve mitochondrial function in both cell and animal models of AD [23–29]. CAW has been shown to possess cognitive enhancing effects. Our laboratory has previously shown that two weeks of treatment with CAW can activate NRF2, increase synaptic density, and improve memory and executive function in models of Aβ accumulation and also healthy aging [23, 31]. In Aβ overexpressing mice, the cognitive enhancing effects of 3–5 weeks of CAW treatment were not consistently associated with decreases in plaque burden [23, 31]. However, the effects of prolonged exposure to CAW on these endpoints remain unknown. This study aims to address this issue by evaluating the effects of prolonged CAW treatment in the 5xFAD mouse model of Aβ accumulation, including cognitive testing as well as evaluations of effects on the expression of NRF2 and its target genes. Since it is impractical to chronically inhibit NRF2 in vivo, these studies go on to evaluate the role of NRF2 in CAW’s neuroprotectant effects by using isolated primary neurons from these animals.
MATERIALS AND METHODS
CAW extract
CAW was prepared as previously described [24, 28]. Briefly, Centella asiatica was obtained from Oregon’s Wild Harvest (Redmond, OR) and the water extract was prepared by refluxing 1200 g of the raw plant material with 15 L of water for about 1.5 h, in several batches. This extract was filtered and lyo-philized to a powder. A representative sample of both the raw Centella asiatica and the CAW is retained by our laboratory at –20°C. CAW prepared from this plant material contained 4.4%w/w of triterpenes and 1.7%w/w mono and dicaffeoyquinic acids [32] determined by targeted liquid chromatography-high resolution tandem mass spectrometry analysis [33]. A full description of the chemical composition can be found in our recent publication [32].
Animals
5xFAD animals were generated from a founder breeding pair from The Jackson Laboratory (cat# 006554). The background strain for these mice, and the strain of the non-transgenic WT littermates, is B6SJLF1/J. Litters were kept in a climate-controlled environment with a 12 h light/dark cycle and provided with water and diet ad libitum until aged to 4 months old. All procedures were conducted in accordance with the NIH Guidelines for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee of the Portland VA Healthcare System (IACUC #4255-18). At 4 months of age (Fig. 1), 5xFAD and WT littermates were given CAW (2 g/L) in their drinking water or untreated water calculated to deliver approximately 200 mg/kg body weight/day. Water was replaced multiple times a week and treatment given for a total of four months. The amount of water consumed for each cage was recorded and estimates of the CAW water consumed for each group can be found in Supplementary Table 1. The concentration of CAW used in this study was chosen to match the concentration used in previous studies [24, 34] to facilitate a direct comparison of the results. Behavioral testing began at 7 months of age, follo-wing 3 months of CAW exposure and continued during testing. Animals were euthanized and tissue was collected after completion of behavioral testing at 8 months of age. The number of animals tested in each group was WT control female n = 6, WT CAW female n = 6, 5xFAD control female n = 4, 5xFAD CAW female n = 9, WT control male n = 5, WT CAW male n = 6, 5xFAD control male n = 5, 5xFAD CAW male n = 6.

Timeline of CAW treatment and behavioral assessment. Mice were treated with CAW (2 g/L) three months prior to the beginning of behavioral testing and treatment continued throughout the experiment. After testing, animals were sacrificed and tissue was harvested. CAW treatment lasted a total of 4 months.
Behavioral tests
Odor discrimination reversal learning (ODRL)
This test of learning and executive function occurs in three stages: shaping, acquisition, and shift. Animals were food restricted the night before testing and then given access to food in the afternoon once testing for the day was complete. All stages occurred in an opaque 8”x7”x12” chamber bisected by a removable partition.
In the shaping phase, animals were introduced to the testing chamber and trained to dig for food re-wards in bedding material that smells of lavender. Individual mice were exposed to a bowl containing the food reward (Supreme Mini Treat pellet, BioServ, Flemington, NJ) that had been filled to progressively higher levels with the bedding: 0%, 25%, 50%, 75%, and 100%. Once the animal successfully retrieved the food reward 5 times in succession, it proceeded to the subsequent training step.
The acquisition phase followed immediately after the animals completed the shaping phase. In the acq-uisition phase, mice were presented with two cups, one that contained dried beans, and one that contained string. In each trial, one material had a mint odor and the other had a vanilla odor. Each material/scent pair was alternated randomly in every trial but balanced throughout the acquisition phase such that all mice were exposed to roughly equal combinations of each odor and digging material. In the acquisition phase, the cup with the mint scent was always baited with the reward, regardless of the digging material and whether the baited cup was presented on the right or the left side of the chamber was likewise balanced throughout testing. Each trial began when a partition separating the mouse from the bowls was raised. The number of trials required for each mouse to achieve 8 correct digs of any session of 10 was recorded. Once a mouse successfully reached this criterion, it immediately moved to the shift phase.
As in the acquisition phase, in the shift phase the mouse was again presented with two cups containing the same digging materials and odors with the odor + digging material pairings and left/right location balanced throughout the trial. In the shift phase, however, the reward was always found in the cup with dried beans, regardless of odor. Again, the criteria was defined as 8 correct trials out of any session of 10 and the number of trials to reach criteria was recorded. Example pairings for each phase of the ODRL are given in Table 1.
Examples of test pairings for Odor Discrimination Reversal Learning (ODRL test). Representative combinations of odor and digging material pairings during each phase of the ODRL. D1, dried bean; D2, string; O1, vanilla; O2, mint. Bold indicates correct trial
While acquisition phase of the ODRL reflects learning and is influenced by multiple brain regions, including both the hippocampus and cortex [35], the shift phase specifically probes the cognitive flexibility domain of executive function and is very tightly controlled by the prefrontal cortex [36].
Object Location Memory (OLM) test
The OLM test of spatial memory and is mediated by the hippocampus [37]. It has three phases: habituation, training, and testing. In the habituation phase, a mouse was exposed to an empty chamber (38×38×64 cm, made of a white acrylonitrile butadiene styrene) for two 10 min sessions on two consecutive days. Next animals were moved to the training phase, where they were exposed to the same chamber with two identical objects located in the front of the chamber for 10 min once an hour over three hours. Two hours after the training phase, animals were moved to the testing phase, where they were exposed for 5 min to the same objects as the training phase, but one of the objects had been moved to a new location in the chamber. The amount of time the mouse spent interacting the with the object in the novel location as well as the object in the familiar location is recorded via the ANYmaze video tracking system (Stoelting Co, Wood Dale, IL).
Conditioned Fear Response (CFR) test
The CFR test evaluates contextual memory and has been shown to be affected by inputs from the hippocampus, cortex, and amygdala [38]. It has three phases: habituation, conditioning, and testing. In the habituation phase, an animal was exposed to a 16×16×12 inch chamber with a wire floor for 5 min. The conditioning phase occurred immediately following habituation, where the animal was exposed to three 1 s shocks (0.7A) randomly distributed over a 3 min period with no more than one shock per minute. The test phase occurred 24 h after the conditioning phase, where the animal was reintroduced once more to the same chamber but this time not exposed to any shocks. The amount of time spent freezing over a 5 min period is recorded. Freezing time is represented as the change in freezing time from the habituation phase to the test phase in order to account for any baseline differences in overall activity.
Immunohistochemistry
After euthanasia, the brain of each animal was carefully dissected, and each hemisphere was separated. One hemisphere was fixed in 4%paraformaldehyde. This hemisphere was incubated sequentially in a sucrose gradient and frozen. Forty-micron coronal sections were taken on a freezing microtome and incubated in blocking buffer (100 mM TBS, pH 8.0, 2 mg/ml bovine serum albumin, 2%horse serum, 0.5%triton X-100) for 2 h. These sections were then incubated overnight in primary antibody directed against Aβ (44–136, Invitrogen, Carlsbad, CA), then exposed to biotinylated secondary antibody (1:200, Vector Labs, Burlingame, CA) for 2 h, then to an avidin-linked peroxidase complex (ABC, Vector Labs) for 2 h and finally developed with diaminobenzidine (DAB, Sigma) in PBS. Sections were then washed, mounted in Permount (Fisher Scientific, Pittsburg, PA) on slides, coverslipped and scanned using a PrimeHisto XE as per the manual with settings of 10,000 dpi, 8-bit black and white images with an exposure of 1 minute (Pacific Image Electronics, Torrance, CA). The protein expression was quantified in at least three coronal sections per mouse: anterior, middle, and posterior hippocampus and cortex. Hippocampal and cortical areas were traced using a computerized stage and stereo investigator software (Image J, Wayne Rasband, NIH, USA). Aβ levels were expressed as percentage of hippocampus or cortex occupied by these plaques.
Gene expression
The hemisphere that was not fixed and used for immunohistochemistry was sub-dissected and fro-zen at –20°C. Portions of the frontal cortex were homogenized, and RNA was extracted using Tri-Rea-gent (Molecular Research Center) using the proto-col provided by the manufacturer. RNA was reverse transcribed with the Superscript III First Strand Synthesis kit (Invitrogen) to generate cDNA as per the manufacturer’s instructions. Relative gene exp-ression was determined using TaqMan Gene Expres-sion Master Mix (Invitrogen) and commercially available TaqMan primers (Invitrogen) for synaptop-hysin (Mm00436850_m1), post-synaptic density protein 95 (PSD95; Mm00492193_m1), NRF2 (Mm00477784_m1), NAD(P) H dehydrogenase-qui-none oxidoreductase 1 (NQO1; Mm01253561_m1), glutamatecysteine ligase catalytic subunit (GCLC; Mm00802655_m1), heme oxygenase 1 (HMOX1; Mm00516005_m1), and glyceraldehyde-3phosphate dehydrogenase (GAPDH; Hs02758991_g1). Quantitative PCR (qPCR) was performed on a StepOne Plus Machine (Applied Biosystems) and analyzed using the delta-delta Ct method.
Primary neurons
Embryos from a 5xFAD X WT cross were harve-sted to generate primary neurons. Hippocampal and cortical neurons were isolated as described in Kaech and Banker [39]. Briefly embryos were harvested at embryonic day 18 and genotyped via PCR from tail samples taken during dissection. Hippocampi were sub-dissected from embryos, gently minced and trypsinized to generate suspensions of dispersed neurons. These neurons were plated on poly-l-lysine coated plates.
Neuronal gene expression
Hippocampal neurons were plated on poly-l-lysine coated 12-well Tissue Culture plates at a density of 250,000 cells per well. Cells were cultured at 37°C and 5%CO2 for 7 days and then treated with DMSO, 50μg/ml CAW, 10μM ML385, or CAW + ML385 and allowed to continue to grow for 2 days, then media was removed and RNA was extracted, cDNA generated and relative gene expression determined as described above for the mouse tissue.
Reactive oxygen species
Hippocampal neurons were plated on poly-L-lysine coated 96-well tissue culture plates at a density of 20,000 cells per well. Cells were cultured at 37°C and 5%CO2 for 7 days and then treated with DMSO, 50μg/ml CAW, 10μM ML385, or CAW + ML385 and allowed to continue to grow for 2 days. Then media was removed, and reactive oxygen species (ROS) levels were determined by a DCFDA assay kit (enQuire Bioreagents). On day 7, in vitro cells were incubated with the fluorogenic probe for 45 min at 37°C prior to measurement. Values were normalized to protein content determined by a bicinchoninic acid (BCA) protein assay as per the manufacturer’s instructions (Pierce Biotechnology).
Dendritic arborization
Hippocampal neurons were plated on poly-l-lysine coated glass coverslips at a density of 130,000 cells per 60 mm dish containing 4 coverslips in MEM med-ium (Life Technologies), 5%FBS (Atlanta Biologicals), and 0.6%glucose (Sigma-Aldrich). After 4 h, coverslips were flipped cell side down into 60 mm dishes containing neural stem cell-derived glial cells (provided by Dr. Gary Banker, Jungers Center, OHSU) and maintained in Neurobasal Medium supplemented with 1x GlutaMAX (Life Technologies) and 1x B-27Plus (Life Technologies). Dishes were fed every week by removing 1 ml of the culture medium and adding 1 ml fresh Neurobasal media containing GlutaMAX and B-27 Plus, with the first feed (at 5 days in vitro) containing 6μM cytosine β-D-arabinofuranoside hydrochloride (AraC; Sigma-Aldrich). At 12 days in vitro, the cell culture feeding contained either CAW (50μg/ml final), ML385 (10μM final), CAW + ML385, or DMSO. At 19 days in vitro, coverslips were fixed in 4%paraformaldehyde, rinsed in PBST and stained with Anti-MAP2B (Sigma-Aldrich #M4403; 3.3μg/ml) and Goat anti-mouse IgG1-Cy3 (Jackson ImmunoResearch #115-165-205; 1.5μg/ml). Stained neurons were imaged using a Zeiss ApoTome2 microscope, blinded and analyzed for morphology via the Sholl method by using Fiji software [40]. We measured 30 non-overlapping, easily isolatable neurons per coverslip.
Statistics
All bar graphs have error bars indicating standard error of the mean. Statistical significance was determined by the General Linear Model (GLM) for all in vivo experiments except OLM and Aβ plaque area quantification. GLM was utilized to account for unequal group sizes. Post-hoc pairwise comparisons were tested using Tukey and the lsmeans statement in SAS. In each of these figures the F values and model statistical significance is presented on the graph. For the OLM analysis, preference for the novel location was determined within each group by t-test. The quantification of Aβ plaque area was likewise determined by t-test for each sex. Statistical significance for the in vitro assays was determined by ANOVA followed by Bonferroni post-hoc pairwise comparisons. Significance was defined as p≤0.05. Analyses were performed using Excel, GraphPad Prism 6, and SAS 9.4.
RESULTS
Long-term CAW treatment attenuates memory deficits in 5xFAD mice
To assess the effects of prolonged CAW treatment we evaluated memory in 5xFAD mice and their WT littermates following 3 months of CAW exposure in their drinking water using CFR and OLM tests. In the CFR test of contextual memory, the animal is allowed to freely explore a chamber after which it is exposed to a mild foot shock. The following day the animal is reintroduced to that same chamber and the amount of time that the animal spends frozen is recorded. If the mouse remembers the association between the painful stimuli and the chamber, the amount of time frozen will be higher. Performance in the CFR has been shown to be mediated by inputs from the cortex, hippocampus, and amygdala [38]. A deficit in freezing time was observed in male and female 5xFAD mice compared to CAW-treated WT mice (Fig. 2A, B). A similar trend was observed when compared to control-treated WT mice, although it did not reach statistical significance. Male and female 5xFAD mice exposed to prolonged CAW treatment exhibited freezing behavior that was indistinguishable from either control or CAW treated WT mice (Fig. 2A, B).
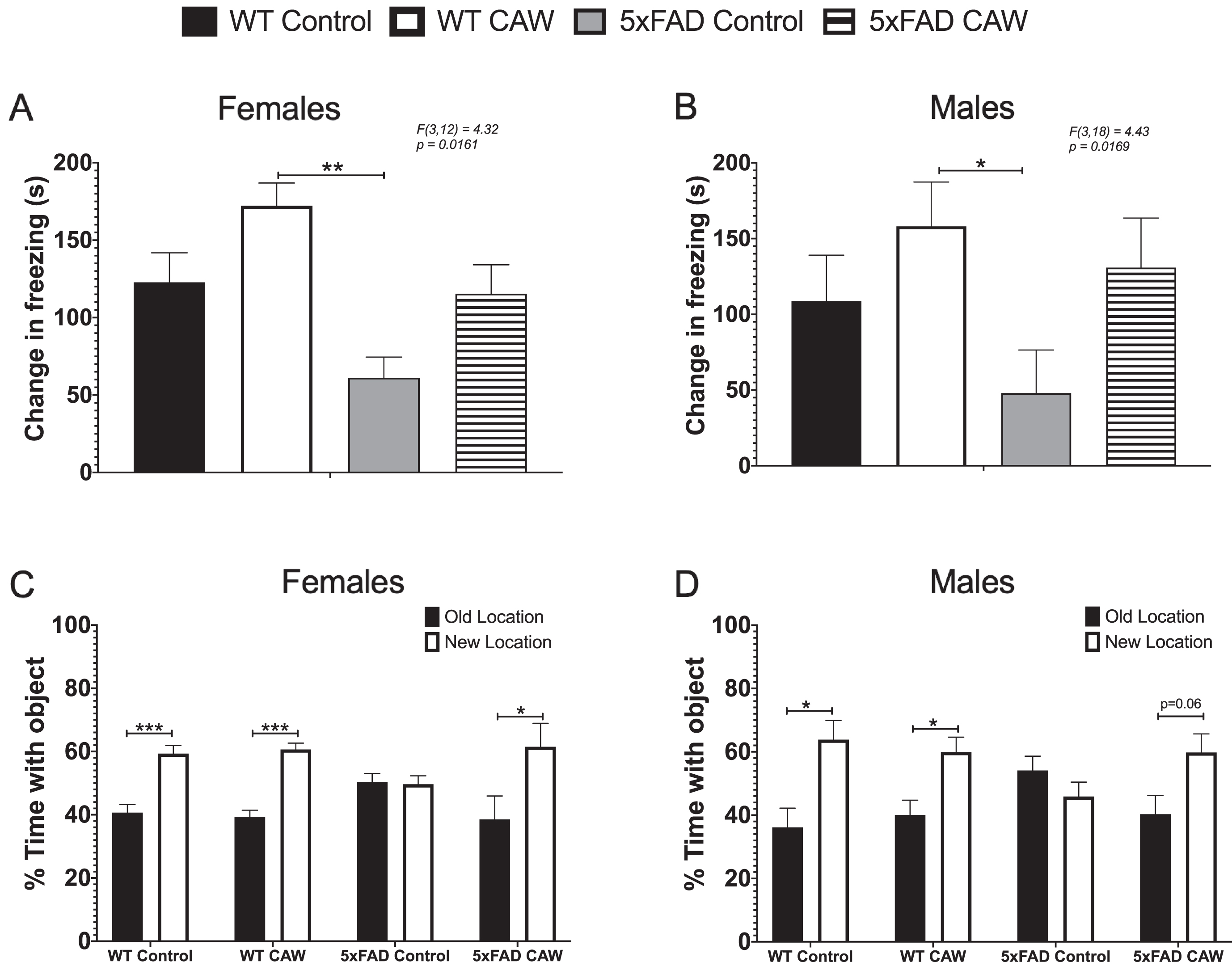
CAW improves memory in 5xFAD mice. Deficits in CFR performance, represented by the change in time freezing in the test phase relative to the habituation phase, were observed in control-treated female (A) and male (B) 5xFAD mice relative to CAW-treated WT mice (2g/L in the drinking water). A similar trend was seen as compared to WT controls, but it did not reach statistical significance in either sex. CAW attenuated these deficits in male and female mice such that the mice were not significantly different from either WT group. CAW treatment also restored the novel location preference in female 5xFAD mice (C). A similar but non-significant trend was also observed in male 5xFAD mice (D). n = 4–9, *p < 0.05, **p < 0.01, ***p < 0.001
The OLM test was used to evaluate spatial memory and has been shown to be mediated by the hippocampus [37]. In this test, the mouse is exposed to two identical objects in fixed locations throughout training. Two hours after the final training session, one object is moved to a novel location and the amount of time the mouse spends exploring each object is measured. Because of the exploratory nature of rodents, if the animal remembers the location of the object during training it will spend more time exploring the new location. We found that unlike WT animals, 5xFAD mice did not display any preference for the novel location, but CAW treatment restored a significant preference in female 5xFAD mice (Fig. 2C) and a nearly significant (p = 0.06) preference in male 5xFAD male (Fig. 2D). CAW treatment did not impact WT performance in this test in either female (Fig. 2C) or male (Fig. 2D) mice.
CAW treatment improves learning and executive function in 5xFAD mice
The ODRL test was utilized to study learning and executive function. Learning is assessed in the acquisition phase of this test while the shift phase evaluates cognitive flexibility domain of executive function. While the learning phase involved inputs from both the hippocampus and cortex, the cognitive flexibility part of the ODRL has been shown to be mediated by the prefrontal cortex [35, 36]. We found that CAW-treatment significantly reduced the number of trials necessary to reach criteria in the acquisition phase for female 5xFAD mice (Fig. 3A). A similar, but non-significant, pattern of response to CAW in the shift phase (Fig. 3B). CAW treatment had no effect on the performance of WT female mice in either phase of this test (Fig. 3A, B). There was also a reduction the number of trials required to reach criteria in both the acquisition and shift phase of this test in CAW-treated male 5xFAD mice (Fig. 2C, D). Again, as with the females, CAW treatment did not affect the performance of WT male mice.
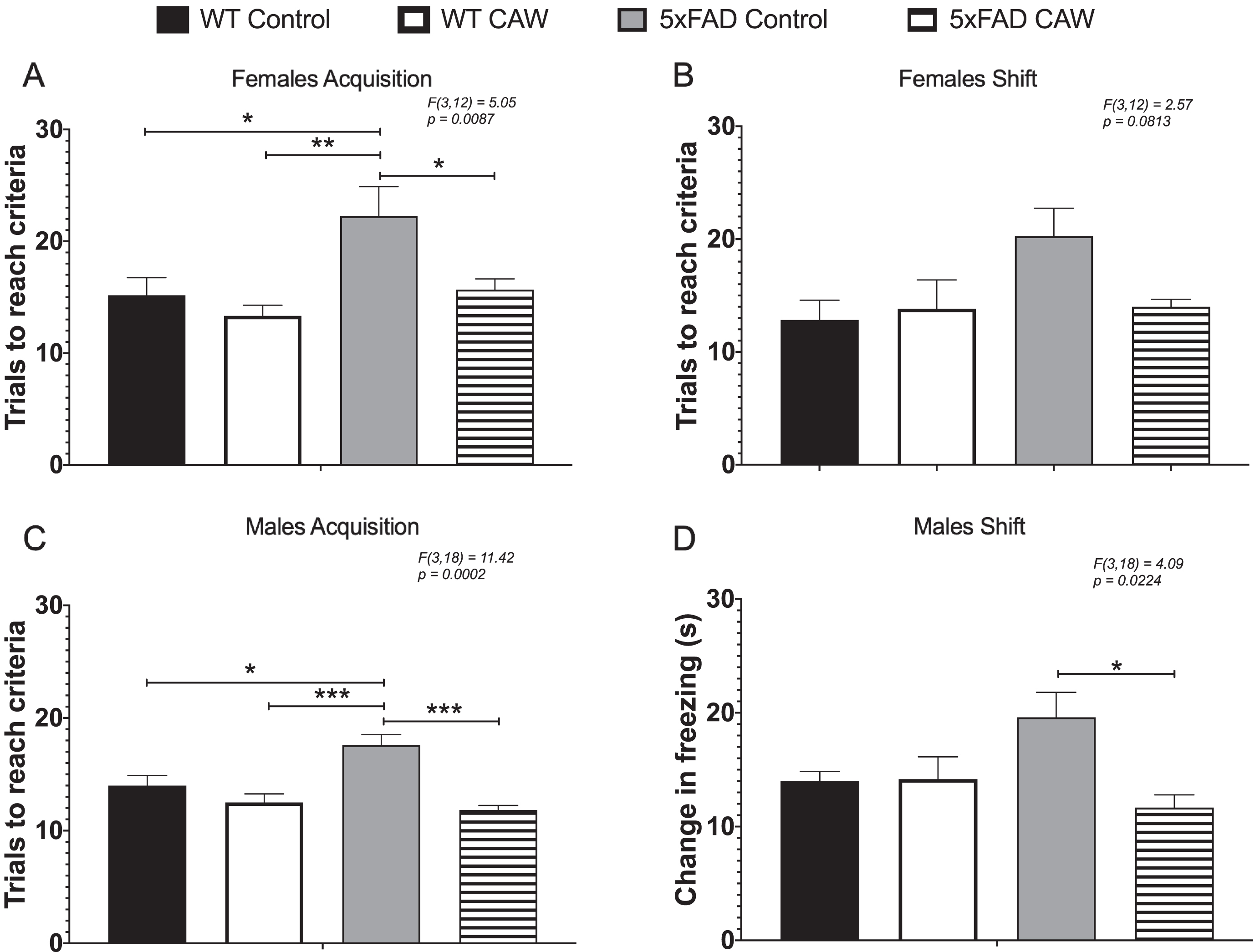
CAW improves learning and cognitive flexibility in 5xFAD but not WT mice. CAW treatment (2 g/L in the drinking water) improved impairments in ODRL performance in the acquisition phase of the ODRL in female (A) 5xFAD mice. A similar improvement was also seen in the shift phase for female CAW-treated 5xFAD mice (B) although this did not reach statistical significance. In male 5xFAD mice CAW also improved performance in both the acquisition (C) and shift (D) phases. n = 4–9, *p < 0.05, **p < 0.01, ***p < 0.001.
Long-term CAW treatment reduces Aβ plaque burden in 5xFAD mice
Aβ plaque area was significantly reduced in the cortex of both male and female 5xFAD animals treated with CAW (Fig. 4A). CAW treatment likewise reduced the hippocampal plaque burden in 5xFAD female mice but, notably, had no effect on hippocampal plaque area in male mice (Fig. 4B).
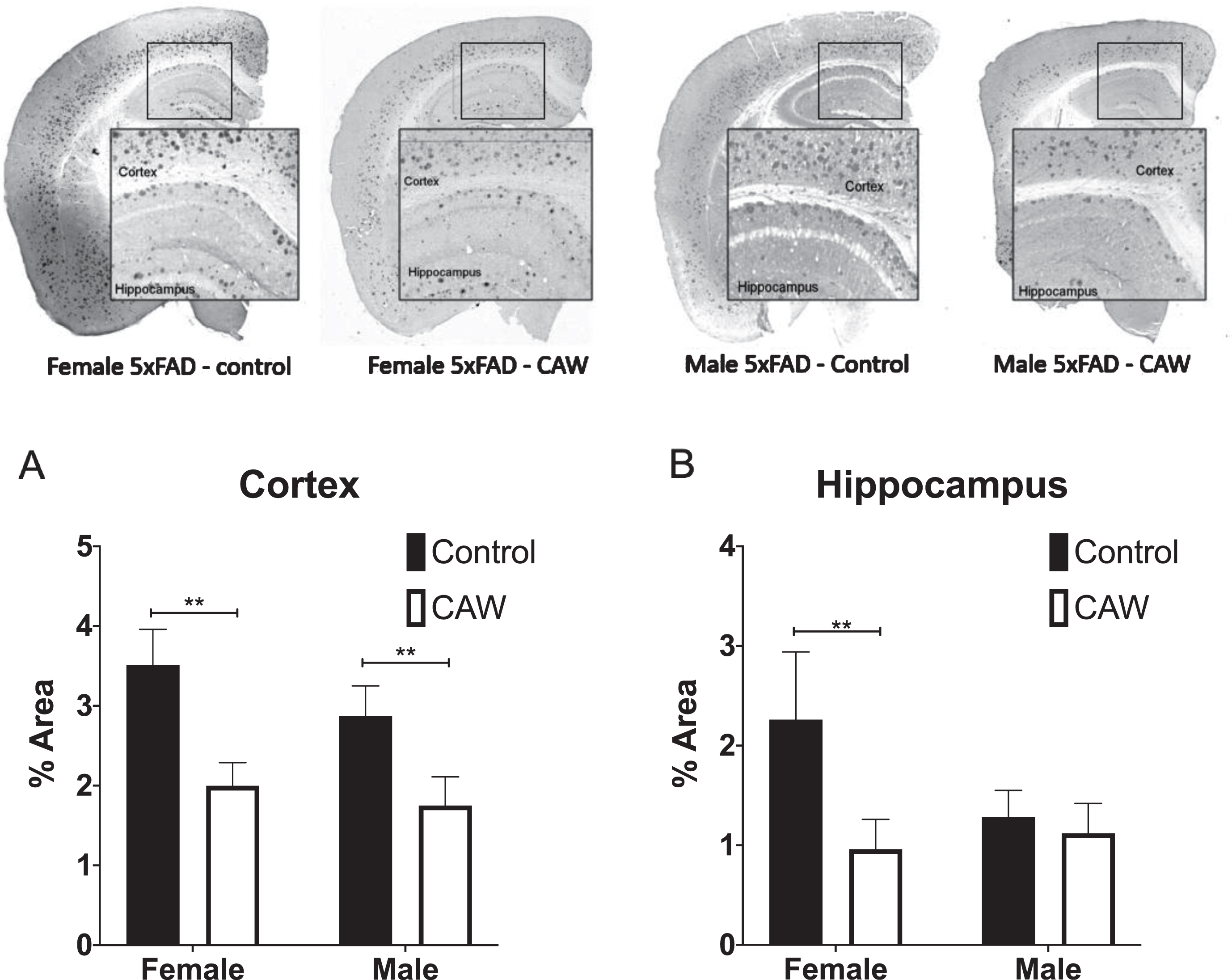
CAW reduced cortical Aβ plaque burden in female and male 5xFAD mice and hippocampal plaque burden in female 5xFAD. Representative images are shown. CAW treatment (2 g/L in the drinking water) significantly reduced Aβ plaque area in both male and female mice (A) but only reduced hippocampal plaque area in female mice (B). n = 4–9, **p < 0.01.
CAW modulates cortical gene expression in 5XFAD mice
A trend toward decreased gene expression in synaptophysin and PSD95 was observed in the frontal cortex of female 5xFAD mice relative to WT animals (Fig. 5A, B) and a trend toward restored expression with CAW treatment; however, the only group difference to reach statistical significance was between the synaptophysin expression of the CAW-treated WT mice and the control treated 5xFAD mice (Fig. 5A). In male mice, the pattern of expression of these synaptic genes was similar although none of the group differences reached statistical significance (Fig. 5C, D).
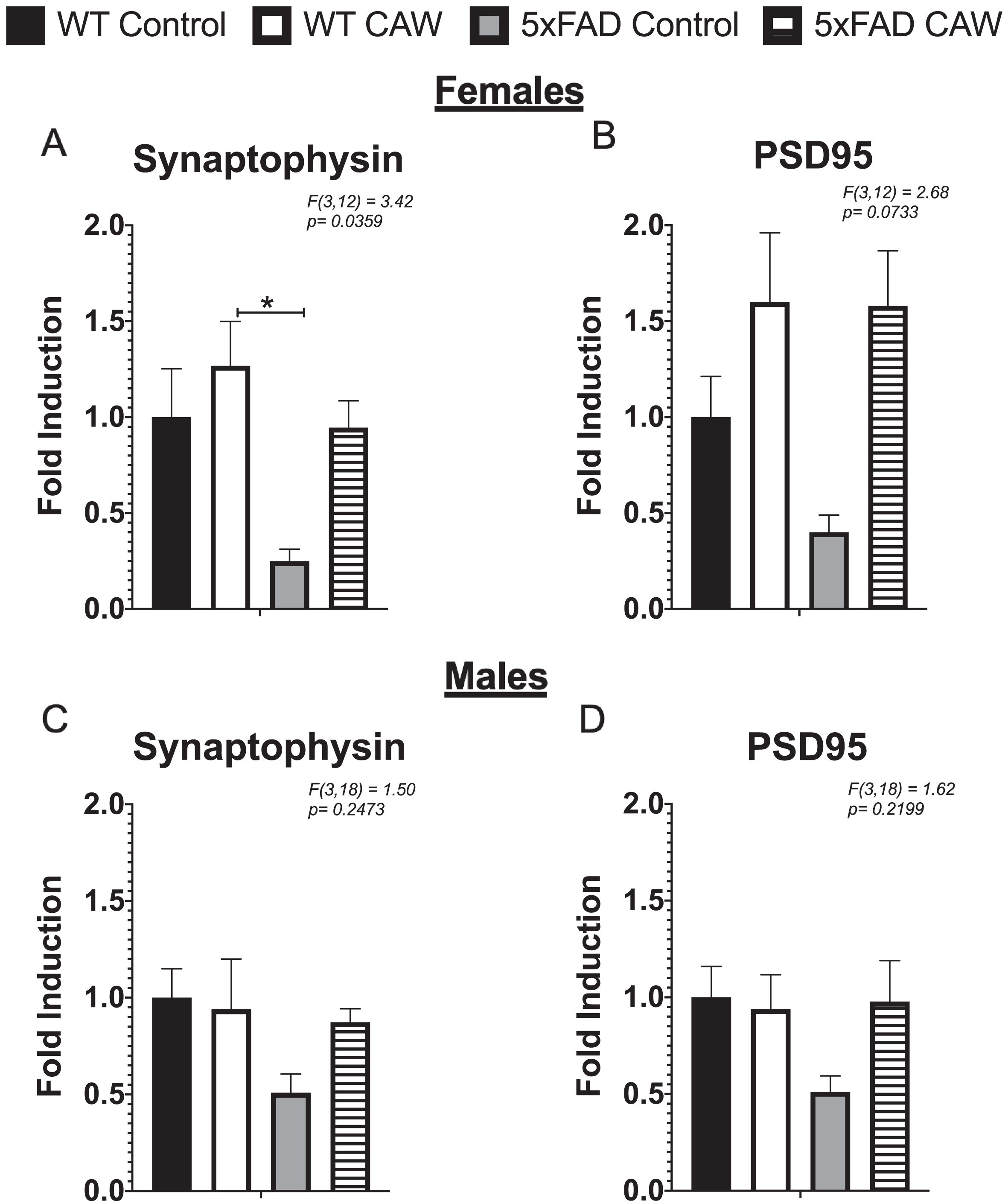
CAW does not significantly increase synaptic gene expression in the cortex of 5xFAD mice. CAW treatment (2 g/L in the drinking water) did not robustly alter the expression of synaptophysin (A) or PS95 (B) in the cortex female mice. There was similarly no significant effect of CAW treatment in male mice for either synaptophysin (C) or PSD95 (D). n = 4–9, *p < 0.05, **p < 0.01, ***p < 0.001.
CAW treatment did robustly increase the expression of NRF2 and its antioxidant target genes HMOX1 and NQO1 in the cortex of female 5xFAD mice relative to WT control mice (Fig. 6A, C, D). A similar pattern was also observed with the antioxidant target gene GCLC, but it did not reach statistical significance (Fig. 6B). For male 5xFAD mice, long-term CAW treatment increased the expression of NRF2 and all three of the target genes relative to WT control mice (Fig. 6E-H).

CAW increases antioxidant gene expression in the cortex of 5xFAD mice. There was a trend towards increased expression of NRF2 regulated antioxidant genes following CAW treatment (2 g/L in the drinking water) in the cortex in female 5xFAD amice relative to WT. This increase reached statistical significance NRF2 (A), HMOX1 (B), and NQO1 (D). The same pattern was evident in male mice with significant induction in 5xFAD mice observable for all antioxidant genes (E, F, G, H). n = 4–9, *p < 0.05, **p < 0.01, ***p < 0.001.
Antioxidant effects of CAW are blocked by NRF2 inhibition in primary neurons
CAW treatment increased the expression of NRF2 and its antioxidant response element (ARE) containing target genes in cortical neurons isolated from both 5xFAD and WT mice (Fig. 7A-D). ARE-like elements are also present in the NRF2 promoter suggesting possible autoregulation [41], an idea supported by associations between changes in NRF2 transcript and both cytoplasmic and nuclear NRF2 protein levels [41, 42]. The induction observed by CAW treatment was blocked by co-treatment with ML385, a NRF2 inhibitor [43]. Similarly, the ability of CAW to attenuate increased intracellular ROS seen in 5xFAD neurons was likewise prevented by co-treatment with ML385 (Fig. 8). Treatment with ML385 alone exacerbated this increase in ROS in 5xFAD neurons.
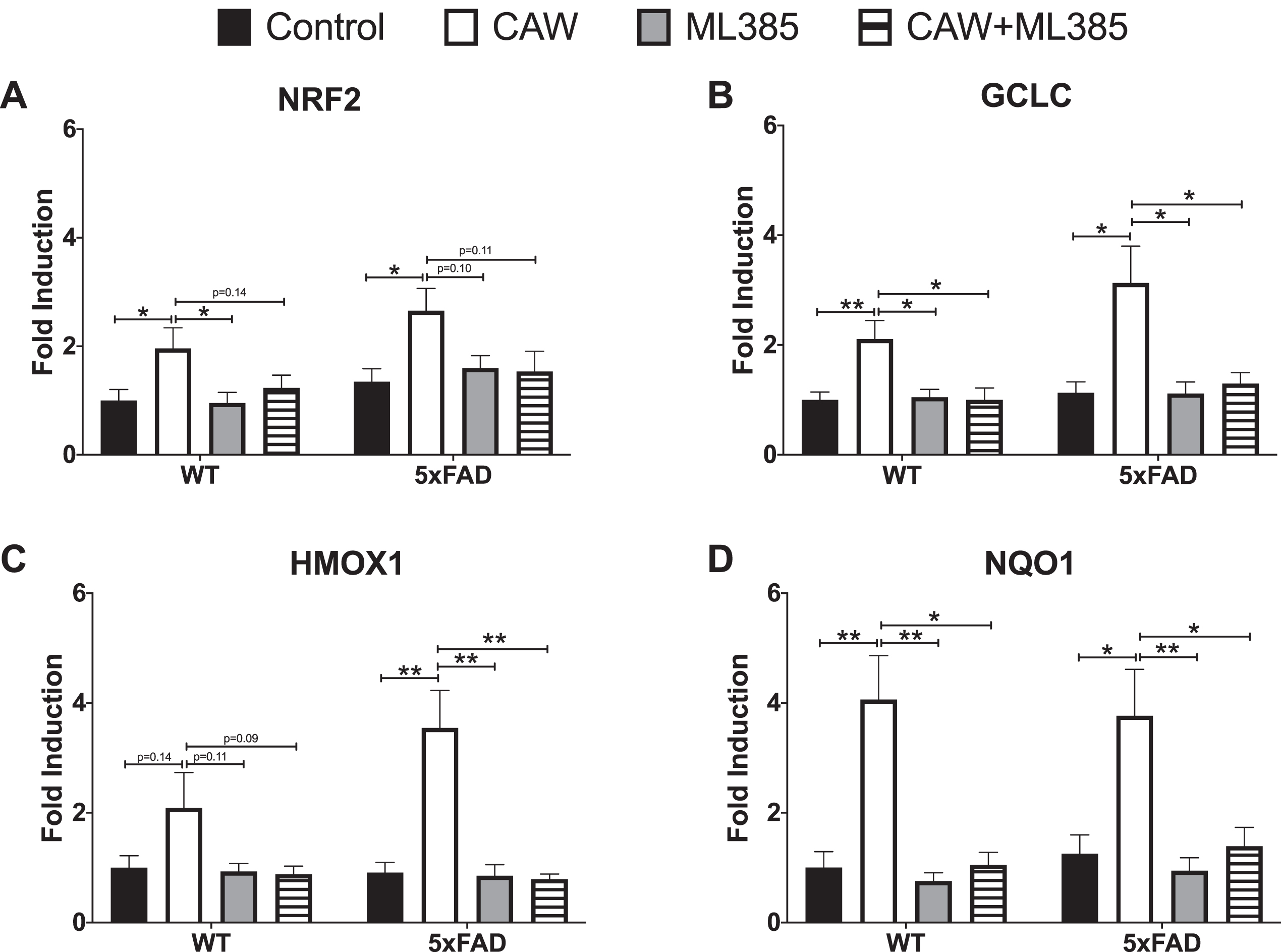
CAW (50μg/mL) increases gene expression of NRF2 and its antioxidant targets in cortical neurons isolated from 5xFAD and WT animals. Co-treatment with ML385 (10μM) blocks this effect. n = 8–12, *p < 0.05, **p < 0.01.
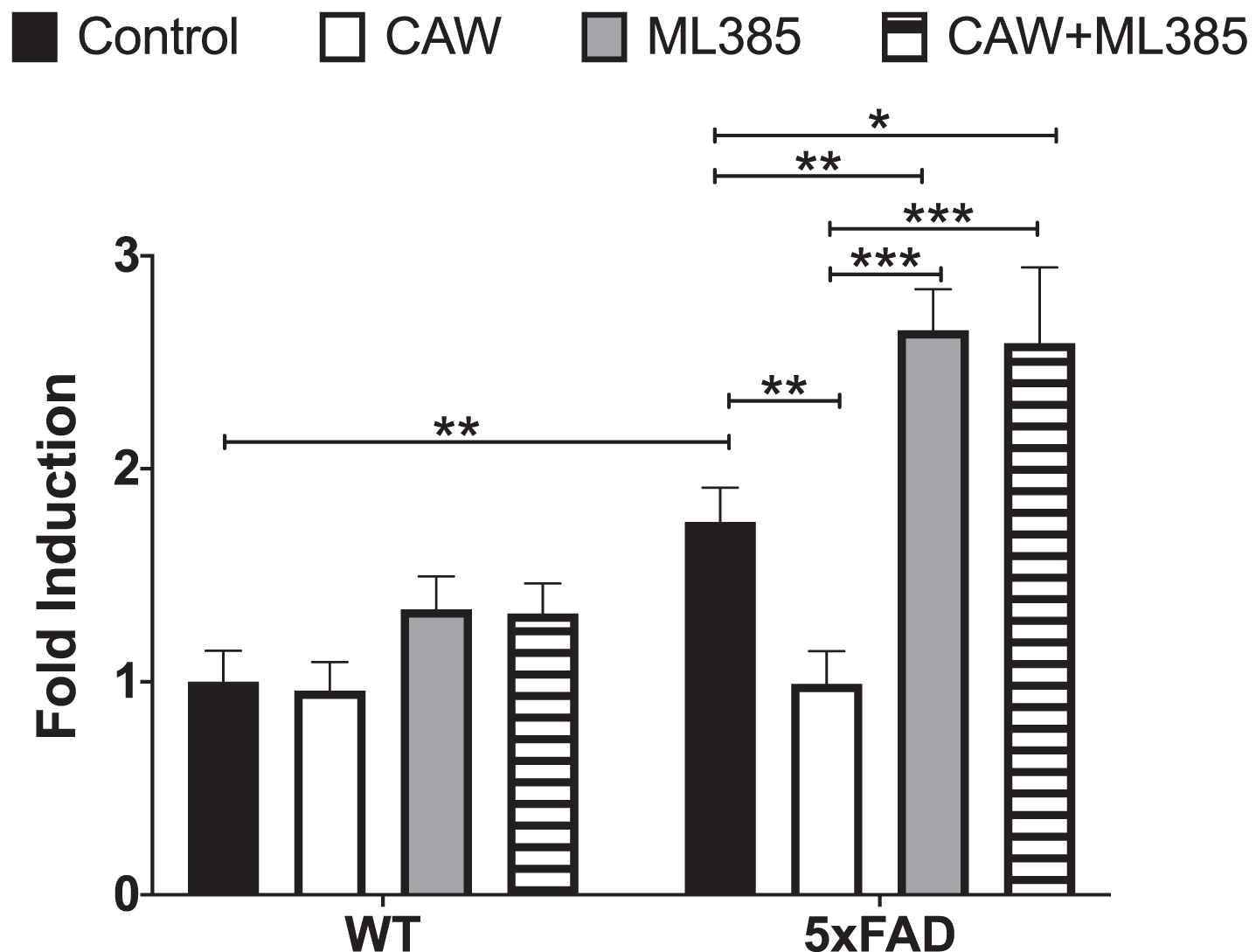
CAW attenuates increased ROS in 5xFAD neurons. Treatment with ML285 (10μM) alone and co-treatment with CAW (50μg/mL) results in further elevation of intracellular ROS. n = 8–12, *p < 0.05, **p < 0.01, ***p < 0.001.
NRF2 inhibition blocks effects of CAW on synaptic plasticity
We and others have previously reported that Aβ overexpressing neurons grown in culture for several weeks display a dystrophic phenotype characterized by reduced dendritic arborization [26, 45]. Consistent with these previous findings we found that the degree of arborization in hippocampal 5xFAD neurons was markedly reduced relative to WT neurons after 19 days in vitro (Fig. 9A-C). This reduction was ameliorated by CAW treatment. In contrast, treatment with the NRF2 inhibitor ML385 alone exacerbated this deficit in arborization and co-treatment with CAW prevented the arborization-inducing effects of the extract.
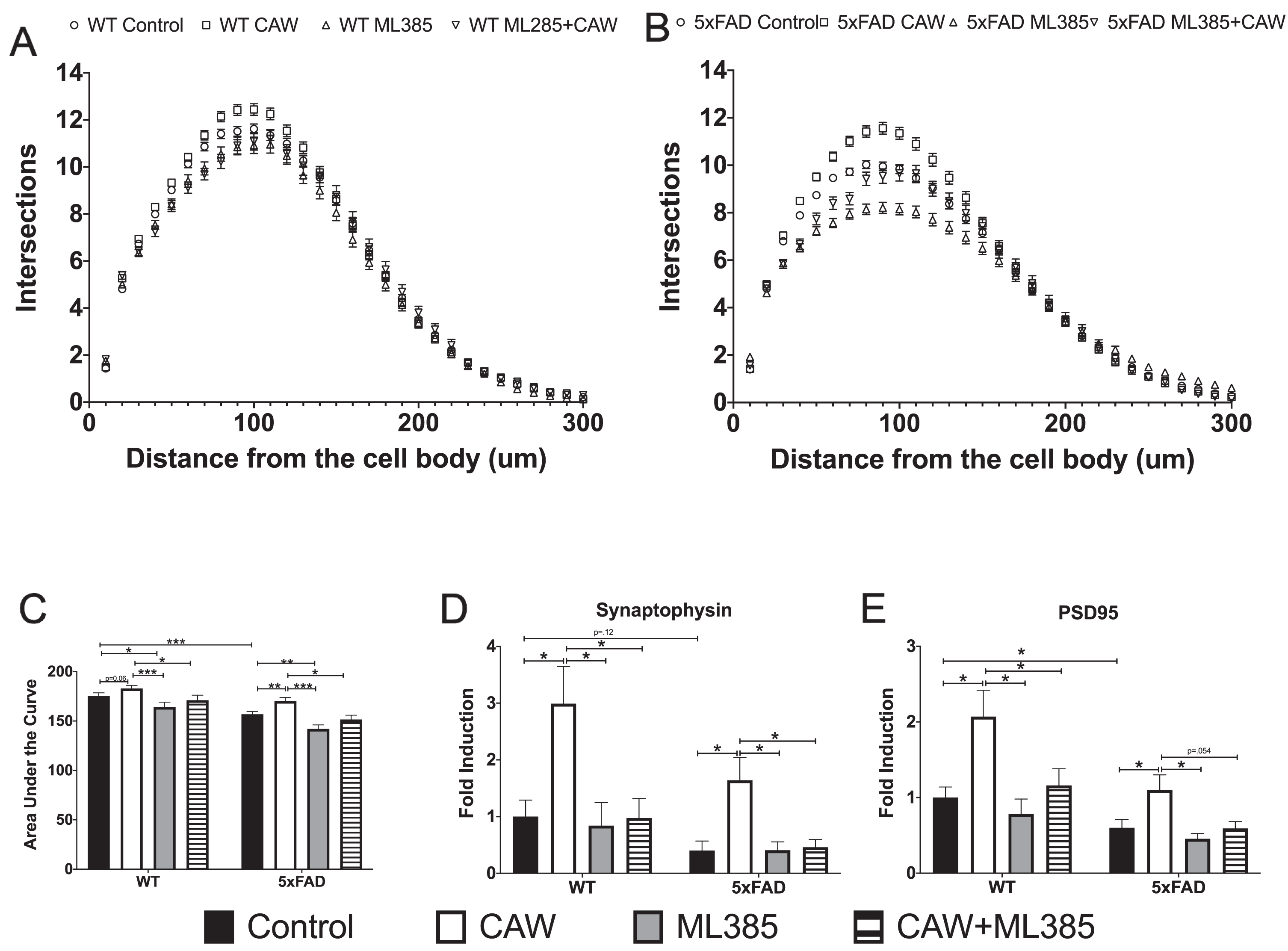
NRF2 inhibition prevents effects of CAW on dendritic arborization and synaptic gene expression. Co-treatment with ML385 (10μM) blocks CAW-induced (50μg/mL) increases dendritic arborization in WT (A) and 5xFAD (B) hippocampal neurons (data is quantified by sholl analysis curves (A and B) as well as area under the curve (C), n = 200–300 cells per condition) as well as CAW-induced increases in the expression of synaptophysin (C) and PSD95 (D) in 5xFAD and WT cortical neurons. n = 8–12, *p < 0.05, **p < 0.01, ***p < 0.001.
Because we have previously observed a robust effect of CAW on the expression of the synaptic genes synaptophysin and PSD95 [26, 46], we therefore decided to investigate it as an endpoint in ML385 and CAW co-treated neurons despite not observing an effect with prolonged treatment in vivo. Expression of both synaptophysin (Fig. 9D) and PSD95 (Fig. 9E) was significantly reduced in 5xFAD neurons relative to WT neurons and CAW increased this expression in neurons of both genotypes. This effect was lost in neurons that were co-treated with both CAW and ML385. Interestingly ML385 treatment alone did not alter synaptic gene expression in either genotype.
DISCUSSION
The medicinal herb Centella asiatica has been shown to have a cognitive-enhancing effect in aging and neurodegenerative disease in both animal models as well as human populations [23, 47–51]. Previously our laboratory has shown that two to three weeks of treatment with the water extract of the plant, CAW, significantly improved spatial memory, recognition memory, and executive function in the 5xFAD mouse model of Aβ accumulation, and these improvements were accompanied by increased NRF2-regulated antioxidant signaling [23, 24]. In this study, we investigate the cognitive and antioxidant effects of prolonged CAW treatment in 5xFAD mice and to begin to elucidate the role of NRF2 in mediating those effects using primary neurons from these animals.
We found that three months of CAW treatment had significant cognitive effects in 5xFAD mice. Impro-vements were observed in learning, memory, and executive function tasks. The magnitude of these changes was quite similar to what we have previously reported in 5xFAD mice following a two-week CAW treatment [24], suggesting that a tolerance to CAW does not develop over time and the cognitive effects are not diminished with prolonged exposure. Prolonged CAW treatment did not affect cognitive performance in WT animals. This again is similar to what we have reported in the past with shorter durations of this concentration of CAW [23, 24]. It is possible that prolonged treatment with higher concentrations of CAW would elicit a cognitive improvement in WT mice. This is in fact what we have seen with short term exposure to higher concentrations of CAW [23].
In addition, in contrast to our prior studies with short term CAW treatment, prolonged CAW treatment reduced the Aβ plaque burden in the cortex of 5xFAD mice. Although our previous studies have shown that two weeks of CAW treatment does not affect cortical Aβ accumulation [23, 31], others have reported an Aβ-reducing effect of long-term Centella asiatica treatment in the PSAPP mouse model of Aβ accumulation [52]. In that study, Dha-nasekaran et al. showed that 8 months of treatment with an ethanol extract of Centella asiatica administered in the chow selectively reduced Aβ accumulation in the hippocampus but not cortex of female animals. Interestingly in the present study, utilizing a water extract of the plant, administered for 4 months in a different mouse model of Aβ accumulation, we observed a decrease in Aβ accumulation in both the hippocampus and cortex of female mice but just the cortex of male mice. This sex difference could be in part because of the fact that control treated male 5xFAD mice had substantially lower hippocampal Aβ plaque area than their female counterparts which perhaps could affect the detection of a further reduction. We have observed this reduced plaque load in male 5xFAD mice in our previous studies as well [23]. Importantly, however, these effects on plaque burden do not appear to be necessary for cognitive enhancement as we saw similar improvements in both sexes with CAW treatment in both hippocampal and cortically mediated tasks.
In addition to these histological sex differences, variability in the magnitude of response to CAW was also evident in some of the behavioral and gene expression results. This is in line with our previous work with CAW in models of aging and AD which likewise showed differing magnitudes of effect in male and female animals [23, 27]. It is unclear what precisely accounts for these differing responses. One possibility is that differential expression of NRF2 or the 5xFAD transgenes, both of which have been reported between male and female animals [53–55], affected the response to CAW. An alternative hypothesis is that although the male and female animals consumed roughly the same volume of CAW (Supplementary Table 1) due to the size differences between male and female mice that may actually be a different level of exposure per gram body weight. Studies are underway in our laboratory to quantify the amounts of active compounds in blood and tissue of treated animals to see if this could explain the sex differences.
The cognitive changes that we observed in 5xFAD mice treated with CAW were accompanied by an increase in NRF2-regulated antioxidant response genes in the frontal cortex. This is consistent with our previous reports following the shorter CAW treatment paradigm [23, 24]. There is significant evidence in the literature of a link between antioxidant response and cognitive function in aging and AD. In fact, increased oxidative stress is considered to be an early event in AD brains [2] and diminished antioxidant capacity along with increased markers of oxidative stress are evident in both the blood and brains of AD patients [4–6]. These same alterations in ROS levels, and cognitive function, are seen in mouse models of AD [56–60]. Rodent studies also have demonstrated a relationship between antioxidant capacity and memory. In mice, aging-related cognitive decline is associated with increased oxidative damage [61], and decreased brain and plasma antioxidants [62]. Moreover, overexpressing antioxidant enzymes has been shown to improve memory in rodents [63].
Our own work suggests that NRF2 specifically plays an important role in maintaining cognitive function. We have previously reported that aged NRF2 knockout (NRF2KO) mice show even greater impairments in synaptic gene expression and cognitive function than their aged WT counterparts [46]. Additionally, NRF2 activation by various compounds has been shown to improve cognitive function in rodent models of aging as well as in older adults [64–70]. To further explore the direct role NRF2 may be playing in mediating the effects of CAW in the context of Aβ accumulation, we co-treated primary neurons from 5XFAD mice with CAW and the NRF2 inhibitory compound ML385. We found that NRF2 inhibition blocked the antioxidant and synaptic effects of CAW. These results indicate that NRF2 activation is required for the beneficial neuronal effects of CAW in vitro and suggest a role for NRF2 in synaptic plasticity in primary neurons. This is consistent with our previous work with isolated NRF2KO neurons which also display impaired arborization and reduced synaptic gene expression [46] as well as studies from other groups that have shown a relationship between NRF2 activation and markers of synaptic plasticity, including dendritic spine density [71] and neurite outgrowth [72]. We have also recently reported that loss of NRF2 prevents the cognitive enhancing effects of CAW in a mouse model of aging which further supports a role for the transcription factor in the mechanism of action of CAW in healthy aging [34]. However, it remains to be seen whether NRF2 is similarly required for the in vivo effects of CAW in AD models. Our laboratory is currently generating 5xFAD mice in which NRF2 has been knocked out in order to better understand the specific role the transcription factor plays in maintaining synaptic density and cognitive health in the context of in vivo Aβ accumulation.
It remains unknown which of the compounds wit-hin the CAW extract is responsible for its neuroprotective and cognitive enhancing action. There is evidence that the triterpenes compounds within the plant (asiatic acid, asiaticoside, madecassic acid, and madecassoside) have beneficial effects on cognition [73–78]; however, our own group has shown that caffeoylquinic acids (CQAs) within CAW also participate in its neuroprotective and cognitive enhancing effects in models of Aβ accumulation [29, 32]. Triterpenes and CQAs have both been reported to activate NRF2 [79–84] and thus could be responsible for NRF2-mediated effects of CAW. However, our chemical analysis has shown that CAW is an extremely complex mixture containing hundreds of distinct chemical compounds [33] including many phenolic compounds, like hydroxycinnamic acids and flavonoids that have been reported to activate NRF2 in other biological systems [85–87]. Future studies that focus on individual compounds from the extract, alone or in combination, or that utilize bioassay guided fractionation to identify relevant compounds are necessary to determine which compounds are responsible for each specific effects of CAW. This will be a critical step in standardizing the extract in order to move to clinical testing in humans.