
Abstract
Background:
Recent studies show that an increased T217-phosphorylation of tau in plasma could diagnose AD at an early stage with high accuracy and high specificity, while the potential toxic role of tau T217-phosphorylation is not known.
Objective:
To study the potential toxic role of tau T217-phosphorylation.
Methods:
We performed stereotactic brain injection, behavioral testing, immunohistochemistry and immunofluorescence, western blotting, Golgi staining, in vitro recombinant tau polymerization, and other measurements.
Results:
We first constructed tau T217-wild-type (T217), T217-phospho-mimic (T217E), and T217-non-phospho-mimic (T217A) plasmids or their virus vectors on the basis of wild-type tau. We found that expressing tau-T217E induced a significantly increased tau phosphorylation at multiple AD-associated sites with inhibited proteolysis and increased cleavage/fibrillization of tau, while expressing tau-T217A abolished the above changes of tau both in vitro and in vivo. By mutating T217E on tau-P301L, a dominant mutation identified in patients with frontotemporal dementia, we did not observe significant exacerbation of tau-P301L phosphorylation and cognitive impairment although the increased tau cleavage and propagation were shown.
Conclusion:
T217-phosphorylation exacerbates wild-type tau hyperphosphorylation with aggravated tau cleavage/fibrillization and cognitive impairments, while overexpressing T217E on the basis P301L does not exacerbate tau phosphorylation or the P301L-induced cognitive deficits, although it aggravates tau cleavage and propagation.
INTRODUCTION
Intracellular accumulation of hyperphosphorylated tau is an important pathological hallmark of Alzheimer’s disease (AD) [1], the most common neurodegenerative disease affecting an increasing number of the elderly population. Growing studies demonstrated that tau hyperphosphorylation could, on the one hand, lead neurons to escape an acute apoptosis [2–5]. On the other hand, tau hyperphosphorylation at multiple sites could cause tau aggregation and thus induce a chronic neurodegeneration [6–13]. Accordingly, measuring the increased tau-pT181 in cerebrospinal fluid (CSF) or measuring the increased tau in the brain tissue by tau-PET imaging have been used for the diagnosis of AD [14–16]. The therapeutic strategies aimed at specifically removing or degrading total tau using antibodies/vaccines [17, 18], or proteolytic target chimera (PROTAC) [19–21] have been developed.
Most recently, associations of plasma tau-pT217 with tau positron emission tomography (tau-PET) was identified in early AD [22]. By establishing a highly sensitive method, it was found that tau-pT217 in CSF was outperformed tau-pT181 as a biomarker for the differential identification of AD and PET amyloid-positive patients [23, 24]. Furthermore, a head-to-head comparison study demonstrated that the increase of CSF tau-pT217 appeared earlier than the change of tau-PET markers, and earlier than the previously established phosphorylation site of tau-pT181 for AD diagnosis [24–26], and the increase of plasma tau-pT217 could be used for discriminating AD from other neurodegenerative diseases [27]. These studies strongly suggest that phosphorylation of tau at Thr217 is critical; however, it is not known why tau-pT217 is so important, and how it affects the neuronal functions.
In the present study, we also detected an increased level of tau-pT217 in the brains of 3xTg AD mice, and the increase was more significant at 3 months than 6 months. By constructing different types of Thr217 plasmids, i.e., T217 wild-type (T), T217E (E, phospho-mimic), T217A (A, non-phospho-mimic), we found that phosphorylation of tau at Thr217 increased differentially the phosphorylation level of tau at other AD-associated sites, accompanying with an inhibited tau proteolysis in cells. By in vivo overexpressing different type of AAV-tau-T217, we found that overexpressing AAV-T217E exacerbated the wild-type human tau-induced behavioral deficits with an increased tau phosphorylation/cleavage and glial activation. Interestingly, by T217 phospho-mimic mutation on the basis of tau-P310L, a dominant tau mutation identified in patients with frontotemporal dementia, we did not observe significant exacerbating effects on tau phosphorylation and cognitive functions by T217E, though a significantly increased cleavage and propagation of tau-T217E-P301L was detected.
MATERIALS AND METHODS
Plasmids, viruses, and antibodies
The plasmid p3xflag-CMV-MCS-3flag (empty vector, namely V), p3xflag-CMV- tau-T217 wild-type-3flag (namely T), p3xflag-CMV-tau-T217 mutated to Glu (namely E), p3xflag-CMV-T217 mutated to Ala-3flag (namely A), p3xflag-CMV- tau-P301L-3flag (namely L), p3xflag-CMV-tau-P301L and T217 mutated to Glu-3flag (namely LE), and p3xflag-CMV-tau-P301L and T217 mutated to Ala-3flag (namely LA) were from TsingKe Biological Technology (Beijing, China). AAV-hSyn-eGFP-2a-MCS-3flag (empty vector, namely V), AAV-hSyn-eGFP-2a-tau-wild-type-3flag (namely T), AAV-hSyn-eGFP-2a-tau-T217 mutated to Glu-3flag (namely E) and AAV-hSyn-eGFP-2a-tau-T217 mutated to Ala-3flag (namely A), AAV- hSyn-eGFP-2a-tau-P301L-3flag (namely L), AAV-hSyn-eGFP-2a-tau-P301L and T217 mutated to Glu-3flag (namely LE), and AAV-hSyn-eGFP-2a-tau-P301L and T217 mutated to Ala-3flag (namely LA) were from OBIO Technology (Shanghai, China). The antibodies used in present study were listed in Table 1. Cycloheximide (CHX, HY-12320) purchased from MCE (MedChemExpress, New Jersey, USA) was used to inhibit protein synthesis with 100μg/ml [3, 28].
Antibodies used in the present study
WB, western blotting; IF, immunofluorescence; IHC, immunohistochemistry
Animals and stereotactic brain injection
Wild-type C57BL/6 mice (2 months old, male) were from Shulaibao Biotechnology (Wuhan, China). 3xTg AD mice (stock number 004807) were from the Jackson Laboratory of the United States, in compliance with the “People’s Republic of China Animal and Plant Inspection and Quarantine Law”. Laboratory animals were managed in strict accordance with the Chinese Laboratory Animal Management Regulations. The mice were kept in the same environment with a temperature of 23°C, sufficient water, free intake, and a circadian rhythm of 12:12 h. All animal experiments were carried out following the policies for animal and human use in neuroscience research. Animal care was carried out following the provision and general recommendation of the Chinese Experimental Animals Administration Legislation. The procedure was approved by the Animal Protection and Use Committee of Huazhong University of Science & Technology.
The mice were anesthetized by isoflurane and the virus vectors were injected onto the CA3 subset hippocampus (KOPF brain stereotactic locator, Germany). The coordinates for the injection were as follows: CA3 (anterior-posterior: –1.82 mm; mediolateral: –2.2 mm; dorsoventral: –2.2 mm from bregma and dura, flat skull). The behavioral tests were carried out at 1 month after the injection by the following order, and the mice were sacrificed for the biochemical measurements after behavioral tests.
Behavioral tests
For open field (OF) test, the mouse was placed in an open field of 50 cm×50 cm×50 cm. The bottom of the box was divided into 9 parts, with the center area accounting for 50%, and the edges and corners each accounting for 50%. A video tracking system (sans, Jiangsu, China) was used to calculate the central stagnation time of mouse within 5 min. Before each mouse performs each behavioral experiment, the test bench was sprayed 75% alcohol and wiped it clean.
The novel object recognition (NOR) test was carried by following a previous report [29]. The mouse was placed in an open field of 50 cm×50 cm×50 cm, two objects were placed in the corner of the same side, and the mouse was allowed to explore freely for 5 min. One of the objects was changed after 24 h and the mouse was free to move around. When the mouse is within 3 cm of the object, the experimenter recorded the exploration preference of the object within 5 min, and used the video tracking system (sans, Jiangsu, China) to test the mouse memory preference ability. At the top of center there is a camera to record distance, time, and position. Before each mouse performs each behavioral experiment, the test bench was sprayed with 75% alcohol and wiped clean.
The Morris water maze (MWM) test was carried out as described in a previous study [30]. For spatial learning, the mice were trained in a circular pool with a diameter of 120 cm and a wall height of 60 cm. The pool was artificially divided into four quadrants. The spatial learning and memory capabilities of mice are recorded through the camera video tracking system above the center of the pool (Chengdu Taimeng Software Co. Lid, China). The mice were allowed to acclimate for a week before the experiment. During the learning experiment, the mouse was gently placed in the pool facing the wall and allowed to swim freely. The time for the mouse to find the platform was recorded as the latency, and the mouse was allowed to stay on the platform for 30 s. If the mouse fails to find the platform within 60 s, the experimenter gently guided the mouse to the platform and let the mouse stay on the platform for 30 s. The learning experiment was repeated 3 times a day. The interval between two training sessions for the same mouse was at least 30 min. The mouse was placed in the different quadrant of the pool each time, but not in the platform quadrant; it lasted for 5 days.
48 h after the last training, the spatial memory ability was tested by removing the platform. The time for the mouse to reach the previous platform position was recorded as the latency, and the times to cross the platform area within 60 s was recorded as target platform crossings, and the stay time of the mouse in the target quadrant was recorded as time in target quadrant.
The fear conditioning (FC) test was carried out as described previously [31, 32]. The equipment is a box with a soundproof door measuring 17 cm×17 cm×25 cm. The bottom of the box can be artificially controlled to give electric shocks. The camera above the center of the box can record movement trajectory, distance, and time. The mouse was placed in the box to acclimate for 3 min, then a 2 s plantar electric shock (100 mA, 1 min interval) was repeated 3 times, and then the mouse was placed back into its cage. After 24 h, the mouse was put back into the box, and the freezing time of the mouse was recorded within 3 min through a video tracking system (sans, Jiangsu, China). When the mouse’s body deviates from the original position within 10%, it was defined as freezing. Before performing each behavioral experiment on each mouse, the test bench was sprayed with 75% alcohol and wiped it clean.
Immunohistochemistry and immunofluorescence
The abdomen of the mouse anaesthetized with isoflurane was fixed upward and then fixed by cardiac perfusion with normal saline (NS) and 4% paraformaldehyde (PFA). After the fixation was complete, the brain was taken, soaked in 4% PFA for 24 h, the brain was dehydrated in 30% sucrose-PBS for 48 h, and then cut into 30μm coronal sections. According to the previous method [33], immunohistochemistry (IHC) and immunofluorescence (IF) experiments were performed. The slices were incubated with primary antibody over night at 4°C. Incubated with secondary antibody for 1 h at 37°C, then the slices were washed and sealed with neutral resin (IHC) or 50% PBS (IF). The images were taken using an ordinary optical microscope (Nikon, Tokyo, Japan) or Automatic section scanning system (Olympus. Tokyo, Japan) for IHC or using a confocal microscope for IF (Carl Zeiss LSM800).
Western blotting
Western blotting was performed by the methods established in our laboratory [2]. The hippocampus were separated from the mouse brain, weighed and homogenized in buffer containing Tris x Cl (pH 7.6) 10 mmol/L, PMSF 1 mmol/L mixture (2 mg/L each of aprotinin, leupeptin and pepstain A) and cocktail 1 mmol/L according to 1 : 10, then three volumes of the homogenized tissue were added to one volume of lysis buffer containing Tris×Cl (pH 7.6) 200 mmol/L, 8% sodium dodecyl sulfate (SDS), 40% glycerol, and was boiled at 95°C for 10 min, decomposed by ultrasonic. The protein concentration was measured, then the lysates were stored in –20°C. Before using, bromophenol blue-β-mercaptoethanol mixture (1 : 3) was added and boiled at 95°C for 10 min. The sample was loaded according to the concentration and performed SDS-polyacrylamide gel electrophoresis (10% gel) separation. After the separation, the gel was transferred to the nitrocellulose (NC) membrane. After that, the NC membrane was blocked with 5% skim milk for 1 h and incubated with the primary antibody overnight at 4°C. The membrane was washed with 1 x TBST for 3×5 min the next day, and incubated with the Odyssey secondary antibody for 1 h at room temperature, then washed again with 1 x TBST for 3×5 min and imaged by Odyssey (LI-COR Biosciences, USA), finally quantitatively analyzed by ImageJ software.
Golgi staining
The kit (FD Rapid GolgiStainTM Kit, PK401) was purchased from FD Neurotechnologies, Inc. The mouse was anesthetized and the brain was soaked in the AB mixture (1:1) solution. After 24 h, the brain was placed into new AB solution and soaked for one month. During this period, it was gently shaken twice a week. One month later, the brain was placed into C solution and soaked for 3–7 days, sliced, 100μm, patched, and dried. The brain slice was stained by follows: washed with ddH2O 2×4 min; soaked by mixture (D solution: E solution: ddH2O = 1:1:2) for 10 min; washed with ddH2O for 2×4 min; dehydrated in 50%, 75%, 95% alcohol gradients for 4 min each, then absolute ethanol 4×4 min; xylene 3×4 min; left for a few hours, sealed, and imaged with an optical microscope, counted as described previously [7, 34].
In vitro recombinant tau polymerization and measurement
As described previously [35], 10μM tau proteins were incubated at 37°C with equal molar ratio of heparin in 100 mM sodium acetate buffer (pH 7.0) containing 2 mM dithiothreitol (DTT).The extent of fibrillization was assessed at different time points by Thioflavin T fluorescence assay diluted in 100 mM glycine buffer (pH 8.5) and fluorometric readings were taken at 450 nm (excitation) and 510 nm (emission). The assay was performed in triplicate.
Statistical analyses
The data were presented as mean±SEM. One-way or two-way ANOVA with Turkey post hoc tests and GraphPad Prism 8 software were used for multi-group comparison. The statistically significance was set at p < 0.05.
RESULTS
Level of T217-phosphorylated tau is increased in the hippocampus of 3xTg AD mice
We first detected the phosphorylation level of tau-T217 in the hippocampus of the 3xTg AD mice. By western blotting, we observed that the level of tau-pT217 in the total hippocampal extracts was significantly increased when compared with the age- and sex-matched controls (Fig. 1A, B), and the increase was more prominent at 3 months rather than 6 months. Immunohistochemical staining data confirmed the elevation of tau-pT217 in the hippocampal subsets, in which the increase at CA3 was significant at 3 months of age but no difference at 6 months mainly due to an age-dependent elevation of tau-pT217 level in the control group (Fig. 1C-I). These data together suggest that tau-pT217 elevation could be an early marker of AD, and it also increases age-dependently in normal mice.
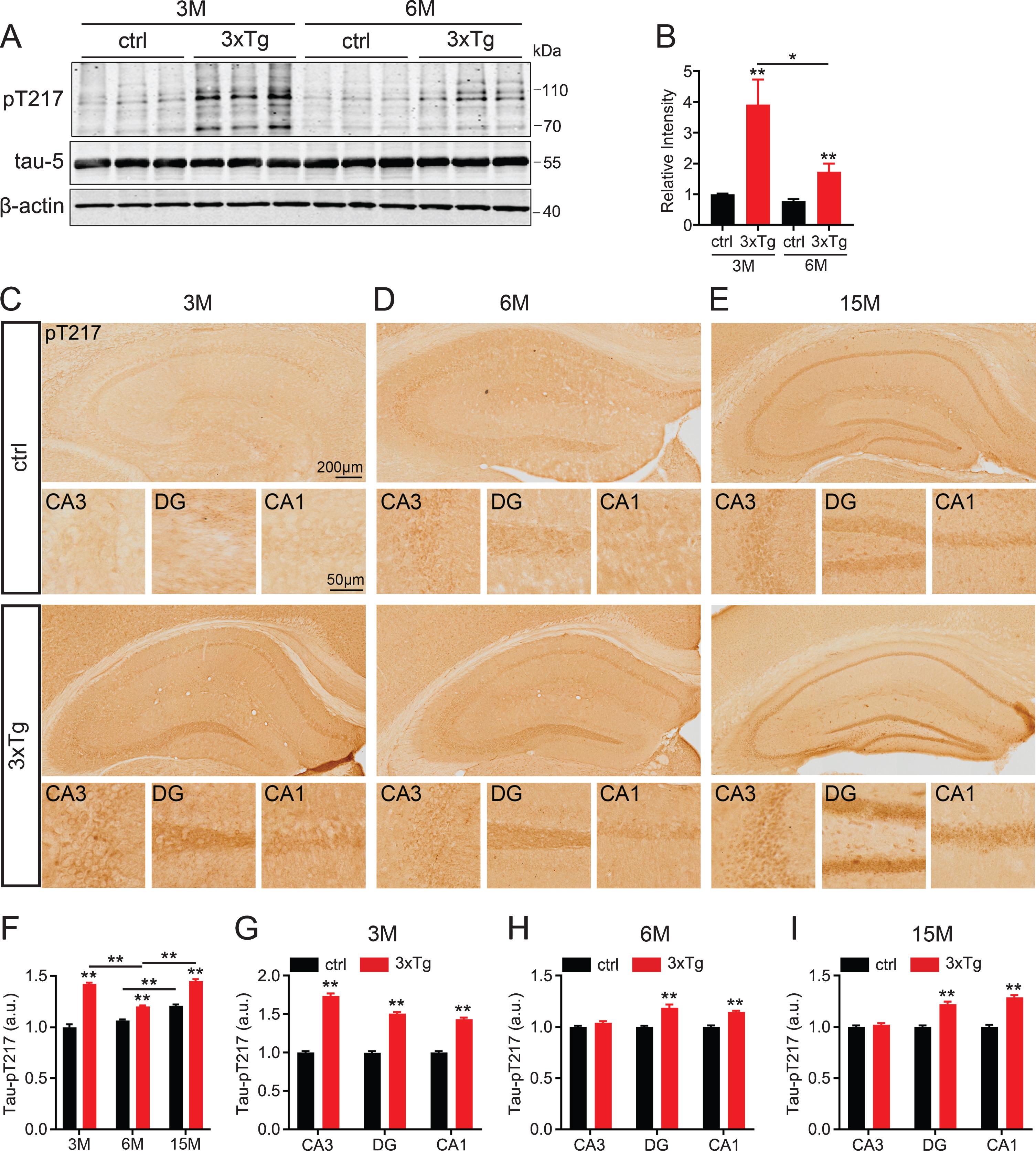
Phosphorylation level of tau-T217 is increased in the hippocampus of 3xTg AD mice. A, B) Protein level of tau-pT217 increased in the hippocampus extracts of 3xTg mice at 3 months and 6 months of age, and significant increase was shown at 3 months by western blotting. Data were expressed as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. *p < 0.05, **p < 0.01. C-I) Protein level of tau-pT217 in the hippocampal subset of 3xTg mice (F) detected by immunohistochemical staining at 3 months (C and G), 6 months (D and H), and 15 months (E and I). Scale bar, whole hippocampus, 200μm, subset, 50μm. n = 3 hippocampal slices in each group, two-tail t-test. Data were expressed as mean±SEM, n = 4 biological replicates each group, one-way ANOVA. **p < 0.01.
T217-phosphorylation on wild-type tau enhances tau phosphorylation at other sites with inhibited tau degradation and increased fibrillization in vitro
To explore the toxic effects of tau-T217 phosphorylation, we constructed T217-phosphorylation mimic (tau-T217E) and the non-phosphorylation mimic (T217A) plasmids. After transfected the plasmid in HEK293 cells for 48 h, we found that expressing tau-T217E significantly increased tau phosphorylation level at other sites, including T205, S214, T231, S262, and S396 (Fig. 2A, B). We also measured tau degradation by treated the cells with CHX for 3, 6, 9, 12, 18, and 24 h started at 48 h after the transfection. We observed that expressing tau-T217E inhibited tau degradation compared with wild-type tau and tau-T217A groups (Fig. 2C, D). To further verify whether T217 phosphorylation affects tau aggregation, we purified tau-T217, tau-T217E, and tau-T217A and measured tau aggregation in vitro. Compared with tau-T217 and tau-T217A groups, a significantly enhanced aggregation of tau-T217E was shown by Thioflavin T staining (Fig. 2E). These data suggest that tau phosphorylation at T217 increases tau phosphorylation at multiple other sites and promotes tau aggregation and fibrillization.
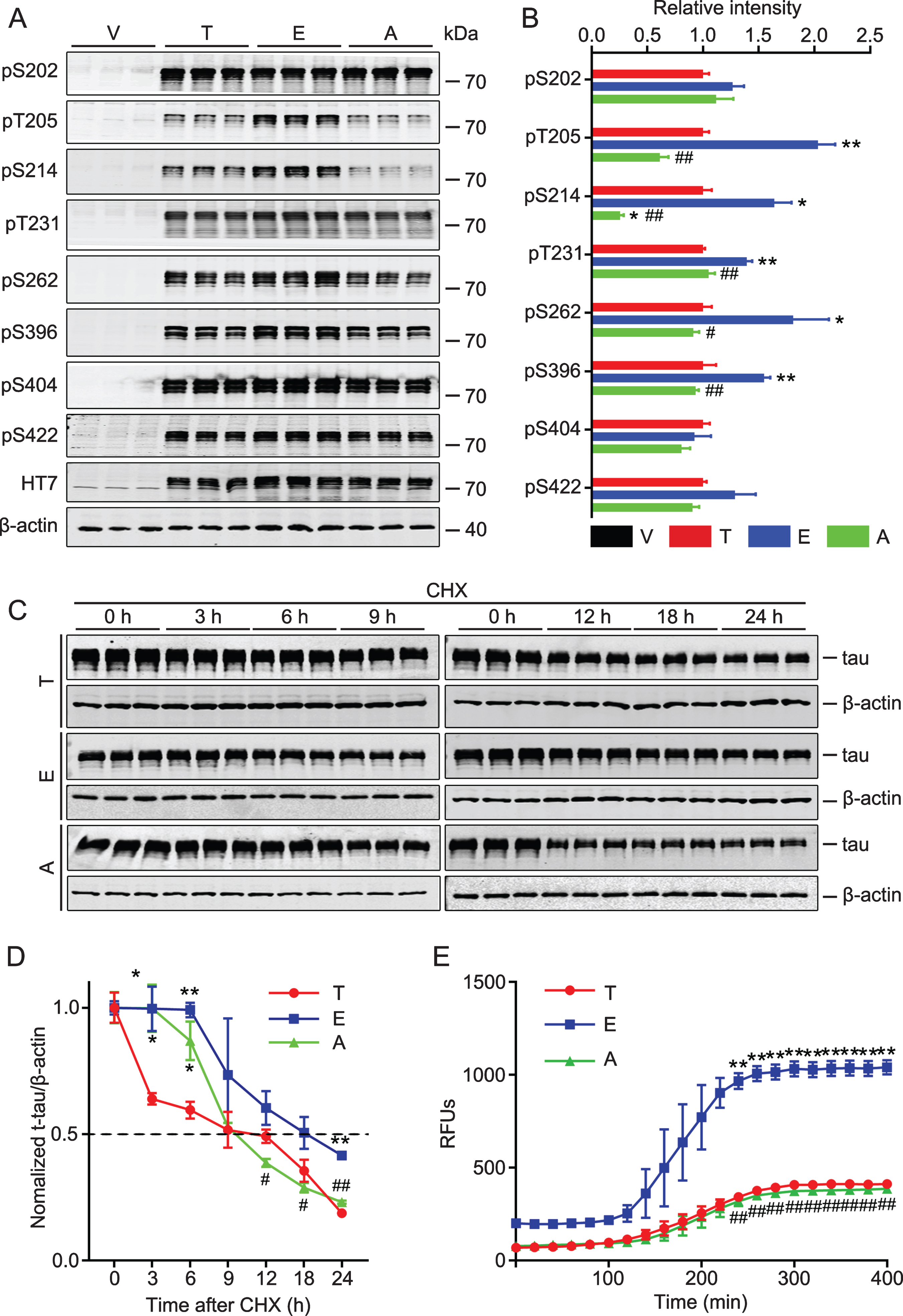
Overexpressing tau-T217E increases tau phosphorylation at other sites with inhibited tau degradation and increased fibrillization in vitro. A, B) HEK293 cell was transfected with wild-type tau (T), tau-T217E (E), tau-T217A (A), or the empty vector (V) for 48 h. Expressing tau-T217E increased tau phosphorylation at multiple AD-associated sites while expressing T217A restored tau phosphorylation level detected by Western blotting. Data were presented as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus T; #p < 0.05, # #p < 0.01, versus E. C, D) CHX was added to block new protein translation at 48 h after plasmid transfection, and the most significantly inhibited proteolysis in tau-T217E group was detected by western blotting at different time points as labeled. The dotted line indicates the half-life of each group. Data were presented as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus T; #p < 0.05, # #p < 0.01, versus E. E) Significant fibrillization of tau-T217E was detected by Thioflavin-T at 450 nm excitation and 510 nm emission respectively. Data were presented as mean±SEM, n = 3 for each time point, one-way ANOVA. **p < 0.01, versus T; # #p < 0.01, versus E.
T217-phosphorylation on wild-type tau exacerbates tau pathologies with aggravated spine and cognitive damages in mice
To explore the pathological effects of tau-T217E in vivo, we infused AAV-Vector, AAV-tau-T217, AAV-tau-T217E, or AAV-tau-T217A into the unilateral hippocampal CA3 subset of C57 mice. After one month, we measured tau phosphorylation/cleavage, dendrite number, and behavioral changes of the mice. We observed that overexpressing tau-T217E significantly increased the phosphorylation level of tau at T181, T205, S214, T231, S262, and S422 compared with wild-type tau, while overexpressing tau-T217A almost restored the phosphorylation to the wild-type tau level (Fig. 3A, B). We also observed that overexpressing tau-T217E remarkably increased the levels of truncated tau-N368 (AEP-cleaved tau) [33] and tau-C3 (caspase-cleaved tau at Asp421) [36] compared with tau-T217, and the level of the cleaved tau was almost restored to the wild-type level in tau-T217A group (Fig. 3A, B). Overexpressing tau-T217E only slightly increased the level of T22-positive tau (the oligomer tau) [37] (Fig. 3A, B). By Golgi staining, we observed that all tau-expressing groups showed a significantly reduced number of neuronal dendritic spine in hippocampal CA3 subset, and the reduction in tau-T217E group was most significant and the spine number in tau-T217A group was restored to the level of the wild-type tau group (Fig. 3C, D).
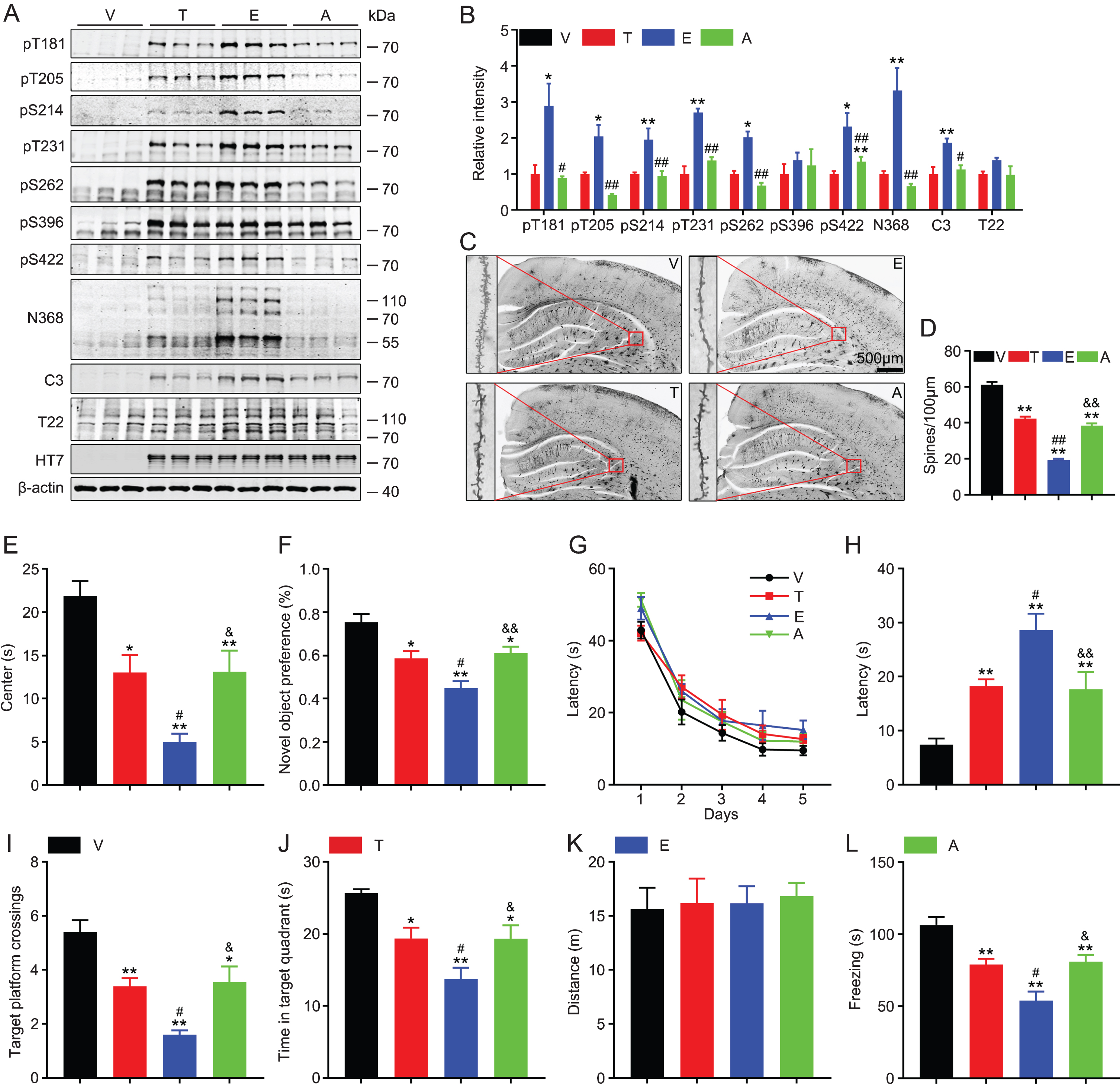
Overexpressing tau-T217E exacerbates tau pathologies and aggravates tau toxicity in inducing cognitive deficits in mice. The eGFP-non-fused virus constructs including wild-type tau (T), tau-T217E (E), tau-T217A (A), or the empty vector (V) was infused into the unilateral hippocampal CA3 subset of 2-month-old C57 mice for 1 month, and then the following parameters were measured. A, B) Overexpressing tau-T217E increased tau phosphorylation and cleavage (tau N368 and tau C3) without affecting tau oligomerization (T22), while overexpressing T217A restored tau phosphorylation and cleavage to wild-type tau level detected by western blotting. Data were presented as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus T; #p < 0.05, # #p < 0.01, versus E. C, D) Overexpressing all three types of tau induced significant spine reduction and the most significant reduction was detected in T217E group measured by Golgi staining. Scale bar, 500μm. Data were presented as mean±SEM, at least 20 neurons from three to four mice per group were analyzed. **p < 0.01, versus V; # #p < 0.01, versus T; &&p < 0.01, versus E. E) Overexpressing all three types of tau induced significantly decreased duration in center and the most significant reduction was detected in T217E group measured by open field test. Data were presented as mean±SEM n = 9∼12 each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus V; #p < 0.05, versus T; &p < 0.05, versus E. F) Overexpressing all three types of tau induced significantly decreased preference toward novel object and the most significant decrease was detected in T217E group measured by novel object test. Data were presented as mean±SEM, n = 9∼12 each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus V; #p < 0.05, versus T; &&p < 0.01, versus E. G-K) Overexpressing all three types of tau did not affect learning ability during 5 days training trials by water maze test, but significantly impaired spatial memory during probe trial carried out at day 7 by removed the hidden platform, and the most significant effect was shown in T217E group. No difference in distance moved among groups. Data were presented as mean±SEM, n = 9∼12 each group, two-way ANOVA for G, one-way ANOVA for others. *p < 0.05, **p < 0.01, versus V; #p < 0.05, versus T; &p < 0.05, &&p < 0.01, versus E. L) Overexpressing all three types of tau induced similarly decreased freezing time and the most significant decrease was detected in T217E group measured by fear conditioning test. Data were presented as mean±SEM, n = 9∼12 each group, one-way ANOVA. **p < 0.01, versus V; #p < 0.05, versus T; &p < 0.05, versus E.
In the open field test, all tau-overexpressing groups showed decreased stay in the center compared with the empty vector group (Fig. 3E). In novel object recognition test, all type tau-overexpressing mice showed decreased preference toward novel object (Fig. 3F). In Morris water maze test, no significant difference was detected in latency to find the hidden platform among four groups during the learning phase (Fig. 3G). During memory test carried out at day 7, the latency to the previous target area was significantly increased in all tau-overexpressing groups (Fig. 3H). Similar results were also shown for the times crossed in the target platform (Fig. 3I) and the time stayed in the target quadrant (Fig. 3J). No difference in traveled distance (motor ability) was detected (Fig. 3K). In fear-conditioning test, all tau-overexpressing mice showed shorter freezing time than the empty-vector control group (Fig. 3L). During the above behavioral testes, we also observed that the changes were more significant in tau-T217E group than wild-type tau and tau-T217A group, and the T217-non-phosphorylation mutation to 217A restored the behavioral performance of the mice (Fig. 3E-L). These data indicate that overexpressing different types tau in hippocampal CA3 induces cognitive deficit and T217 phosphorylation exacerbates tau-induced cognitive deficits in mice.
T217-phosphorylation on wild-type tau exacerbates tau-induced glial activation without affecting tau propagation
To measure the effect of T217 phosphorylation on tau propagation, we infused AAV-vector, or AAV-tau-wild-type, or AAV-tau-T217E, or AAV-tau-T217A with non-fused-eGFP into the unilateral hippocampal CA3 subset of C57 mice. After one month, the expression of human tau was confirmed by immunofluorescence co-staining of HT7 and GFP in the ipsilateral hippocampal CA3 subset, but no HT7 signal was detected in the contralateral hippocampus (Fig. 4A), suggesting no tau transmission from ipsilateral to the contralateral hippocampus. Note that HT7 only reacts with human tau. Then we measured glial activation by immunofluorescence staining using IBA1 (microglial marker) and GFAP (astrocytes marker) respectively. All tau-overexpressing groups showed microglial activation in both ipsilateral and contralateral hippocampal CA3, and the activation was most significant in tau-T217E group, and T217 mutation into the non-phosphorylation 217A restored IBA1 to normal control level (Fig. 4B, C). For astrocytes, overexpressing tau only induced astrocytes activation in ipsilateral side with similar feature as IBA1, but without affecting the contralateral side (Fig. 4D, E). These data indicate that wild type tau phosphorylation at T217 does not induce tau propagation though it exacerbates microglial activation at the contralateral side.
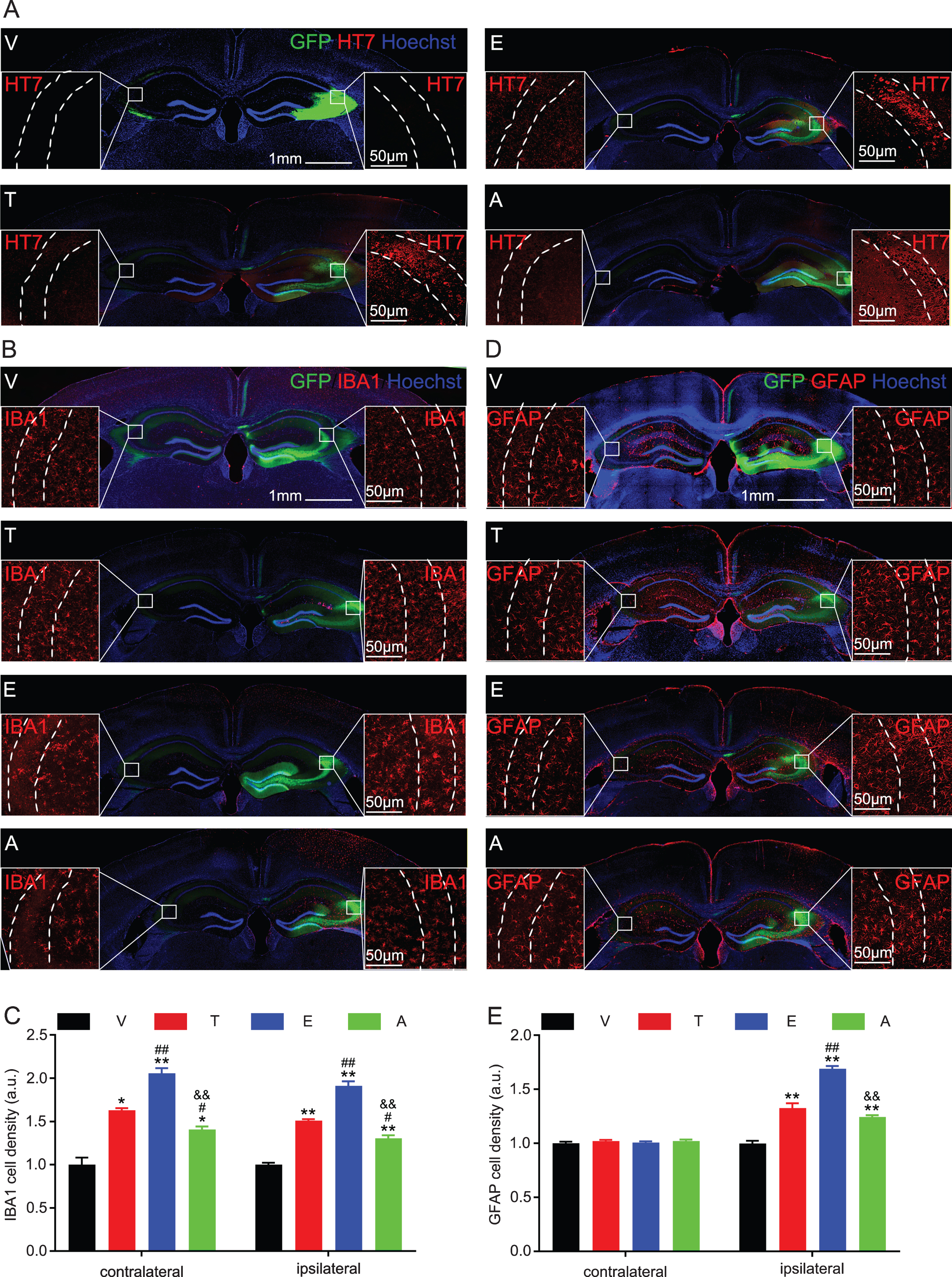
Overexpressing tau-T217E activates glial cells in ipsilateral or/and contralateral CA3 subset. The eGFP-non-fused virus constructs including wild-type tau (T), tau-T217E (E), tau-T217A (A), or the empty vector (V) was infused into the unilateral hippocampal CA3 subset of 2-month-old C57 mice for 1 month, and then tau transmission and glial activation were measured by immunofluorescence co-staining. A) Tau expression at the ipsilateral hippocampal CA3 was confirmed by HT7. Scale bar, hippocampus, 1 mm, enlarged CA3, 50μm. B-E) Microglial and astrocyte activation at ipsilateral or contralateral hippocampus were detected by IBA1 or GFAP, and the activation was most significant in T217E group. Data were presented as mean±SEM, n = 3 hippocampal slices from 3 mice in each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus V; #p < 0.05, # #p < 0.01, versus T; &&p < 0.01 versus E. Scale bar, 50μm (CA3) and 1 mm (hippocampus).
T217-phosphorylation on tau-P301L does not exacerbate phosphorylation and cognitive impairment though it promotes tau cleavage and propagation
Then we studied whether T217-phosphorylation in tau-P301L exacerbates the toxicity of tau-P301L by constructing tau-P301L (L), tau P301L-T217E (LE), tau-P301L-T217A (LA), and their AAV-constructs, respectively. After transfection the plasmids in the HEK293 cells for 48 h, we observed that expressing three types of tau did not induce significant elevation of tau phosphorylation at S202, T205, S214, T231, S262, and S396 (Fig. 5A, B).
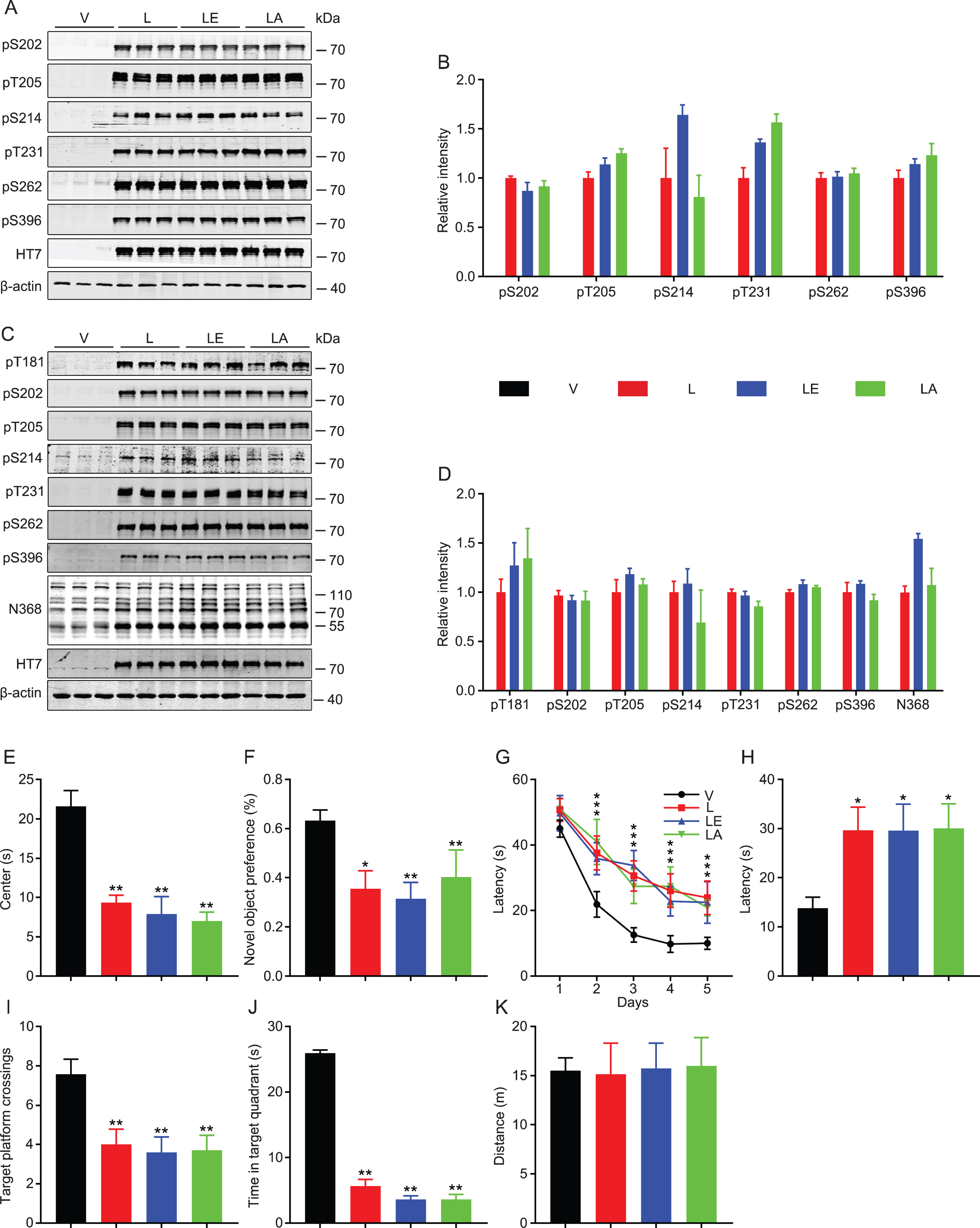
Mutating T217E on P301L does not significantly exacerbate tau-P301L-induced pathologies and cognitive deficits. A, B) Tau-P301L (L), tau-P301L-T217E (LE), tau-P301L-T217A (LA), or the empty vector (V) was transfected in HEK293 cells for 48 h, and the phosphorylation level of tau was measured by western blotting. Expressing T217E did not significantly affect tau phosphorylation. Data were presented as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. C, D) The eGFP-non-fused AAV-constructs tau-P301L (L), tau-P301L-T217E (LE), tau-P301L-T217A (LA), or the empty vector (V) were infused into the unilateral hippocampal CA3 subset of 2-month-old C57 mice for 1 month, and then tau phosphorylation and cleavage were measured by western blotting. Expressing T217E remarkably increased tau cleavage (N368) without significantly affecting tau phosphorylation in mice. Data were presented as mean±SEM, n = 3 biological replicates each group, one-way ANOVA. E-K) Overexpressing all three types of tau in unilateral hippocampal CA3 for one month induced similar degree behavioral deficits measured by open field test (E), novel object recognition test (F), water maze test (G-K). Data were presented as mean±SEM, n = 7∼10 each group, two-way ANOVA for G, one-way ANOVA for others. *p < 0.05, **p < 0.01, versus V.
By overexpressing AAV-L, AAV-LE, and AAV-LA virus in mouse unilateral hippocampal CA3 for one month, we observed that overexpressing tau-P301L-T217E remarkably increased the level of cleaved tau-N368 without affecting its phosphorylation at T181, S202, T205, S214, T231, S262, and S396 (Fig. 5C, D). We observed that overexpressing all three types of tau induced similar level of behavioral abnormalities measured by open field test, novel object recognition test, and Morris water maze test, respectively (Fig. 5E-J) without affecting motor function (Fig. 5K).
After overexpressing the eGFP-non-fused tau virus constructs into hippocampal CA3 for one month, we confirmed different types of exogenous human tau-P301L expression by HT7 immunofluorescence staining in the ipsilateral side of the hippocampal CA3 (Fig. 6A). Interestingly, we detected HT7-positive signal on the contralateral hippocampal CA1 of tau-LE and tau-L groups. The results showed that HT7 signal was more significant in tau-LE than tau-L, while the weakest signal was shown in tau-LA group (Fig. 6A, B), suggesting that T217 phosphorylation exacerbated tau-P301L propagation. By immunofluorescence co-staining with IBA1 and GFAP, we also observed that T217 phosphorylation exacerbated tau-P301L-induced glial activation (Fig. 6C-F).
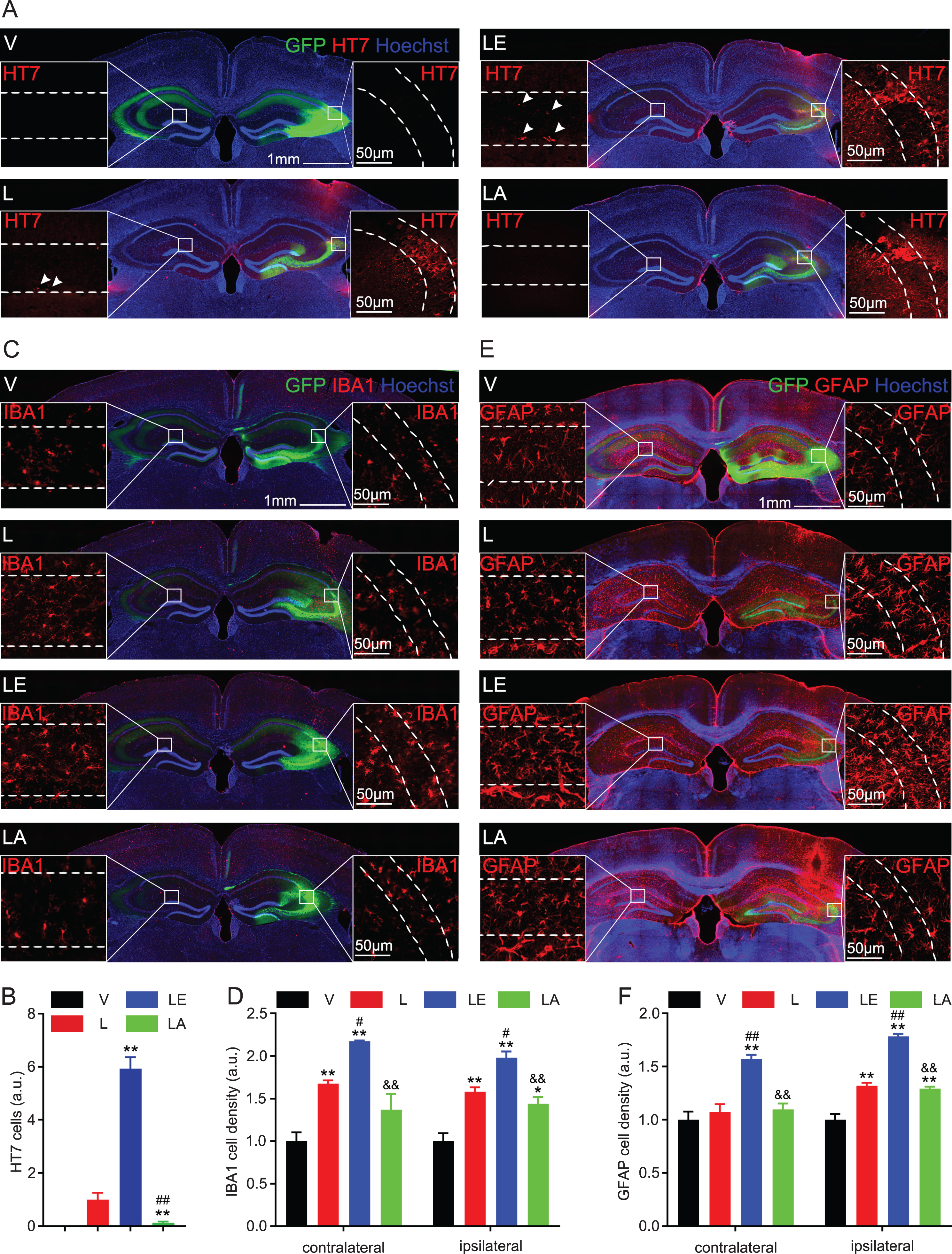
Mutating T217E on P301L promotes tau transmission and glial activation. A, B) The eGFP-non-fused AAV-constructs tau-P301L (L), tau-P301L-T217E (LE), tau-P301L-T217A (LA), or the empty vector (V) were infused into the unilateral hippocampal CA3 subset of 2-month-old C57 mice for 1 month. The ipsilateral expression of different types tau was confirmed by HT7, and the contralateral CA1 propagation of HT7-positive tau in tau-L and tau-LE groups was also detected, and the propagation in tau-LE group was more significant than tau-L group. Data were presented as mean±SEM, n = 3 hippocampal slices from 3 mice in each group, one-way ANOVA. **p < 0.01, versus T; # #p < 0.01, versus E. Scale bar, 1 mm (hippocampus), 50μm (CA3 and CA1). C-F) Microglial and astrocyte activation at both ipsilateral and contralateral hippocampus were detected by IBA1 or GFAP, and the activation was most significant in T217E group. Data were presented as mean±SEM, n = 3 hippocampal slices from 3 mice in each group, one-way ANOVA. *p < 0.05, **p < 0.01, versus V; #p < 0.05, # #p < 0.01, versus T; &&p < 0.01 versus E. Scale bar, 1 mm (hippocampus), 50μm (CA3 and CA1).
DISCUSSION
Intraneuronal accumulation of tau is one of the hallmark pathologies observed in the brain of AD patients [25, 38–40], and the number of neurofibrillary tangles mainly composed of hyperphosphorylated tau is positively correlated with neurodegeneration and cognitive deficits [7, 8]. Tau is a microtubule associated protein, with the generally recognized functions in promoting microtubule assembly and maintaining the stability of the microtubules. Recent studies indicate that tau protein possesses acetylation transferase activity which can promote self-and β-catenin acetylation, respectively [3, 42]. Tau phosphorylation also regulates cell viability, i.e., it confers cells resist to the acute apoptosis with the mechanisms involving preserving β-catenin, while the intracellular accumulation of the hyperphosphorylated tau promotes a chronical neurodegeneration [2]. With hyperphosphorylation, tau proteins are more vulnerable to other types of posttranslational modifications, such as acetylation, sumoylation, etc., and the comprehensive interaction of tau hyperphosphorylation with these additional posttranslational modifications inhibits tau proteolysis and promotes tau accumulation [6, 43]. Therefore, hyperphosphorylation is the primary cause of tau accumulation and the related chronic neurodegeneration.
There are over 85 putative phosphorylation sites contained in human wild-type full-length tau protein, among them, over 45 have been identified to be hyperphosphorylated in the AD brains [44–46]. Therefore, finding out critical toxic phosphorylation sites is important for diagnosis and drug development of AD and the related tauopathies. Recent studies demonstrated that the increased level of tau-pT181 in blood and CSF could be used as a simple, accessible, and scalable test for screening and diagnosing AD [24, 25]. Most recently, it has been reported that the phosphorylation level of tau at T217 was significantly increased in the plasma and CSF of AD patients [22–25, 27]. Compared with the previously reported markers, the increased level of tau-pT217 in the plasma appears even earlier than the increased pT181 and can better distinguish AD from other neurodegenerative diseases [24, 25]. The increased plasma tau-pT217 also appears earlier than brain tau-PET with same accuracy as CSF p-tau [27]. These data suggest that tau phosphorylation at T217 is an important early indicator for AD. However, it is not known why and how T217-phosphorylation could predict AD.
To explore the influence pT217 on other sites phosphorylation, we simulated T217 phosphorylation and non-phosphorylation mutations in human full-length wild-type tau. Both in vitro and in vivo studies demonstrated that T217-phosphorylation promoted tau hyperphosphorylation at other AD-associated sites, including T181, T205, S214, T231, S262, S396, and S422, which is consistent with the pathological process of tau [47, 48]. Simultaneously, T217-phosphorylation inhibited tau degradation. Then, we mutated T217 on tau-P301L and measured their phosphorylation level. Different from what has been observed in wild-type tau, phosphorylation mimics at T217 did not significantly increase other sites phosphorylation levels of tau-P301L. Previous studies demonstrated that P301L mutation of tau could increase tau phosphorylation [49, 50]. We speculate that the phosphorylation level of tau-P301L may be already reach to the plateau, or pT217 on P301L may produce a special confirmation that impedes tau phosphorylation at other sites, which worth of further study.
Previous studies demonstrated that overexpressing human tau in the hippocampal CA3 induced learning and memory impairments in mice [7]. Here we also observed overexpressing different types of tau proteins, including wild-type tau, T217E and T217A, or T217E/A on the bases of P301L, induced learning and memory deficits with anxiety-like behavior. Furthermore, overexpressing phospho-mimic tau-T217E exacerbated wild-type tau-induced cognitive deficits, while expressing tau-T217A rescued the cognitive functions. In consistent with the phosphorylation data, mutating T217 into E on the basis of tau-P301L did not significantly aggravate the tau-P301L-induced cognitive impairments.
Tau transmission plays an important role in causing neurodegeneration [51, 52]. Therefore, we also detected the effects of T217-phosphorylation on tau transmission on the basis of wild-type and P301L, respectively. We observed that overexpressing tau-T217E on the basis of wild-type for one month did not induce tau transmission to the contralateral hippocampus. However, overexpressing tau-T217E on the basis of P301L mutation for one month induced significant tau propagation to the contralateral hippocampus, it was more significant than tau-P301L. Previous studies demonstrated that microglial activation affects tau transmission [53–55]. In the present study, we also observed that overexpressing different types tau could activate glial cells, and the effect of tau-T217E was most significant. These data suggest that T217-phosphorylation only on the basis of P301L mutation exacerbates tau transmission, although expressing all different types of tau could activate glial cells.
Together, we found in the present study that T217-phosphorylation of tau on the basis of wild-type induces multiple other sites hyperphosphorylation and exacerbates tau-induced cognitive impairments, while T217-phosphorylation on the basis of P301L does not aggravate other sites phosphorylation nor exacerbate cognitive deficits though it exacerbates tau propagation.
Footnotes
ACKNOWLEDGMENTS
This study was supported in parts by grants from the Science and Technology Committee of China (2016YFC1305800), the Natural Science Foundation of China (91949205, 31730035, 81721005), the Special project of technological innovation of Hubei Province (2018ACA142, to J-Z.W.), and Guangdong Provincial Key S&T Program (018B030336001).
DATA AVAILABILITY STATEMENT
The data used to support the findings of this study are available from the corresponding author upon request.