
Abstract
Alzheimer’s disease (AD) is a neurodegenerative disorder in the central nervous system, and this disease is characterized by extracellular senile plaques and intracellular neurofibrillary tangles. Amyloid-β (Aβ) peptide is the main constituent of senile plaques, and this peptide is derived from the amyloid-β protein precursor (AβPP) through the successive cleaving by β-site AβPP-cleavage enzyme 1 (BACE1) and γ-secretase. AβPP undergoes the progress of post-translational modifications, such as phosphorylation and glycosylation, which might affect the trafficking and the cleavage of AβPP. In the recent years, about 10 phosphorylation sites of AβPP were identified, and they play complex roles in glycosylation modification and cleavage of AβPP. In this article, we introduced the transport and the cleavage pathways of AβPP, then summarized the phosphorylation and glycosylation sites of AβPP, and further discussed the links and relationship between phosphorylation and glycosylation on the pathways of AβPP trafficking and cleavage in order to provide theoretical basis for AD research.
Alzheimer’s disease (AD) is a common and incurable neurodegenerative disease. The disease seriously threatens human health and brings a huge economic burden to society. In 2018, there were about 50 million people who suffered from dementia in the world and 70%of them were caused by AD [1]. Therefore, the prevention and treatment of AD is of great significance to human health.
In the past several decades, researchers have carried out many investigations to identify the key factors for AD development. They have developed some hypotheses, including amyloid cascade hypothesis [2], tau hypothesis [3], and inflammatory hypothesis [4], etc. Most researchers seem to consider that two proteins called amyloid-β (Aβ) and tau are heavily involved in AD development. On one hand, soluble oligomers of Aβ might stimulate astrocytes and microglia, and over-activated astrocytes and microglia release inflammatory cytokines, leading to neuroinflammation [5]. On the other hand, aberrant level of insoluble Aβ in the brain can form senile plaques (SPs). In addition, the excessive accumulation of Aβ may cause a series of events, such as mitochondrial dysfunction and endoplasmic reticulum (ER) stress, resulting in the loss of neurons [6–8]. Therefore, the hypothesis that excessive Aβ deposition causes neuronal loss and AD development is called “amyloid cascade hypothesis”. As another key risk factor of AD, tau, a microtubule-associated protein, is a primary constituent of cytoskeletal protein of neurons and affects the development of AD. The hyperphosphorylated tau contributes to the formation of neurofibrillary tangles (NFTs) inside neurons, affecting synaptic plasticity and inducing cell death [9, 10]. The hypothesis that hyperphosphorylated tau results in neurons loss and cognitive impairment is termed “tau hypothesis”. Beyond Aβ and tau, emerging evidence suggests that neuroinflammation is widely associated with AD. The neuroinflammation contributes to the formation of SPs and NFTs. In turn, SPs and NFTs can activate the neuroinflammatory response [11, 12]. This crosstalk between neuroinflammation and SPs as well as NFTs may eventually lead to neuronal damage and apoptosis [13]. Some studies suggest that Aβ might not be the primary causative agent for the development of AD; therefore, the “amyloid cascade hypothesis” is controversial [14, 15]. AD is a kind of multifactorial disease, and the pathogenesis remains unknown. To explain this problem, the dual pathway hypothesis, which suggested that a common upstream event drives Aβ deposition and hyperphosphorylation of tau and these two convergent events induce synaptic and neuronal loss, is proposed and attempted to refine the amyloid cascade hypothesis [16].
In general, the amyloid cascade hypothesis is widely accepted in the academia. Aβ is derived from amyloid-β protein precursor (AβPP), which is successively cleaved by β-site AβPP-cleavage enzyme 1 (BACE1) [17–19] and γ-secretase [20, 21]. However, AβPP undergoes post-translational modifications (PTMs), including glycosylation [22–25] and phosphorylation [26], and these modifications can influence AβPP cleavage [27, 28]. So, it is crucial for AD research to elucidate the mechanism of AβPP’s PTMs. In the previous reports, we had summarized the glycosylation modification of AβPP and the potential roles on cleavage of this protein in AD [22]. In this review, firstly, we briefly introduce AβPP expression and its trafficking. Secondly, we primarily summarize the phosphorylation sites of AβPP, the interactive relationship between phosphorylation and glycosylation, and their roles on AβPP cleavage.
TRANSPORT AND CLEAVAGE PATHWAYS OF AβPP
An overview of AβPP
In human, the APP gene resides on the long arm of chromosome 21 at position 21.3 (21q21.3). As a type I transmembrane glycoprotein, AβPP is ubiquitously expressed in the most organs and tissues. The family of AβPP-like proteins has three numbers: AβPP, APLP1 (AβPP-like protein 1), and APLP2 (AβPP-like protein 2) [29]. Only AβPP contains Aβ domain. Because of the diverse splicing of AβPP pre-mRNA, about 10 kinds of AβPP isoforms containing around 639–770 amino acid residues are generated after translation. These isoforms are expressed in the different tissues of human. The isoforms of AβPP695, AβPP751, and AβPP770 are well known. These three isoforms show two different characteristics that: 1) the AβPP695 isoform is predominantly expressed in neurons, while AβPP751 and AβPP770 isoforms are more expressed in peripheral tissues; 2) unlike AβPP751 and AβPP770, AβPP695 lacks the Kunitz-type protease inhibitors (KPI) domain. However, what all of them have in common is that these three isoforms have a large extracellular domain, a shorter transmembrane domain, and a cytoplasmic domain [30] and can generate Aβ peptide [31, 32]. The large extracellular domain contains a cysteine-rich globular domain E1, an acidic domain AC, a spiral-rich domain E2, and an Aβ domain. The E1 domain contains a heparin-binding site and a metal ion (copper and zinc)-binding domain site. The E2 domain is a kind of coiled coil substructure formed by 6 α-helices. The cytoplasmic domain (also called intracellular C-terminal domain) contains a conserved YENPTY motif. This motif is responsible for protein-protein interaction and may play an important role on the function of AβPP and the development of AD [33–35]. Several sites of AβPP can be phosphorylated and glycosylated by various enzymes. These PTMs would affect the trafficking and the cleavage of AβPP.
Transport of AβPP
After AβPP is synthesized in the membrane-bound polyribosomes in ER, the N-terminal signal peptide of AβPP is cleaved. In turn, AβPP is transported into ER-Golgi intermediate compartment, Golgi apparatus, and trans Golgi network (TGN) [36, 37]. Finally, only about 10%of a newly synthesized AβPP is transported to plasma membrane by the secretory vesicles, while most of the AβPP resides in Golgi apparatus and TGN [38–40]. In addition, some AβPP located in Golgi apparatus are transported to lysosomes via adaptor protein-3 (AP-3). On the surface of cell membrane, AβPP is cleaved by various lyases involving in cleavage pathways of AβPP or internalized into the endosome by clathrin-coated vesicles mediated by its C-terminal motif YENPTY. Although there are various subcellular trafficking pathways of AβPP between organelles, the plasma membrane trafficking of AβPP is a key step for its cleavage [41]. When internalized into the early endosome, AβPP might be recycled back to the surface of cell membrane, or be delivered to lysosomes for degradation, or be transported to TGN in a retromer-mediated pathway for the next cycle [36, 43]. Internalization of AβPP depends on clathrin-coated vesicles and cholesterol, and the former is the main way for endocytosis of AβPP.
AβPP is also transported from neuronal cell bodies to distal terminals by fast anterograde axonal transport of system [44]. AβPP is transported into axons via kinesin-dependent anterograde transport [45]. Kinesin-1, a member of kinesin super family and a microtubule motor protein, is composed of two heavy chains (kinesin-1 motor proteins) and two light chains (kinesin-1 adaptor proteins). The kinesin-1 motor proteins, equipping with ATPase activity, provide motile power for transporting of cargoes, while the kinesin-1 adaptor proteins are involved in attachment for cargoes [46]. The interaction between kinesin-1 adaptor proteins and AβPP could regulate the axonal anterograde transport of AβPP, and AβPP may function as a kinesin-1 receptor of membrane compartment [47]. To explore the function of AβPP and the mechanism of membrane compartment in anterograde transport, Kamal et al. found that the amyloidogenic pathway of AβPP occurred in membrane compartment and this proteolysis would liberate kinesin-1 from the membrane. These results suggest that AβPP, functioning as a kinesin-1 membrane receptor, mediates the fast axonal transport of BACE1 and presenilin-1 and that Aβ production occurs in axonal membrane compartment [48]. Altogether, AβPP is an anterograde transport cargo in axon and impaired axonal transport is related to AβPP cleavage and Aβ production [49].
Intracellular transport of AβPP can be modulated by some transmembrane proteins. Here, we mainly introduced three of these transmembrane proteins: low-density lipoprotein receptor-related protein 1 (LRP1), SorLA, and Sortilin.
LRP1 can bind N-terminal KPI domain and C-terminal cytoplasmic domain of AβPP [50, 51]. As a result of lacking the KPI domain in AβPP695, the C-terminal interaction is likely to associate with brain function of AβPP695. LRP1 interacts with AβPP through Fe65 (a functional linker) in the secretory pathway, resulting in the formation of LRP-Fe65-APP trimeric complex [52]. LRP1 might function as a sorting receptor. On one hand, LRP1 facilitates a part of AβPP to delivery to the cell surface by common secretory pathway. On the other hand, the majorities of AβPP are transported by LRP1 into anterograde transport vesicles [53].
Sorting-related receptor with A-type repeats (SorLA) is highly expressed in neurons of the hippocampus, cerebellum, and cerebral cortex [54]. A study has revealed that SorLA can influence the early and late onset AD [55]. SorLA has an independent ligand-binding site termed the vacuolar protein sorting 10 protein (Vps10p) domain, which can directly bind or sequester AβPP in intracellular compartments [56]. SorLA located in Golgi compartments and endosomes strongly co-localizes with AβPP. After endocytosis, sorLA can be shuttled between endosomes and TGN by cytosolic adaptors, such as GGAs (Golgi-localizing, γ-adaptin ear homology domain, ADP-ribosylation factor-binding proteins) and PACS1 (phosphofurin acidic cluster sorting protein 1). SorLA is likely to connect with retromer (an adaptor complex) and involves in retrograde transport of AβPP [57]. SorLA can be delivered back to early endosomes when binding to adaptor protein-1 (AP-1) and GGAs; however, sorLA would be transported into reverse direction when binding to retromer. PACS1 might regulate SorLA in either direction. Compared to the anterograde transport of AβPP, the retrograde transport mediated by retromer would enhance amyloidogenic pathway of AβPP, resulting in the accumulation of Aβ [58]. Further, a study substantiates that the absence of retromer for SorLA would reduce the level of full length AβPP and increase Aβ production [59]. These results indicated that anterograde sorting of SorLA may pay a role in amyloidogenic pathway of AβPP and Aβ generation.
Sortilin is termed neurotensin receptor 3, a multiligand sorting receptor. Sortilin is expressed in neurons of the central nervous system as well as peripheral nervous system. Sortilin contains an extracellular domain, Vps10p domain, and an intracellular domain, which includes F/YXXXXF/Y, YSVL, and dileucine motifs (HDDSDEDLLE) [60, 61]. In contrast to extracellular domain, the intracellular domain is mainly related to protein sorting [62]. Sortilin participates in anterograde transport in the manner of sortilin-substrate complex[63]. Researchers have uncovered that Sortilin was also a risk factor in sporadic AD [64]. Sortilin binds AβPP (AβPP770 as an example) via head-to-head and tail-to-tail interactions in neurons. That is, AβPP1–287 and AβPP713–770 interact with sortilin78–385 and sortilin-FLVHRY sequence, respectively [65]. Sortilin, as an interaction partner of AβPP, regulates AβPP metabolism with the promotion of sAβPPα production [50]. These results indicate that the anterograde transport of AβPP modulated by sortilin would enhance nonamyloidogenic pathway in neurons.
In summary, these three proteins can affect AβPP trafficking, subcellular localization, and cleavage. However, their possibly detailed roles, especially in the interplay with glycosylation/phosphorylation of AβPP, are needed to further investigate.
Cleavage pathways of AβPP
The AβPP695 isoform is dominantly expressed in neurons but not in non-neuronal cells such as glia [66]. Its cleavage products are related to the development of AD. Therefore, in this section, we mainly introduced several processing pathways of AβPP695 and the corresponding sites cleaved by lyases. AβPP695 is cleaved through the canonical AβPP processing, including nonamyloidogenic and amyloidogenic pathways, and the noncanonical cleavage pathways.
Canonical pathways of AβPP cleavage
In the nonamyloidogenic pathway, the peptide bond between Lys612 and Leu613 (corresponding to Lys16 and Leu17 of Aβ) in AβPP695 is cleaved by ADAM10, generating an extracellular soluble N-terminal fragment (sAβPPα) and a membrane-bound C-terminal fragment (CTF-α, also called C83) [67]. Next, CTF-α is cleaved by γ-secretase complex, producing a nontoxic P3 peptide (Aβ17–40 or Aβ17–42) and an AβPP intracellular domain (AICD) [68].
In the amyloidogenic pathway, BACE1 cleaves the peptide bond between Met596 and Asp597 amino acid residues in AβPP695, generating a soluble N-terminal fragment (sAβPPβ) and a membrane-bound C-terminal fragment (CTF-β, also called C99), and then the CTF-β is cleaved by γ-secretase to release full-length Aβ1–40 (or Aβ1–42) and AICD [69]. The cleavage sites of γ-secretase locate in the middle of transmembrane domain, and the mechanism is still not fully understood. There is an opinion that γ-secretase first cleaves CTF-β fragment at multiple sites, producing various length Aβ peptides ranging between Aβ37 to Aβ49. However, γ-secretase is initially inclined to produce longer Aβ species, Aβ48 or Aβ49; the Aβ48 and Aβ49 fragments will undergo multiple successive tripeptides cleavages to produce two major Aβ species, Aβ40 or Aβ42, respectively [70].
According to “amyloid cascade hypothesis”, Aβ accumulation is a vital event to drive AD development. Therefore, inhibiting Aβ production and clearing Aβ deposition are the two major strategies for AD treatment. Many clinical trials for decreasing Aβ production via inhibiting BACE1 or elevating ADAM10 were failed due to various reasons [71, 72]. Unfortunately, the decrease of amyloid deposition after therapy with drugs or monoclonal antibodies was shifted to form soluble Aβ, which could not be cleared [73, 74]. The soluble Aβ would result in the neurotoxicity and impairment of cognitive ability [75, 76]. So, multi-target therapies may be adopted to clear Aβ species in the future.
Noncanonical pathways of AβPP cleavage
Besides the canonical nonamyloidogenic and amyloidogenic pathways, there are three alternative noncanonical cleavage pathways of AβPP695: η-secretase pathway, δ-secretase pathway, and meprin β-mediated pathway.
The η-secretase pathway occurs under physiological conditions and the development of AD. Mem-brane-type matrix metalloproteinases (MT-MMPs) play a pivotal role in regulating AβPP metabolism and amyloidogenesis. In vitro, membrane type 1-mat-rix metalloproteinase (MT1-MMP) and MT5-MMP, which have been associated with the pathophysio-logical mechanisms of AD, belong to subfamily of MT-MMPs and are regarded to have biological activity in common with η-secretase due to the same molecular weight of their cleavage products [77, 78]. The peptide bond between Asn504 and Met505 in AβPP695 is cleaved by η-secretase, generating a soluble ectodomain of AβPP (sAPP-η) and a C-terminal fragment (CTF-η). The CTF-η is further cleaved by ADAM10 or BACE1, producing the Aη-α and CTF-α peptides or the Aη-β and CTF-β peptides, respectively. The Aη-α and the Aη-β are released to the extracellular medium, and the CTF-α and the CTF-β left in the cell membrane can be further cleaved by γ-secretase, liberating AICD to the cytoplasm.
Mammalian asparagine endopeptidase (AEP) is a cysteine proteinase and usually located in lysosomes. AEP is also called δ-secretase due to its role in AβPP cleavage. AEP in the brain has a variety of physiological functions. In excitotoxic condition, AEP is activated by acidaemia, a disease of increasing serum ammonia, causing neuron apoptosis through degrading DNA enzyme inhibitor SE translocation, which binds to AβPP. The activation of AEP is a spontaneous and pH-controlled process. The AEP is activated when its N-terminal polypeptide or C-terminal polypeptide sequence is removed, and then it cleaves AβPP695 after asparagine residues. AEP cleaves AβPP695 at Asn585, generating AβPP1–585 and AβPP586–695 fragments [79]. In addition to Asn585, AEP also cleaves AβPP695 at Asn373, producing AβPP1–373, AβPP374–585, and AβPP586–695 fragments. The AβPP1–585 and AβPP1–373 fragments do not exhibit any demonstrable neurotoxic effect, while the toxic effect of AβPP1–373 fragment is specific to certain cell types. For example, AβPP1–373 fragment is toxic to primary neurons of C57BL/6J mice, resulting in cell apoptosis. However, AβPP1–373 fragment is not toxic to PC12 cells and HEK293 cells. AβPP586–695 is cleaved by BACE1 and γ-secretase to produce Aβ. Compared with full-length AβPP695 and AβPP374–695 fragment, the AβPP586–695 fragment has stronger affinity to BACE1[80, 81].
Meprin is a zinc endopeptidase, belonging to the astacin proteinase family. Meprin has two subunits (α and β), but they may not be co-expressed. If β subunits are expressed alone, they form membrane-bound homodimers. Meprin β is expressed in many organisms, such as zebrafish Danio rerio, rodents, and homo sapiens [82–85]. In human, meprin can be expressed in different organs, including brain, skin, intestine and kidney. Meprin β can be expressed under a variety of physiological and pathological conditions and is released from the surface of cell membrane by ectodomain shedding [86, 87]. Studies have shown that AβPP695 is cleaved by meprin β between Ala124/Asp125, Glu380/Thr381, and Gly383/Asp384 [88]. The cleavage pathway that AβPP is cleaved at specific sites by meprin β is known as meprin β-mediated pathway. Furthermore, it is needed to explore the exact mechanism modulating by meprin β in the cleavage of AβPP695 .
In summary, AβPP trafficking and cleavage pathways are shown in the Fig. 1, and enzymatic sites of AβPP are shown in the Fig. 2. It is needed to further clarify the roles of both phosphorylation and glycosylation on the pathways of AβPP trafficking and cleavage.
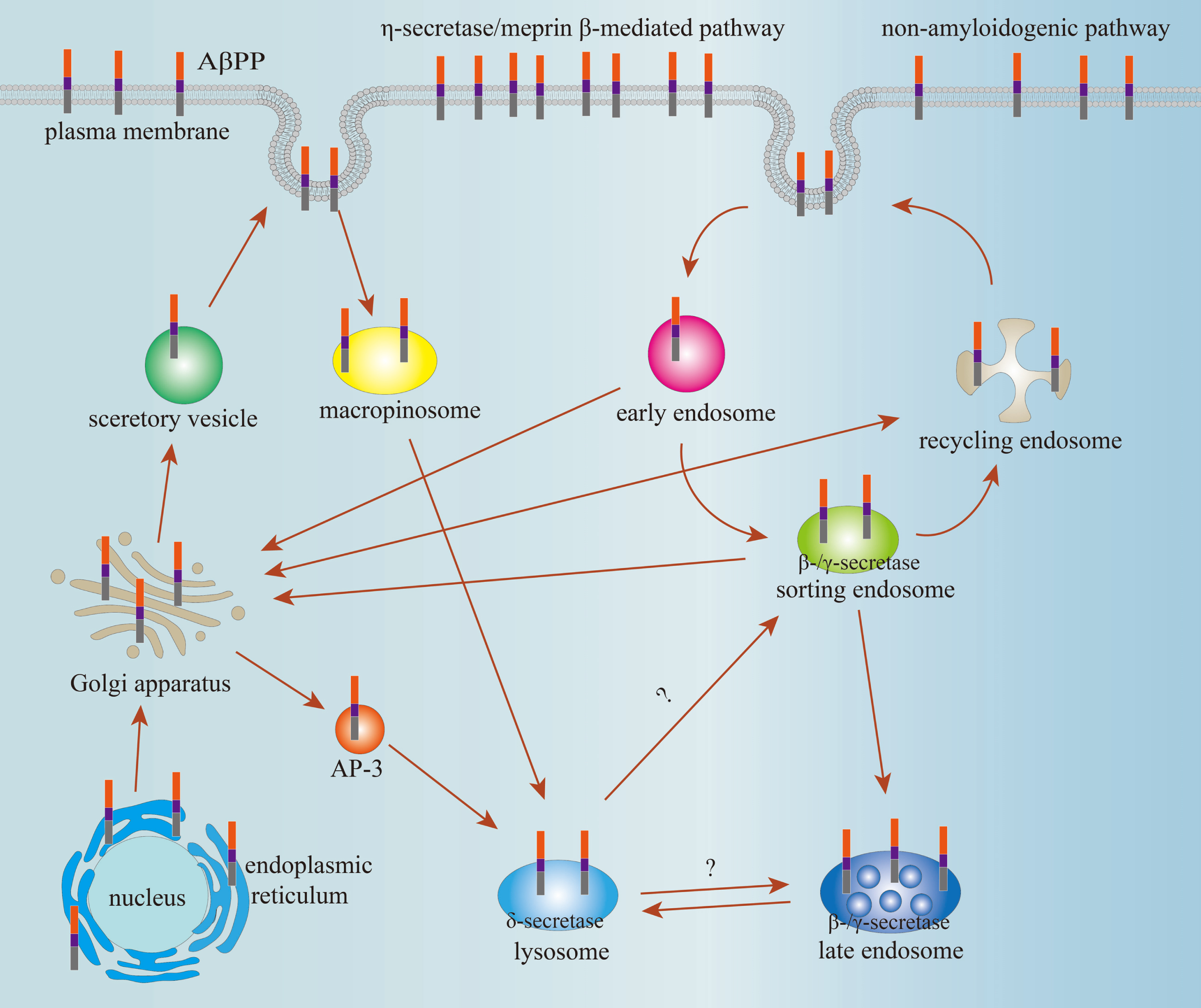
Schematic illustration of AβPP trafficking and cleavage pathways.
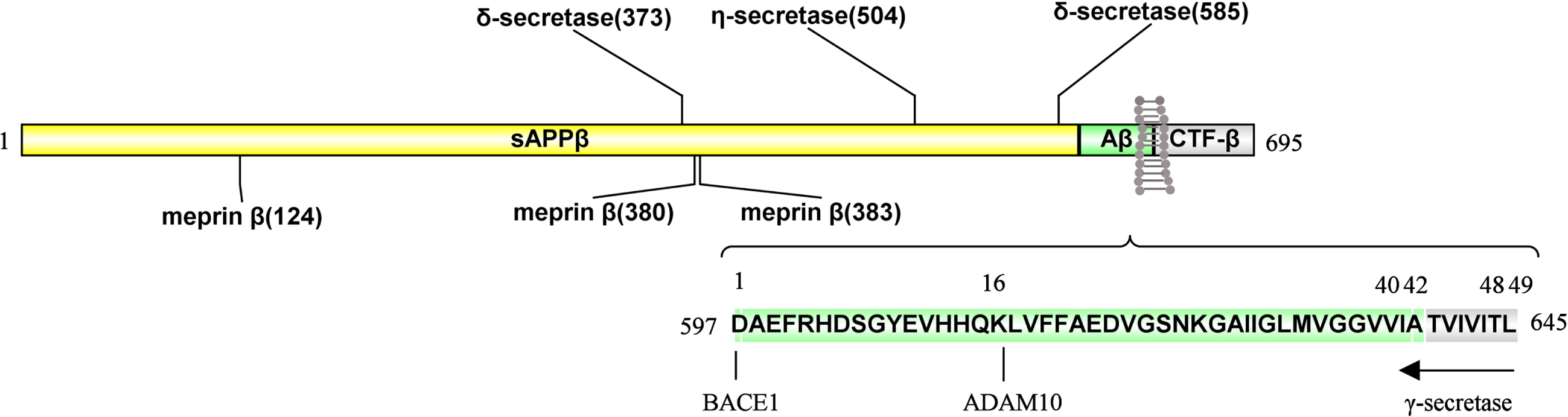
Sites of enzymatic cleavage on AβPP695.
AβPP is transported from ER to Golgi apparatus. Next, AβPP is transported to plasma membrane via secretory vesicle or recycling endosome and also transported to lysosome via AP-3. The AβPP in plasma membrane might be endocytosed into early endosome or cleaved by AβPP lyases. The AβPP in early endosome can either recycle to cell membrane via sorting endosome and recycling endosome successively, or ultimately enter the lysosomes via the late endosome to degrade, or back to the TGN. In addition, the AβPP in sorting endosome and recycling endosome also can be transported to Golgi apparatus. The AβPP cleavage by nonamyloidogenic pathway, η-secretase pathway, and meprin β-mediated pathway mainly occurs at the plasma membrane. Amyloidogenic pathway predominantly takes place in early and late endosomes, and δ-secretase pathway takes place in lysosomes. In addition, AβPP can also be transited directly into lysosomes from the cell surface via macropinosomes, a transient endocytic organelle, and be cleaved by δ-secretase, and subsequently, the fragment of AβPP586–695 is cleaved by β-secretase and γ-secretase. However, the trafficking pathway of AβPP586–695 from lysosome to endosome or other organelles is unclear.
Meprin β, δ-secretase, and η-secretase have been reported to cleave the long N-terminal extracellular domain of AβPP695. Meprin β cleaves AβPP695 at 124th, 380th, and 383th sites, respectively; the δ-secretase can cleave AβPP695 at 373th and 585th sites. Also, the η-secretase cleaves AβPP695 at the 504th site. ADAM10 cleaves peptide bond between Lys612 and Leu613 and BACE1 cleaves the peptide bond between Met596 and Asp597 amino acid residues in AβPP695. However, γ-secretase is initially inclined to produce longer Aβ species, Aβ48 or Aβ49; the Aβ48 and Aβ49 fragments will further undergo multiple successive tripeptides cleavages to produce two major Aβ species, Aβ40 or Aβ42, respectively.
EFFECTS OF PHOSPHORYLATION ON AβPP TRAFFICKING AND CLEAVAGE
Phosphorylation of proteins is the process that a phosphoryl group binds to a hydroxyl group via covalent bond at the specific serine, threonine, and tyrosine residues. Phosphorylation is regarded as a predominant post-translation modification for the regulation of protein function [89]. Abnormal protein phosphorylation is one of the key mechanisms in pathogenesis of AD. The phosphorylation of AβPP occurs in both its intracellular and extracellular portions. Phosphorylation modification impacts on the shaping of interactome, trafficking, and processing of AβPP [90]. In AD brain, researchers have identified about ten phosphorylated amino acid residues in AβPP. The long N-terminal extracellular domain has two phosphorylation sites: Ser198 and Ser206 [91]. The shorter C-terminal cytoplasmic domain has eight phosphorylation sites, known as Tyr653, Thr654, and Ser655; Thr668, Ser675, and Tyr682; Thr686 and Tyr687 [28] (Fig. 3). These ten amino acid residues can be phosphorylated in vivo and in vitro by protein kinases. Moreover, several of these phosphorylation sites have been proved to influence AβPP biological function.
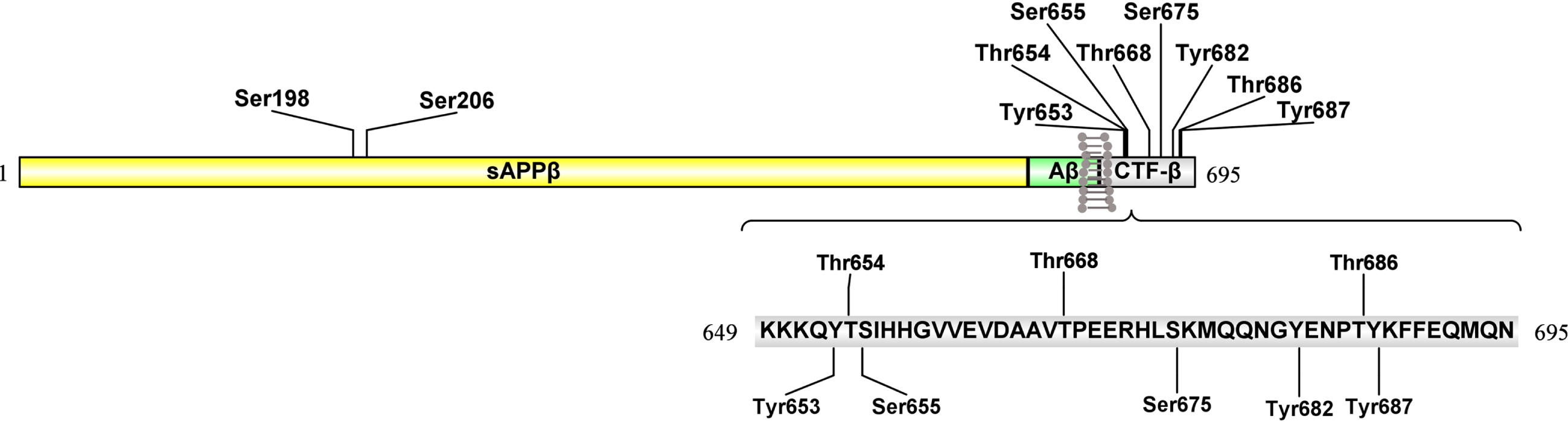
Sites of the phosphorylation on AβPP695.
Protein undergoes the dynamic process of protein phosphorylation and dephosphorylation. Several protein phosphatases (PPs) targeting to Ser/Thr residues have been identified, including PP1, PP2A and PP2B, PP2C, and PP5. PP1, PP2A, PP2B and PP5 can regulate tau phosphorylation whereas PP2C has poor efficiency to phosphorylate tau [92, 93]. Meanwhile, PP1 and PP2A are also involved in AβPP phosphorylation. PP1 can bind AβPP and Fe65 to form “AβPP-FE65-PP1” trimeric complex and dephosphorylate AβPP at Thr668 site via Fe65 dependent pathway [94]. PP2A has an effect on AβPP processing through phosphorylation at Thr668 site. In vitro, when PP2A is inhibited, Thr668 phosphorylation of AβPP is elevated and the productions of sAβPPα and sAβPPβ are increased [95].
The long N-terminal extracellular domain of AβPP695 has two phosphorylation sites, Ser198 and Ser206. In the shorter C-terminal cytoplasmic domain, eight main phosphorylation sites including Tyr653, Thr654, and Ser655; Thr668, Ser675, and Tyr682; Thr686 and Tyr687 have been reported. The kinases that phosphorylate AβPP and the effects on AβPP trafficking and cleavage can be further retrieved in Table 1.
Phosphorylated sites in AβPP695 and effects on its trafficking and cleavage
Ser198 and Ser206 phosphorylation sites
Extracellular domain of AβPP phosphorylation is related to its serine residue located in N-terminal acidic domain. AβPP will be phosphorylated when the secretory vesicles containing AβPP are transported to the surface of cell membrane. The sufficient adenosine triphosphate (ATP), released into extracellular space via secretory or synaptic vesicles, would activate ectokinases to phosphorylate downstream proteins [96]. AβPP is also phosphorylated by ectokinases at cell membrane surface. It has been shown that cAMP-dependent protein kinase-like [97] and casein kinase (CK)-like [98] enzymes have ectoprotein kinase activity. CK, including CK1 (ecto-CK1) and CK2 (ecto-CK2) isoforms, phosphorylate membrane-bound as well as soluble AβPP. Mutation of serine sites 198 and 206 to alanine (Ser198Ala/Ser206Ala) significantly decreases the phosphorylation level of AβPP695, which indicates that Ser198 and Ser206 are the phosphorylation sites of AβPP695. Phosphorylation at Ser198 and Ser206 occurs in post-Golgi secretory compartments and at plasma membrane surface. Subsequently, researchers have found that the phosphorylation of AβPP695 by ecto-CKs predominantly occurs within the acidic domain between amino acids 181 and 224 [99]. Therefore, the Ser198 and Ser206 residues might be involved in ecto-CKs-induced phosphorylation of AβPP695.
Thr654 and Ser655 phosphorylation sites
Thr654 and Ser655, situated in a highly conservative motif Y653TSI656 of AβPP695, can be phosphorylated by protein kinase C (PKC) and calcium/calmodulin-dependent protein kinase II (Ca2+/CaMK II). PKC almost just phosphorylates Ser655 residue of AβPP695, while Ca2+/CaMK II catalyzes AβPP695 phosphorylation at both Thr654 and Ser655 sites [100]. The effect of Thr654 and Ser655 phosphorylation on AβPP695 remains to be further explored. Besides these two kinases, AβPP kinase I and rho-associated coiled-coil kinase 1 (ROCK1) are also involved in the phosphorylation of AβPP at Ser655. The discovery of AβPP kinase I helps us to understand the effect of AβPP695 phosphorylation at Ser655 on its metabolic mechanism and function [101]. ROCK1 phosphorylates AβPP695 at Ser655, resulting in increase of amyloidogenic pathway and acceleration of the AD development [102]. In AβPP695, there is a conservative sequence that contains a characteristic internalization signal, YXXI. If this signal sequence is phosphorylated, it may regulate the internalization process of AβPP695 [103] and even further affect its metabolic pathway. When AβPP751 was phosphorylated by PKC at serine residue in Y709TSI712 motif (corresponding to Y653TSI656 on AβPP695), the trafficking efficiency from Golgi apparatus to lysosomes is reduced and the generation of Aβ1–42 is decreased [104]. This result also indicated that reducing the trafficking of AβPP to lysosomes may be a way to decrease the generation of Aβ1–42. In addition, phosphorylation at Ser655 was reported to enhance the binding between AβPP695 and sorting protein sorLA, resulting in retrieving of AβPP from endosome to the TGN [105].
Thr668 phosphorylation site
The phosphorylation of AβPP695 at Thr668 has been observed in the brain of human and mouse, which may be specific for nervous tissue. It is a normal process that phosphorylation occurs at Thr668, which is associated with the neurite extending and the anterograde trafficking of vesicles [106–108]. Studies have proved that phosphorylation at Thr668 of AβPP affects the cleavage of AβPP by BACE1 and γ-secretase and then further influences the generation of Aβ. Lee et al. [109] found that Thr668 phosphorylation is upregulated in AD brains and may facilitate BACE1 to cleave AβPP695, resulting in increasing of Aβ production. However, Feyt et al. [110] found that phosphorylation of AβPP695 at Thr668 increased AβPP695 cleavage by BACE1 but decreased the following cleavage by γ-secretase, ultimately resulting in the reduction of extracellular Aβ production. Another study also concludes that phosphorylated AβPP695 at Thr668 has a negative effect for the binding between phosphorylated CTFs (pCTFs) and γ-secretase and decreases the Aβ production [111]. In addition, phosphorylation of AβPP in cytoplasmic domain would alter the affinity of binding to other functional proteins [112]. For example, phosphorylation at Thr668 suppresses the interaction of AβPP695 with Fe65 by changing the structure of cytoplasmic domain containing the Y682ENPTY687 motif. This conformational change can regulate AβPP trafficking and cleavage [113]. Also, phosphorylation of Thr668 affects the association of AβPP with Fe65 and the translocation of Fe65. Fe65 that binds to AβPP would be released from cytomembrane when Thr668 residue of AβPP is phosphorylated. Then, Fe65 will be translocated into the nucleus and interacts with non-phosphorylated AICD to up-regulate gene transactivation [114].
In vivo or in vitro, the Thr668 of AβPP695 is phosphorylated by several kinases, including glycogen synthase kinase 3β (GSK-3β) [115], c-jun N-terminal kinase-3 (JNK3) [116], cell division cycle protein (Cdc2) [117], and cyclin-dependent kinase 5 (Cdk5) [66]. GSK-3β signaling pathway plays an essential role in regulating the fate of neurons and synaptic plasticity. The abnormal activity of GSK-3β may dysregulate the physiological activities of AβPP and the generation of Aβ, subsequently affecting the process of AD. Acevedo et al. [118] concluded that copper ion can increase phosphorylation of endogenic AβPP695 at Thr668 and promotes AβPP695 trafficking by GSK-3β-dependent phosphorylation pathway in SH-SY5Y cell line. Rockenstein et al. [119] found that the GSK-3β signaling pathway may have neuroprotective effects by regulating the maturation and cleavage of AβPP and these results indicated that GSK-3β might be a suitable target for treating AD.
Ser675 phosphorylation site
In the AD brain, it has been found that Ser675 of AβPP695, located at cytoplasmic domain, can be phosphorylated. Increasing evidence suggests that phosphorylation of Ser675 can modulate the AβPP processing. Phosphorylation at Ser675 increases mep-rin β-mediated cleavage but decreases ADAM10-mediated cleavage of AβPP at the plasma membrane [120]. The cleavage mediated by meprin β might result in the generation of truncated Aβ2–40/42 peptides, which are highly aggregation-prone. Therefore, in the near future, it is of importance to reveal how phosphorylation of AβPP695 at Ser675 promotes meprin β cleavage pathway of AβPP.
Tyr682 phosphorylation site
Tyr682 is the first amino acid residue resided on the Y682ENPTY687 motif, which sequence is evolutionarily conserved. In vitro, Tyr682 of AβPP695 is phosphorylated by tyrosine kinases, TrK A and c-Ab1 [121, 122]. Several cytoplasmic proteins interplay with each other by binding with AβPP. This interaction can regulate the processing of AβPP [123]. Tyr682 can interact with cytosolic proteins to provide a docking site that contributes to these proteins to combine with AβPP. Cytosolic proteins with Src homology 2 (SH2) domain recognize phosphorylated tyrosine (pTyr) residues and mediate pTyr signaling networks. The human cytosolic protein with SH2 domain can identify the presence of pTyr by binding certain sequences with phosphorylated amino acid [124, 125]. When phosphorylated at Tyr682, AβPP can bind to Shc (SH2-containing) proteins, such as growth factor receptor-bound protein 2 (Grb2) and growth factor receptor-bound protein 7 (Grb7), to form AβPP-Shc complex. This complex will further interact with other AβPP-binding proteins, such as Fe65. The SH2 domain of Grb2 directly interacts with phosphorylated Tyr682 of AβPP [90, 126]. This result indicated that the other cytosolic proteins containing SH2 domain such as Grb7 may directly interact with AβPP phosphorylated at Tyr682.
However, there is no definite conclusion on the interaction between cytosolic proteins without SH2 domain and non-phosphorylated AβPP at Tyr682. In non-phosphorylated condition, some proteins con-taining phosphotyrosine-binding domain will direc-tly bind Tyr682, including mammalian disabled 1 (mDAB-1), JNK-interacting proteins (JIPs), and Munc-18 interacting protein; Mint (X11) family proteins (mint 1, mint 2, and mint 3), Fe65, and an inhibitor of Notch termed Numb. The combination between these cytoplasmic proteins and Tyr682 of AβPP activates the downstream signaling pathway to regulate cleavage and trafficking of AβPP [127–129]. So, Tyr682 regulates the transport and cleavage of AβPP695 through its phosphorylation and dephosphorylation.
In addition, hyperphosphorylation of JIPs contributes to phosphorylation of Thr668 and creates a site for AβPP to bind Pin1, a peptidyl-prolyl cis/trans isomerase. Meanwhile, the promotion of binding between phosphorylated Thr668 of AβPP and Pin1 reduces the combining capacity between Fe65 and phosphorylated Tyr682 of AβPP [130, 131].
Tyr687 phosphorylation site
Tyr687 phosphorylation of AβPP695 also occurs in the AD brain. Tyr687 is the last amino acid residue located at the Y682ENPTY687 motif and is also the primary phosphorylation site for tyrosine kinase. The phosphorylation of AβPP at Tyr687 plays a vital role on transport and cleavage, as well as half-time of AβPP. On one hand, Tyr687 phosphorylation incorporates AβPP into endocytic vesicular structures and regulates AβPP transport from endosomes to the TGN. On the other hand, the endocytic AβPP might undergo amyloidogenic pathway to overgenerate Aβ product [132]. Mutation on Tyr687 will affect phosphorylation of AβPP695 and then influence its cleavage process. Takahashi et al. [133] used a cell model overexpressing AβPP (Tyr687Ala) mutant (this AβPP695 mutant cannot be phosphorylated at 687th residue) to investigate the function of Tyr687 on AβPP cleavage. They found that the proteolytic cleavage of AβPP695 by ADAM10 and γ-secretase was decreased when AβPP695 was mutated (Tyr687Ala). In order to mimic phosphorylation of Tyr687, researchers replaced tyrosine with glutamate at 687 site of amino acid (Tyr687Glu mutant). The result indicates that the Tyr687Glu mutant can increase AβPP’s half-time and reduce Aβ production [134]. In addition, a study has revealed that Fe65 could bind to AICD via its PTB domain [126]. The PTB domain of Fe65 is composed of PTB1 and PTB2 domains. The PTB1 interacts with transcription factor CP2/LSF/LBP1, and the PTB2 domain binds to AICD and histone acetyltransferase to activate transcription [135]. Fe65 associates with AICD through 684NPTY687 motif and Tip60, a histone acetyltransferase, to form “AICD-Fe65-Tip60 (AFT-complex)”. This complex can be shown to concentrate in spherical nuclear spots, which might be the region of DNA translation. However, RAN (ras-related nuclear protein)-binding protein 9 (RanBP9) also can bind AICD and Tip60 to form a novel ternary complex, which prevents nuclear the AICD-induced transcriptional signaling by inhibiting AFT complex formation [136]. So, when AβPP was phosphorylated at Tyr687, AβPP might be cleaved to generate phosphorylated AICD, which binds to Fe65 or RanBP9 and then is imported into nuclear. Intriguingly, the shorter motif N684PTY687 is also essential for coated pit-mediated internalization of the low-density lipoprotein receptor [137]. Although phosphorylation of Thr668 and Tyr687 are related to AICD as well as Fe65, the interaction of AICD with Fe65 seems to be regulated by Thr668 phosphorylation rather than Tyr687 phosphorylation.
To sum up, AβPP phosphorylation sites and their effects on AβPP trafficking and cleavage are shown in the Table 1.
EFFECTS OF GLYCOSYLATION ON AβPP TRAFFICKING AND CLEAVAGE
Glycosylation modification of proteins is one of PTMs and is also a common covalent modification of protein. According to the difference of chemical bonds formed between the amino acid residue and the glycosyl unit, glycosylation modification mainly includes N-linked glycosylation (N-glycosylation) and O-linked glycosylation (O-glycosylation). Un-like N-glycosylation, which occurs in the rough ER and Golgi apparatus, O-glycosylation mainly takes place in the Golgi apparatus. N-linked glycans connect with the nitrogen atom of an asparagine residue whereas O-linked glycans link to the hydroxyl group of serine and threonine of a protein. Compared with O-glycans, N-glycans occur in 90%of glycoproteins. In addition, the glycosylation modification that the group of chondroitin sulfate attached to AβPP is named appican. The core proteins of appican are the spliced isoforms of AβPP, such as AβPP733 and AβPP752. These alternative proteins are translated from AβPP mRNA lacking the exon 15 [138, 139]. These AβPP isomers were found to be ubiquitously expressed except in neurons. There is a serine site that is located in consensus sequence EXSG of AβPP, and this serine residue attaches the xylose sugar of chondroitin sulfate glycosaminoglycan chain via covalent bonds. Here, we gave a brief overview of these three kinds of glycosylation and the potential effects on AβPP trafficking and cleavage.
N-glycosylation of AβPP
The process of protein N-glycosylation starts in the ER and is finished in Golgi apparatus. In the ER, the immature glycoprotein is first formed with the addition of Glc3Man9GlcNAc2 after reaction with Glc3Man9GlcNAc2-pyrophosphate (PP)-dolichol, which is a lipid-linked oligosaccharide and biosynthesized by glycosyltransferases of asparagine-linked glycosylation [140]. The chemical bond is formed between the first N-acetyl-β-D-glucosamine (GlcNAc) in the oligosaccharide and asparagine residue within the sequence of Asn-X-Thr/Ser, where X is referred to any amino acid other than proline. In Golgi apparatus, the oligosaccharide is further modified with the removal of most mannoses but the addition of other carbohydrates to form diverse oligosaccharide chains. After modification of N-glycosylation in ER and Golgi apparatus, a mature glycoprotein would be delivered to cytomembrane [140, 141].
The key enzyme in N-glycosylation is oligosaccharyl transferase complex (OST), which is a component of the translocon in the ER membrane [142]. OST contains seven subunits, including ribophorin I (OST1p), ribophorin II (Swp1p), OST48 (Wbp1p), OST4 (OST4), Stt3-A/Stt3-B (Stt3p), N33/Tusc3 and IAP (OST3p and OST6p), defender against apoptotic cell death 1 (also called DAD1 (OST2p)), and defender against apoptotic cell death 1 (also called DAD1 (OST2p)). Then, the sequential glucose residues of the sugar chain are removed by α-glucosidases I and II in ER. The α-glucosidase I further modifies the immature glycoproteins to remove the terminal mannose (Man) from Man9GlcNAc2 to produce Man8GlcNAc2. Therefore, a number of glycoproteins with an eight- or nine-Man glycan enter the cis-Golgi. Subsequently, under the action of α1,2-mannosidases IA and IB, Man5GlcNAc2 glycan is formed. All of N-linked oligosaccharide chains share a common core structure called trimannosyl core, which is composed of Manα1,3-(Manα1,6)-Manα1,4-GlcNAcβ1,4-GlcNAc-Asn formed by two GlcNAc units and three mannoses.
N-glycosylation regulates biological function through providing recognition signal on the surfaces of cells and plays an important role in folding and stability of protein, subcellular trafficking, and extracellular secretion. Recent studies have implicated that regulator of calcineurin 1 (RCAN 1) is involved in neurodegeneration. Wang et al. [143] showed that RCAN 1 protein was markedly increased in AD brains and its overexpression leaded to neuronal apoptosis. They further found that RCAN 1 can stabilize OST by interacting with OST1p and the overexpression of RCAN 1 enhances protein’s N-glycosylation in ER, resulting in the elevation of the activity of BACE1 and γ-secretase, and marked increase of Aβ production [144].
The N-glycosylation of AβPP was reported by Pahlsson et al. for the first time [145]. In order to further investigate the mechanism of intracellular sorting and processing of the AβPP, Yazaki et al. [146] adopted site-directed mutagenesis to delete two potential N-linked glycosylation sites of AβPP. They substituted the critical asparagine residues Asn467 and Asn496 at AβPP695 with alanine. The results suggested that N-glycosylation may modulate the intracellular sorting of AβPP. Moreover, a recent study suggested that N-glycosylation inhibitor GT863/PE859, a kind of curcumin derivative, can suppress N-glycosylation, leading to inhibit Aβ production via suppressing γ-secretase-mediated cleavage in a substrate-dependent manner rather than inhibiting BACE1 or γ-secretase activity [147]. In addition, APLP-1, a member of the AβPP gene family, whose processing can be modulated by N-glycosylation [148]. These points noticeably prove that AβPP’s N-glycosylation affects the development of AD, but the role of N-glycosylation in regulating AβPP metabolism is needed to extensively study.
O-glycosylation of AβPP
O-glycosylation is formed through covalent bond between serine/threonine or tyrosine residue and the first glycosyl unit in oligosaccharide chain. The first glycosyl unit includes N-Acetyl-D-galactosamine (GalNAc), N-Acetyl-β-D-glucosamine (GlcNAc), and so on. Thus, according to the first glycosyl unit binding serine/threonine or tyrosine, O-glycosylation is classified into GalNAc-type O-linked glycosylation (O-GalNAcylation) and GlcNAc-type O-linked glycosylation (O-GlcNAcylation). The O-GalNAclation is involved in various biochemical processes, such as the process of protein cleavage and cell-cell recognition and communication [149, 150]. The O-GlcNAclation plays a pivotal role in regulating transcriptional activity [151–153], epigenetic regulation [154], protein-protein interaction [155], and response for external stress [156].
O-GalNAcylation is also called mucin-type O-glycosylation being the first sugar added to serine or threonine of a polypeptide acceptor, forming the GalNAc α1-O-serine/threonine structure termed “Tn antigen”. Subsequently, the extra sugars are added, turning out a variety of O-glycan structures. The limiting step of O-GalNAcylation is catalyzed by N-α-acetylgalactosaminyltransferase (GalNAc-T), which belongs to a member of a large family of glycosyltranserases (GTFs). In human, GalNAc-Ts consist of the largest enzyme family among GTFs and have about 20 isozymes. For example, UDP-GalNAc polypeptide, belonging to GalNAc-T family of enzymes, transfers GalNAc from uridine diphosphate N-acetylgalactosamin (UDP) to serine/threonine residue of protein and then produces complex carbohydrate chains. A study has found that Thr291, Thr292, and Thr576 at AβPP695 are mucin-type glycosylation sites by mass spectrometric analyses [157]. Abnormal modification of Mucin-type glycosylation may affect AβPP cleavage within the protein secretory pathway of Golgi complex and trans-Golgi [158]. Recently, Akasaka-Manya et al. [159] have elucidated that GalNAc-T6 glycosylates AβPP at the site proximal to the BACE 1 cleavage site, leading to possibly decrease the level of Aβ1–40 and Aβ1–42.
O-GlcNAcylation is mainly modulated by two kinds of enzymes referred to as O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA). GlcNAc binds hydroxyl of serine/threonine residue of intracellular immature AβPP under the catalysis of OGT. In contrast, the role of OGA is to remove GlcNAc from glycosylated proteins. Therefore, there is a dynamic balance between the gain and loss of O-glycosylation for AβPP. In the brains of patients/mice with AD, the protein level of OGA is decreased more than that of OGT [160, 161]. After inhibiting the activity of OGA by OGA inhibitor O-(2-acetamido-2-deoxy-D-glucopyranosylidene) amino-N-phenylcarbamate (PUGNAc) or siRNA silencing OGA gene, the nonamyloidogenic pathway becomes the major cleavage pathway, which increases the level of sAβPPα and decreases the generation of Aβ [162]. However, the interplay between the cytoplasmic protein O-GalNAcylation and the secretory protein N-/O-glycosylation in AD is needed to describe. A recent report indicated that AD brains showed a change of region-specific glycosylation (both on O-GalNAcylation and N-/O-glycosylation) on both cytoplasmic and membrane glycoproteins. The level of N-/O-glycosylation in serum was significantly decreased in AD patients and the glyco-fingerprint was unique to control cases [163]. Therefore, there might be a novel intracellular cross-talk and a negative interrelationship between the cytoplasmic protein O-GlcNAcylation and the secretory protein N-/O-glycosylation. However, the intracellular cross-talk of AβPP between O-GlcNAcylation and N-/O-glycosylation on AβPP cleavage is needed to further explore.
Chondroitin sulfate proteoglycan of AβPP
Glycosaminoglycans, such as chondroitin sulfate and heparan, have been found in SPs and NFTs. Studies have suggested that heparan sulfate proteoglycan plays a vital role in the pathogenesis of AD [164–166]. Also, many studies have reported the possible functions of appican. For example, appican could inhibit neurite outgrowth in vitro and neurite regeneration following neuronal injury [167, 168]. So, we suggest that the chondroitin sulfate glycosaminoglycan chains could play a vital role in AβPP trafficking and further affect the pathogenesis of AD. Recently, researchers used non-polarized cells to investigate the effect of the glycosaminoglycan on the trafficking of AβPP. They found that the chondroitin sulfate glycosaminoglycans attachment could reduce the rate of endocytosis and accelerate transport of AβPP from the TGN to the plasma membrane. This result suggested that the chondroitin sulfate glycosaminoglycan chain might act as an alternative sorting determinant in cellular AβPP transport. Therefore, this PTM might be dominant over cytoplasmic signals and involved in distinct sorting mechanisms [169]. However, the interplay between chondroitin sulfate chains and phosphorylation on AβPP is less reported and needed to further investigate.
In summary, the glycosylation of AβPP and their effects on AβPP trafficking and cleavage are shown in the Table 2.
AβPP glycosylation and their effects on trafficking and cleavage
INTERACTIONS BETWEEN PHOSPHORYLATION AND GLYCOSYLATION ON AβPP
The phosphorylation and glycosylation of proteins can happen not only on the same amino acid residues but also on the adjacent or distant amino acid residues. Therefore, we could speculate that there may be a direct competition or an indirect regulation between phosphorylation and glycosylation. Proteomic analysis has shown that there might be an extensive interaction between the two PTMs due to spatial competition among the same or adjacent modifications [170, 171]. Wang et al. [172] used NIH3T3 cells to analyze the cross-talk between these two PTMs, and the cells were treated with okadaic acid, PUGNAc, and NAG-thiazoline respectively, or all of the three inhibitors simultaneously. By combining immobilized metal affinity chromatography and TiO2 chromatography to enrich phosphorylated proteins and peptides, using isobaric tags for relative and absolute quantitation (iTRAQ) to quantify protein level, and adopting high throughput mass spectrometry to analyze protein functional groups of GlcNAcylation/phosphorylation, they quantified the phosphorylation changes on 711 phosphopeptides and found that the increasing glycosylation resulted in the alteration of phosphorylation with reduction at 280 amino acid residues and increase at 148 amino acid residues. Therefore, besides inhibiting the phosphorylation of the site itself, the elevated glycosylation of a specific site might interfere with the phosphorylation of other sites. However, we need more solid evidence to prove that the relationship between phosphorylation and glycosylation may play a role on biological functions of proteins.
Phosphorylation and glycosylation can also modify serine/threonine residues on AβPP and affect AβPP’s metabolism. As mentioned above, PUGNAc increases the level of glycosylation of AβPP. When cells were treated with PUGNAc, the Thr576 at AβPP695 is glycosylated, which regulates the trafficking of AβPP695 and the secretion of Aβ1–40 and Aβ1–42 [173]. However, GSK-3β phosphorylates the Thr668 of AβPP695 and affects the generation of Aβ. The interactive effect of phosphorylation and glycosylation at a single or multiple amino acid residues on AβPP695 trafficking and cleavage is remained to further explore.
Interaction of N-glycosylation and phosphorylation on AβPP’s sorting and cleavage
As mentioned above, N-glycosylation seems to be essential for AβPP trafficking and processing, while phosphorylation of AβPP can also have an effect on AβPP cleavage. Therefore, it is worth to further investigate whether impaired N-glycosylation and phosphorylation at the same time would alter AβPP’s trafficking or cleavage. Intriguingly, Tsatsanis and his colleagues [174] focused on modification of N-glycosylation at Asn467/496 and phosphorylation at Ser206 on AβPP. They adopted the mutations on phosphorylation (Ser206Ala) and N-glycosylation (Asn467Lys and Asn496Lys) in AβPP to explore AβPP’s trafficking and degradation. When compared to wild-type AβPP cell line, all these mutant cell lines showed the increased level of total AβPP but the reduced level of cell surface AβPP, total soluble AβPP, and soluble sAβPPβ. These results inferred that the impaired phosphorylation or N-glycosylation of AβPP influences its location on the cell surface and cleavage pathway.
Researchers have proven that N-glycosylation and phosphorylation are essential to AβPP’s location and cleavage. It is reasonable to infer that de-N-glyco-sylation may affect phosphorylation of AβPP. Therefore, in this section, we introduced two enzymes that catalyze deglycosylation of AβPP, termed N-glycanase and endo-β-N-acetyl-D-glucosaminidase, so as to understand the mechanism of deglycosylation. The deglycosylating enzymes of N-glycanase (also called PNGase) and endo-β-N-acetyl-D-glucosaminidase (endo-β-GlcNAc’ase) occur in nature, including various animal cells, plants, and bacteria [175, 176]. PNGase can cleave the bond between the proximal GlcNAc residue and the linkage asparagine residue in glycoproteins, while endo-β-GlcNAc’ase catalyzes the scission of the glycosidic linkage in the N, N’-diacetylchitobiose moi-ety of the core region of N-linked oligosaccharides (Fig. 4) [177]. According to the structural similarity and the enzymatic properties, PNGase can be classified into cytosolic PNGase, acidic PNGase, and PNGase F-like N-glycanase. Because PNGase is a crucial target and valuable tool in analyzing asparagine-linked carbohydrate moieties of glycoproteins, the development of PNGase activity assay technique has advanced considerably [178]. However, the research reports about endo-β-GlcNAc’ase on de-N-glycosylation are rare. Thus, we should further aim at these two deglycosylating enzymes to explore their impacts on interaction of N-glyco-sylation and phosphorylation on cleavage of AβPP.
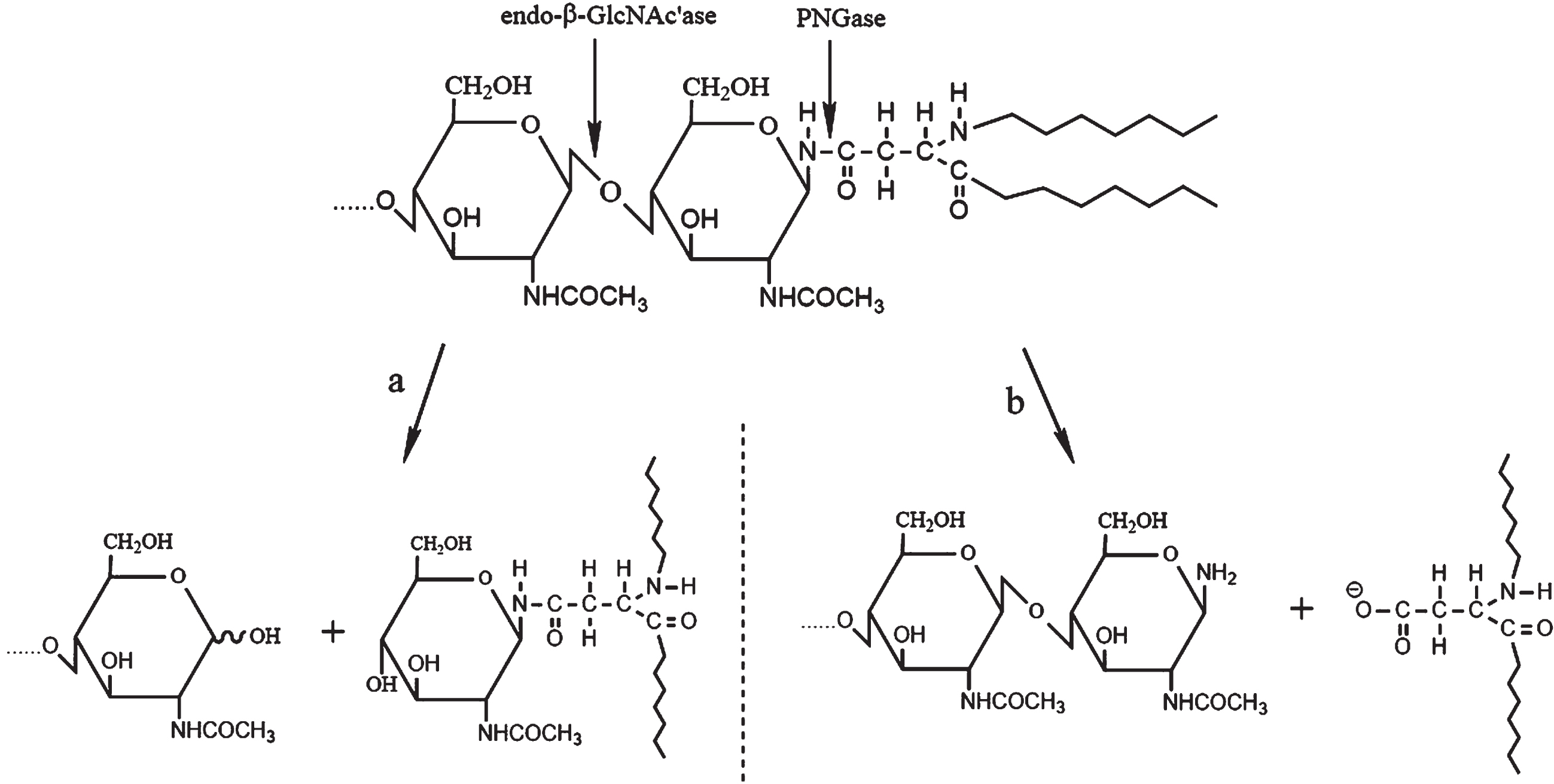
The hydrolytic sites and processes of endo-β-GlcNAc’ase and PNGase on N-linked oligosaccharide chains of glycoprotein. a) Endo-β-GlcNAc’ase. The enzyme catalyzes the scission of the glycosidic linkage in the N, N’-diacetylchitobiose moiety of the core region of N-linked oligosaccharides. b) PNGase. This enzyme can cleave the bond between the proximal GlcNAc residue and the linkage asparagine residue in glycoproteins.
Interaction of O-GlcNAcylation and phosphorylation on AβPP’s trafficking and cleavage
Protein O-GlcNAcylation is in line with protein phosphorylation in that they all occur at serine and/or threonine residues and circulate rapidly. GlcNAc and phosphate can not only competitively occupy a single site and proximal sites but also noncompetitively occupy different sites on the substrates, which means that these two PTMs show a complex interplay. When treated with okadaic acid, the inhibitor of PP1 and PP2A, the global phosphorylation level of the nuclear and the cytoplasmic proteins were elevated but the O-GlcNAcylation level was decreased [179]. Meanwhile, the kinases PKA, PKC, and tyrosine kinase might also be involved in the process of protein O-GlcNAcylation; activators of these kinases reduces the levels of global O-GlcNAcylation of cytoskeletal and cytoskeleton-associated proteins, while inhibition of these kinases results in the increased levels of O-GlcNAcylation in cultured mice cerebellar neurons [180].
The dyshomeostatsis between phosphorylation and O-GlcNAcylation has been increasingly paid attention to. Tau, a pathogenic factor of AD, displays dysregulated relationship between O-GlcNAcyla-tion and phosphorylation; increasing the O-GlcNA-cylation of tau decreases the level of phosphorylated tau [181]. It has been found that an imbalance between phosphorylation and O-GlcNAcylation on tau also exists in transgenic (3xTg-AD) AD mice; the mice at 12 months showed the increase on hyperphosphorylated tau on both Ser396 and Thr205 (the phosphorylation of these two sites are positively correlative with the formation of NFTs) but the reduction on tau O-GlcNAcylation in the hippocampus [182]. This result meant that there might be a competitive modification between phosphorylation and O-GlcNAcylation on tau protein. In addition, O-GlcNAcylation might alter protein’s charge property and structure and thus affect protein’s fate. For example, Frenkel-Pinter et al. [183] adopted the amino acid sequence Ac-301PGGGSVQIVYK311-NH2, which is a corresponding amino acids 301–311 in the longest isoform of full-length tau protein, to examine the effects of the glycan and phos-phate on tau aggregation. The glycopeptide Ac-301PGGGS(GlcNAc)VQIVYK311-NH2 induces the aggregation of PHF6 (Ac-306VQIVYK311-NH2 hexapeptide derived from tau) more rapidly than the phosphopeptide Ac-301PGGGS(p)VQIVYK311-NH2 does. For AβPP, another pathogenic factor associated with “amyloid cascade hypothesis”, it also has a number of sites modulated by O-GlcNAcylation and phosphorylation. Therefore, there might be a complex interplay between O-GlcNAcylation and phosphorylation on AβPP, and the interaction between O-GlcNAcylation and phosphorylation on AβPP’s trafficking and cleavage is needed to further explore.
PROBLEMS AND PROSPECTS
AβPP, a type I membrane glycoprotein, is related to AD development. Here, we introduced five AβPP cleavage pathways: nonamyloidogenic/amyloidoge-nic pathway, η-/δ-secretase pathway, and meprin β-mediated pathway. AβPP can be cleaved by several key enzymes in these cleavage pathways, with the generation of various cleavage productions. So, it is crucial for us to keep a close eye on trafficking and cleavage pathways of AβPP and to ultimately pay attention to Aβ production. Besides, the trafficking and cleavage of AβPP are also closely associated with PTMs, including phosphorylation and glycosylation.
Phosphorylation and glycosylation are two important processes in the post-translational modification of proteins. They may play essential roles in many physiological processes by modifying amino acid residues at the same or different sites. The nascent AβPP needs to undergo post-translational modification processes, such as glycosylation and phosphorylation, to become mature AβPP. AβPP is modified by either one of these two PTMs or by both of them. Moreover, several sites of AβPP may be modified simultaneously. It is needed to further explore and to discuss whether there are synergistic or antagonistic influences between glycosylation and phosphorylation, as well as the detailed mechanism on how they interact with each other. Recently, these two modifications have been attracted extensive attention. However, at present, the roles of glycosylation and phosphorylation in PTMs of AβPP are often studied at the cellular level and is needed to further investigate at the animal level. Furthermore, the effects of phosphorylation sites Tyr653 and Thr686 on the AβPP cleavage pathway also are needed to further explore. With the continuous development and application of mass spectrometry in AβPP research [184, 185], more and more modification sites of glycosylation and phosphorylation on AβPP will be found, and the roles of the two PTMs on AβPP trafficking and cleavage in the pathogenesis of AD will be known in depth.