
Abstract
Background:
Palmitic acid (PA) promotes brain pathologies including Alzheimer’s disease (AD)-related proteins, neuroinflammation, and microglial activation. The activation of neurons and microglia via their Fc gamma receptors (FcγRs) results in producing inflammatory cytokines.
Objective:
To investigate the expression of FcγRs, FcγR signaling proteins, AD-related proteins, proinflammatory cytokines, and cell viability of neurons and microglia in association with PA exposure as well as the effects of FcγR blockade on these parameters in response to PA.
Methods:
200 and 400μM PA-conjugated BSA were applied to SH-SY5Y and HMC3 cells for 24 h. For FcγR blockage experiment, both cells were exposed to FcγR blocker before receiving of 200 and 400μM of PA-conjugated BSA for 24 h.
Results:
PA significantly increased AD-related proteins, including Aβ and BACE1, as well as increasing TNFα, IL-1β, and IL-6 in SH-SY5Y and HMC3 cells. However, the p-Tau/Tau ratio was only increased in SH-SY5Y cells. These results were associated with an increase in FcγRs activation and a decrease in cell viability in both cell types. FcγRs blockage diminished the activation of FcγR in SH-SY5Y and HMC3 cells. Interestingly, blocking FcγRs before PA exposure reduced the increment of AD-related proteins, proinflammatory cytokines caused by PA. FcγRs blocking also inhibits cell death for 23%of SH-SY5Y cells and 64%of HMC3 cells, respectively.
Conclusion:
These findings suggest that PA is a risk factor for AD via the increased AD-related pathologies, inflammation, FcγRs activation, and brain cell death, while FcγR blockage can alleviate these effects.
INTRODUCTION
A high-fat diet (HFD), rich in saturated fatty acids, promotes pathophysiological changes in the brain, and contributes to cognitive decline [1, 2]. Previous study founded that 3 and 30μM of IP-administered saturated fatty acid palmitic acid (PA) caused a decrease in locomotion in a dose dependent manner after injection for 2 h compared to vehicle-treated controls [3]. Moreover, in vitro studies indicated PA causes lipotoxicity in various types of cell lines, including neuronal cell lines (SH-SY5Y) [4], human hepatoma cell lines [5], and astrocytes [6]. In addition, it is known that HFD-induced obesity promotes neuroinflammation and reactive changes in glial cells [7, 8]. These events following HFD consumption led to brain pathologies including increased risk of Alzheimer’s disease (AD) [9, 10]. In recent years, several studies have reported that neuroinflammation, mainly driven by activated microglia, plays an important role in the pathogenesis of AD [11, 12]. AD is characterized by progressive cognitive loss and severe neurodegeneration [13]. The pathological hallmarks of AD are the accumulation of amyloid plaques consisting of amyloid-β (Aβ) peptides and neurofibrillary tangles which are formed from abnormally hyperphosphorylated tau [14, 15]. The abnormal regulation of amyloid precursor protein, including beta-site amyloid precursor protein cleaving enzyme 1 (BACE1) and insulin-degrading enzyme (IDE), caused the accumulation of Aβ, resulting in neuroinflammatory reactions [16, 17]. The mechanisms of neuroinflammation triggered by Aβ and tau protein have been shown to induce the release of proinflammatory cytokines, inflammatory mediators, free radicals and chemokines [18].
Microglia are capable of detecting invading pathogens using surface receptors called Fc gamma receptors (FcγR) [19, 20]. FcγRs are an important bridge between the innate and adaptive immune systems which connect the humoral and cellular immune systems to mediate inflammation. FcγRI, FcγRII, and FcγRIII are expressed in the human brain [21]. FcγRs transmembrane signaling is mediated primarily by two amino acid sequence motifs, ITAMs (Immunoreceptor Tyrosine-based Activation Motifs) and ITIMs (Immunoreceptor Tyrosine-based Inhibition Motifs) that act as the receptors interface with activating and inhibiting signaling pathways, respectively [22]. The FcγR activating signaling pathways are initiated by activated spleen tyrosine kinase (Syk) of the Src family resulting in a phosphorylated ITAMs which is present in the FcγR complex. The recruitment of Syk in FcγR complex initiates the activation of the downstream signaling and the phagocytosis of macrophage [23]. FcγR activation mediated internalization of Aβ and tau by microglia and further induced phagocytosis and degradation of amyloid plaques [24, 25]. Functions of ITAM-bearing receptors are actively counteracted by the action of ITIM-bearing receptors which then activated through SH2- domain-containing inositol polyphosphate 5′ phosphatase (SHIP) 1 results in the inhibition or downmodulation of immune cell effector functions [22]. Previous studies suggested that Aβ promoted the expression of proinflammatory cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6, resulting in elevation of tau hyperphosphorylation, neuronal damage and microglia activation [26–28]. As a consequence, microglia in conjunction with FcγRs are involved in stimulating the clearance of Aβ and tau, releasing cytotoxic mediators, and leading to neuronal cell death [29]. However, the expression of FcγRs, AD-related proteins, including Aβ, AβPP, BACE1, IDE, and tau, proinflammatory cytokines including TNF-α, IL-1β, and IL-6, and cell viability of neurons and microglia in high concentrations of saturated fatty acids, particularly PA, have not been thoroughly investigated. In addition, previous studies have shown that Fc gamma receptors are found on the surface of neurons and interact with soluble Aβ oligomers and mediate neurotoxicity, synaptic dysfunction, and cognitive impairment in AD pathogenesis [30, 31]. However, the role of FcγR blockage on SH-SY5Y and HMC3 cells in response to PA as part of AD-related protein expression, including Aβ, AβPP, BACE1, IDE, and tau, proinflammatory cytokines including TNF-α, IL-1β, and IL-6, and cell viability have not been investigated.
Therefore, this study focused on the effects of various concentrations of PA on SH-SY5Y cells, which is the model for examining neuronal function and differentiation, and human microglia clone 3 (HMC3) as resident innate immune cells of the brain. The aims of the present study were to investigate: 1) the expression of AD-related proteins, including Aβ, AβPP, BACE1, IDE, and Tau protein; 2) presence of inflammatory cytokines, including levels of TNFα, IL-1β, and IL-6; 3) the expression of FcγRs, including FcγRI, FcγRII, and FcγRIII together with Syk or SHIP1 signaling from FcγRs; and 4) cell viability in neurons and microglia after various levels of PA exposure. The hypothesis of the study was that high-doses of PA increased the expression of AD-related proteins, and proinflammatory cytokines in regard to the association with FcγRs and it signaling, resulting in neuronal and microglial cell death. Furthermore, we aimed to investigate: 5) the role of FcγRs blockage pretreatment in neuronal and microglial cells before being exposed to various concentrations of PA. Thus, our hypothesis was that the blockage of FcγRs could reduce the effect of PA on the expression of AD-related proteins, proinflammatory cytokines, and cell death.
MATERIALS AND METHODS
Cell culture models
SH-SY5Y and HMC3, two types of brain cell lines, were used in this study. The SH-SY5Y (ATCC® CRL-2266™) human neuroblastoma cell line SHSY5Y is often used in in vitro models of neuronal function and differentiation [32]. HMC3 (ATCC® CRL-3304™), a human microglia clone 3 cell line, is used as an example of a resident innate immune cell of the brain [33]. They are both well-established cell models for studying neurodegenerative diseases and were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were grown in Eagle’s minimum essential medium (EMEM), sodium bicarbonate, 10%fetal bovine serum (FBS), and 1%penicillin-streptomycin (pen/strep) at 37°C in a 5%CO2 incubator with the media renewed every 2-3 days.
Preparation of palmitic acid (PA)-conjugated bovine serum albumin (BSA)
PA-conjugated BSA was used as the model of lipotoxicity in this study, since a previous study demonstrated that PA-conjugated BSA caused greater levels of lipotoxicity than PA-unconjugated BSA [34]. PA was purchased from Sigma-Aldrich (Dorset, UK). The PA-conjugated BSA complex was prepared using a modified method [35]. Briefly, 10.26 mg of PA was dissolved in 1 mL of 0.01 M Sodium hydroxide (NaOH) in a dry bath of 80°C to reach a final concentration of 40 mM PA stock solution. 1%free fatty acid (FFA)-free BSA was dissolved in EMEM at 37°C, which was filtered through a 0.2μm filter. Then the PA was incubated with FFA-free BSA by adding different volumes of PA stock solution to reach the concentrations of 8 and 16 mM for conditions PA200 and PA400 respectively. PA-conjugated BSA and the vehicle were stored at –20°C and melted in a water bath at 37°C for 30 min for further treatment.
Palmitic acid-conjugated bovine serum albumin treatment
In the study SH-SY5Y and HMC3 cells were treated with two different concentrations of PA-conjugated BSA, specifically 200 (PA200) and 400 (PA400)μM for 24 h. NaOH concentration corresponding to the highest PA concentration in each study was used as the experimental control. Then, we investigated the expression of AD-related proteins including Aβ, AβPP, BACE1, IDE, p-Tau, and Tau, and the FcγR signaling pathway by western blot analysis. Proinflammatory cytokines including TNFα, IL-1β, and IL-6 were determined by qRT-PCR. FcγRs level and cell viability were determined by flow cytometry.
FcγR blockage pretreatment
SH-SY5Y and HMC3 were plated and cultured in EMEM at 37°C in a 5%CO2 incubator overnight. After that, 107 cells were pre-treated with 20μl of FcγR blocking reagent (Cat no. 130-059-901; Miltenyi Biotec B.V. & Co. KG, Bergisch Gladbach, Germany) for 1 h following the commercial protocol. Then, PA-conjugated BSA at the concentrations of 200 and 400μM were added and the cells were incubated for 24 h. NaOH concentration corresponding to the highest PA concentration was used as the experimental control. Then, we investigated the expression of AD-related proteins including Aβ, AβPP, BACE1, IDE, p-Tau, and tau, and the FcγR signaling pathway by western blot analysis. Proinflammatory cytokines including TNFα, IL-1β, and IL-6 were determined by qRT-PCR. Cell viability was determined by MTT assay.
Western blot analysis
All treatment groups in the SH-SY5Y and HMC3 cells were lysed with an extraction buffer and separated by electrophoresis on 10%sodium dodecyl sulfate–polyacrylamide gel and then transferred onto nitrocellulose membranes. They were then probed overnight at 4°C with primary antibodies (1:1000) against AD-related protein expression. These included Aβ (sc28365; Santa Cruz Biotechnology, USA), AβPP, tau, p-Tau (Thr181), ((AβPP, 12843; tau, 4019; p-Tau (Thr181), 12885), Cell Signaling Technology, USA), BACE1, IDE (BACE1, ab2077; IDE, ab133561, Abcam, USA), and FcγR signaling proteins p-Syk and Syk (p-Syk, 2710; Syk, 2712, Cell Signaling Technology, USA). These included Aβ (Santa Cruz Biotechnology, USA), AβPP, tau, p-Tau, (Cell Signaling Technology, USA), BACE1, IDE (Abcam, USA), and FcγR signaling proteins p-Syk and Syk (Cell Signaling Technology, USA). β-actin was used as a loading control (Santa Cruz Biotechnology, USA). This was followed by incubation with horseradish peroxidase-conjugated secondary antibody (1:1000) at room temperature for 1 h. The membranes were visualized using Clarity™ Western ECL Blotting Substrate (Bio-Rad Laboratories Ltd.) and scanned using the ChemiDoc™ Touch Gel Imaging System. The western blotting images were analyzed by Image J (NIH image) analysis software.
RNA extraction and quantitative real-time PCR (qRT-PCR)
All treatment groups had the RNA extracted RNA using the RNeasy Mini Kit (QIAGEN). Then, cDNA was synthesized using the Tetro™ cDNA Synthesis Kit (Bioline, USA) according to the manufacturer’s instructions. qRT-PCR was performed using the SYBR-Green-based method, the primer pairs shown as follows: TNFα, 5′-GCTGCACTTTGGAGTGATCGF -3′ (forward) and 5′-CTTACCTACAACATGGGCTACAG-3′ (reverse); IL-1β, 5′-TTACAGTGGCAATGAGGATGAC-3′ (forward) and 5′-GTCGGAGATTCGTAGCTGGAT-3′(reverse); IL-6, 5′-ATGAACTCCTTCTCCACAAGCGC-3′ (forward) and 5′-GAAGAGCCCTCAGGCTGGACTG-3′ (reverse);
β-actin, 5′-CCAGATCATGTTTGAGACC-3′ (forward) and 5′-ATGTCACGCACGATTTCCC-3′(reverse). The relative changes in gene expression were normalized against the β-Actin and quantified using the 2-ΔΔCT method.
Flow cytometry for Fc gamma receptor expression
The expression of FcγRI, FcγRII, and FcγRIII by SH-SY5Y and HMC3 cells after being exposed to PA-conjugated BSA was determined by flow cytometry. After treatment for 24 h, the cells were harvested and washed with 1x PBS. The collected cells were then resuspended in 10%AB serum in 0.02%sodium azide (NaN3) dissolved PBS and incubated on ice for 30 min to block non-specific binding sites. FcγRs were probed with anti-FcγRI (CD64), anti-FcγRII (CD32), and anti-FcγRIII (CD16) (BD Pharmingen™, USA) for 30 min on ice in the dark. Finally, cells were washed with 1%BSA in 0.02%NaN3 dissolved PBS and fixed with 1%paraformaldehyde. Cells were examined immediately using a BD FACSCelesta™ flow cytometer.
Flow cytometry for cell viability using Annexin V/propidium iodide staining
SH-SY5Y and HMC3 cells treated with PA-conjugated BSA were used to determine cell apoptosis using flow cytometry-based methods. After 24 h of treatment, cells were harvested then washed and probed with Annexin V/Propidium Iodide (PI) Staining (BD Pharmingen™ FITC and PE Annexin V Apoptosis Detection Kits, USA). Samples were measured immediately using a BD FACSCelesta™ flow cytometer results being represented on a two-parameter dot-plot graph. The representative graph shows four quadrants (Q) that are composed of a Q1 quadrant representing necrotic cells (PI positive and Annexin V negative); Q2 quadrant representing late apoptosis (both PI and Annexin V positive); Q3 quadrant representing viable cells (both PI and Annexin V negative); and Q4 quadrant representing early apoptosis (PI negative and Annexin V positive).
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
SH-SY5Y and HMC3 were seeded for 1×104 cells/well in a 96-well plate and incubated at 37°C in a 5%CO2 incubator overnight. After renewal of the culture media and 24 h-treatment in each experiment, the EMEM without FBS was replaced. After that, 15μl of MTT solution in PBS (5 mg/ml) was directly added to the plate then incubated for 4 h. Finally, the culture media was removed, and formazan crystals were dissolved in 100μl of DMSO. The absorbance was measured at a wavelength of 570 nm and was subtracted by the interferences measured at a reference wavelength of 630 nm [36, 37].
Oil Red O staining
All treatment groups of SH-SY5Y and HMC3 cells were harvested and washed with 1x PBS then 1×104 cells were diluted in 100μl of EMEM. The cells were centrifuged at 1000×g, 5 min by cytospin to isolate, concentrate, and deposit a monolayer of cells from a dilute cell suspension onto a circular area on a slide. After the slides dry, rinsed with 70%alcohol. Then stain in Oil Red O solution for 10 min and differentiate with 70%alcohol 2 times followed by running tap water until the water is clear. The slides were counterstained with hematoxylin for 1 min then washed with tap water 3 min. To remove excess stain, rinse with 1%acid then washed with tap water 3 min follow by rinse with lithium carbonate and washed with tap water 3 min. Finally, mount with mounting medium. The results showed as bright red staining of lipids, while the nuclei are counterstained blue with hematoxylin solution.
Statistical analysis
Data has been reported as the mean±standard error of mean and processed using the statistical software GraphPad Prism 7.00 (GraphPad Software). One-way ANOVA followed by Tukey’s post hoc test was performed to investigate AD-related proteins, proinflammatory cytokines, cell viability by MTT assay, FcγRs signaling proteins, and fat accumulation by Oil Red O staining. Two-way ANOVAs followed by Tukey’s post hoc test were performed for cell viability by flow cytometry and FcγRs levels. A p-value <0.05 was considered statistically significant.
RESULTS
The effect of PA-conjugated BSA on the expression of AD-related proteins by SH-SY5Y and HMC3 cells
SH-SY5Y cells exposed to 200 and 400μM of PA-conjugated BSA significantly increased the production of Aβ peptides (p = 0.0336) and BACE1 (Fig. 1A-C) but decreased the expression of IDE (Fig. 1D). These cells treated in a similar manner had a significantly increased p-Tau/Tau ratio (Fig. 1E). Similar to the response in HMC3 cells, 200 and 400μM of PA-conjugated BSA exposure caused a significant increase in levels of Aβ peptides and BACE1 enzyme and decreased IDE as shown in Fig. 1F-I. Contrary to what was observed in SH-SY5Y cells, neither dose of PA-conjugated BSA had any effect on the p-Tau/tau ratio of HMC3 cells (Fig. 1J).
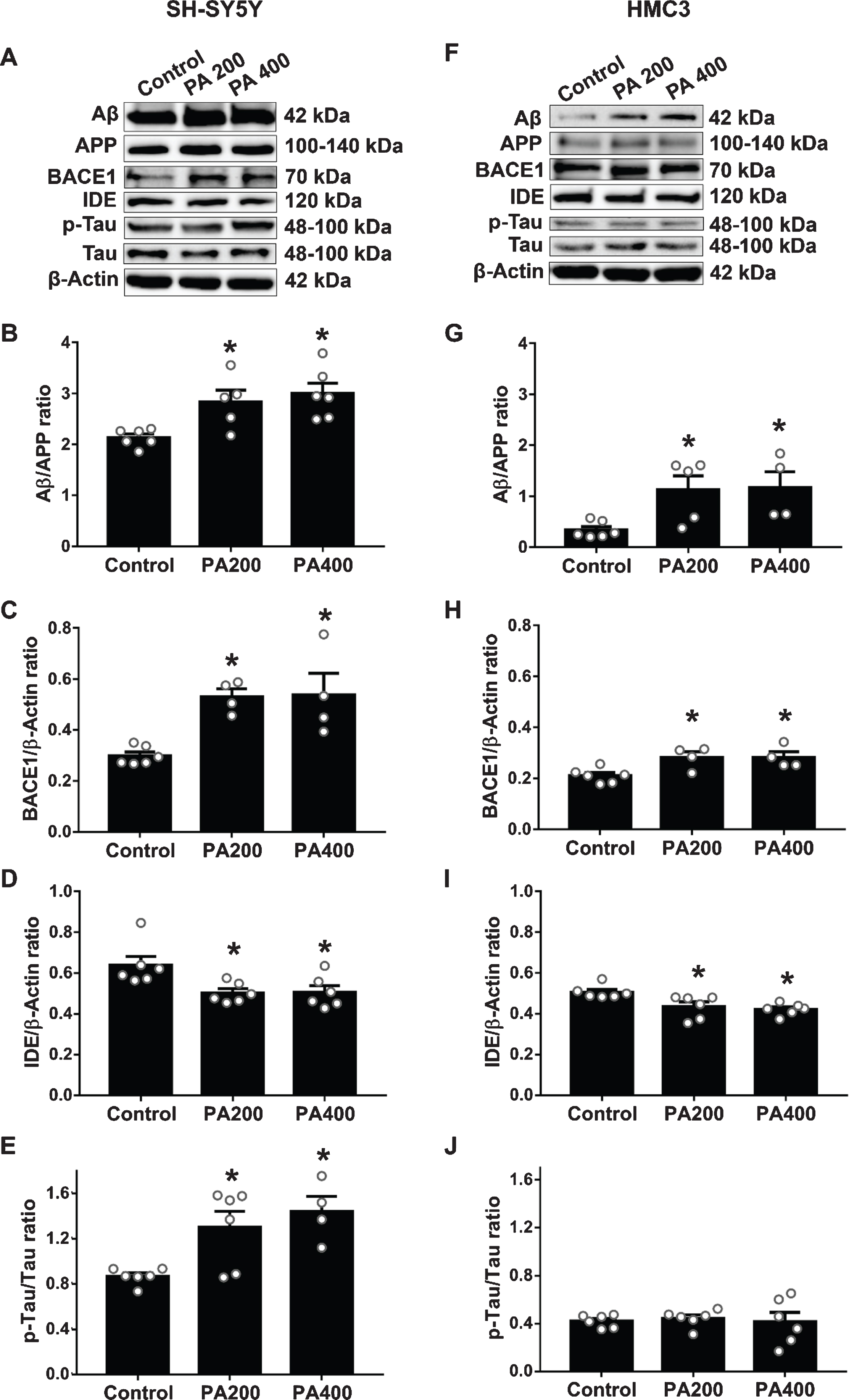
The effect of palmitic acid (PA)-conjugated bovine serum albumin (BSA) on the expression of AD-related proteins and enzymes in SH-SY5Y and HMC3 cells (N = 4–6/group). Western blot analysis results from SH-SY5Y cells: (A) bands representative of named proteins, followed by specifics: (B) amyloid-β (Aβ)/amyloid-β protein precursor (AβPP) ratio, (C) beta-site amyloid precursor protein cleaving enzyme 1 (BACE1), (D) insulin-degrading enzyme (IDE), and (E) phosphorylated Tau (p-Tau)/tau ratio. For HMC3 cells show (F) bands representative of named proteins, followed by specifics: (G) Aβ/AβPP ratio, (H) BACE1, (I) IDE and (J) p-Tau/Tau ratio. All data are means±SEM. *p < 0.05 versus control. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of PA-conjugated BSA on mRNA expression of proinflammatory cytokines by SH-SY5Y and HMC3 cells
SH-SY5Y cells treated with 200μM of PA-conjugated BSA did not show any changes in the expression of proinflammatory cytokines including TNFα (Fig. 2A), IL-1β (Fig. 2B), and IL-6 (Fig. 2C). However, after SH-SY5Y cells were exposed to 400μM of PA-conjugated BSA, an increase in TNFα, IL-1β, and IL-6 levels (Fig. 2A, B, and C, respectively), when compared with control group and 200μM of PA-conjugated BSA were observed. Similarly, 200μM of PA-conjugated BSA did not affect the production of proinflammatory cytokines in HMC3 cells. However, HMC3 cells treated with 400μM of PA-conjugated BSA showed a significant increase the levels of TNFα, IL-1β, and IL-6 (Fig. 2D, E, and F, respectively), when compared with control group.
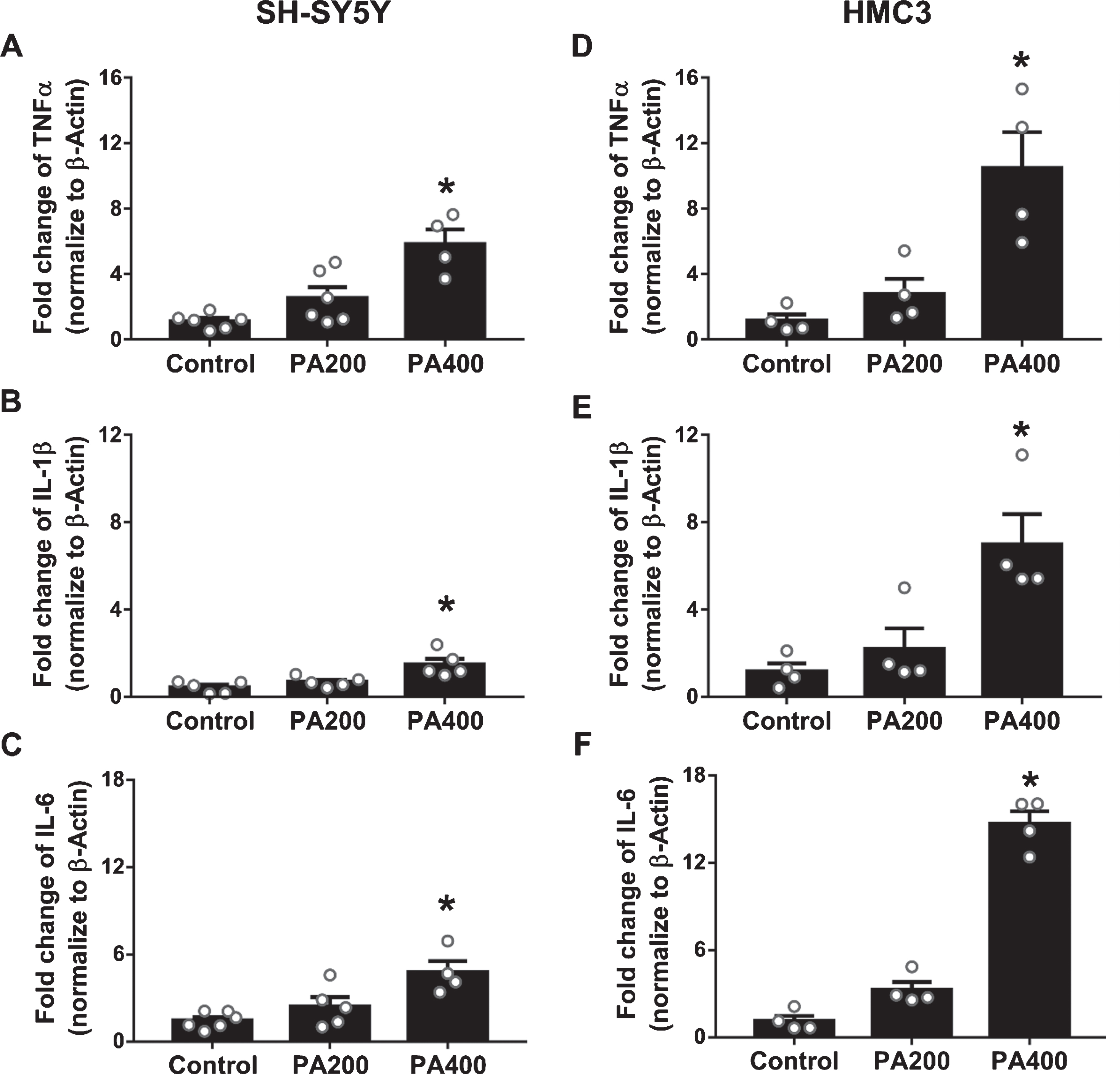
The effects of palmitic acid (PA)-conjugated bovine serum albumin (BSA) on the production of inflammatory cytokines in SH-SY5Y and HMC3 cells (N = 4–6/group). (A) Tumor necrosis factor-α (TNFα), (B) interleukin (IL)-1β, and (C) IL-6 from SH-SY5Y cells, and (D) TNFα, (E) IL-1β, and (F) IL-6 from HMC3 cells. All data are means±SEM. *p < 0.05 versus control and PA200. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of PA-conjugated BSA on the expression of FcγRs and their signaling proteins by SH-SY5Y and HMC3 cells
There were no changes in the expression by SH-SY5Y cells treated with both 200 and 400μM of PA-conjugated BSA on FcγRs including FcγRI, FcγRII, and FcγRIII in comparison to the control group (Fig. 3A). However, HMC3 cells treated with 400μM PA-conjugated BSA showed significantly increased levels of FcγRII and FcγRIII (Fig. 3B). FcγRI levels in HMC3 cells exposed to 200 and 400μM PA-conjugated BSA did not differ among the groups. The western blot analysis showed that SH-SY5Y cells treated with 200 and 400μM of PA-conjugated BSA had significantly increased p-Syk/Syk ratios (Fig. 3C) which indicated that PA-conjugated BSA treatment can activate the FcγR signaling pathway. HMC3 cells treated with 200 and 400μM of PA-conjugated BSA significantly increased p-Syk/Syk ratios, similar to SH-SY5Y cells (Fig. 3D).

The effect of palmitic acid (PA)-conjugated bovine serum albumin (BSA) on the Fc gamma receptors (FcγRs) expression of SH-SY5Yand HMC3 cells (N = 4/group). The dimensional dot-plots of: (A) SH-SY5Y and (B) HMC3 cells represent FcγR positive cells. Bar graphs show the average percentage of positive cells calculated in comparison with the control group (N = 6/group). Furthermore, western blot analysis show (C) phosphorylated Syk (p-Syk)/Syk ratio of SH-SY5Y cells and (D) p-Syk/Syk ratio of HMC3 cells. All data are means±SEM. *p < 0.05 versus control. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of PA-conjugated BSA on the viability of SH-SY5Y and HMC3 cells
The relative viability of SH-SY5Y and HMC3 cells treated with various concentrations of PA-conjugated BSA is shown in the representative dot-plot and bar graph of flow cytometry data (Fig. 4A, B). SH-SY5Y cells treated with 200 and 400μM of PA-conjugated BSA showed significantly decreased cell viability in a dose-dependent manner (Fig. 4A). However, in HMC3 cells, the cell viability was only significantly decreased when treated with 400μM of PA-conjugated BSA (Fig. 4B).
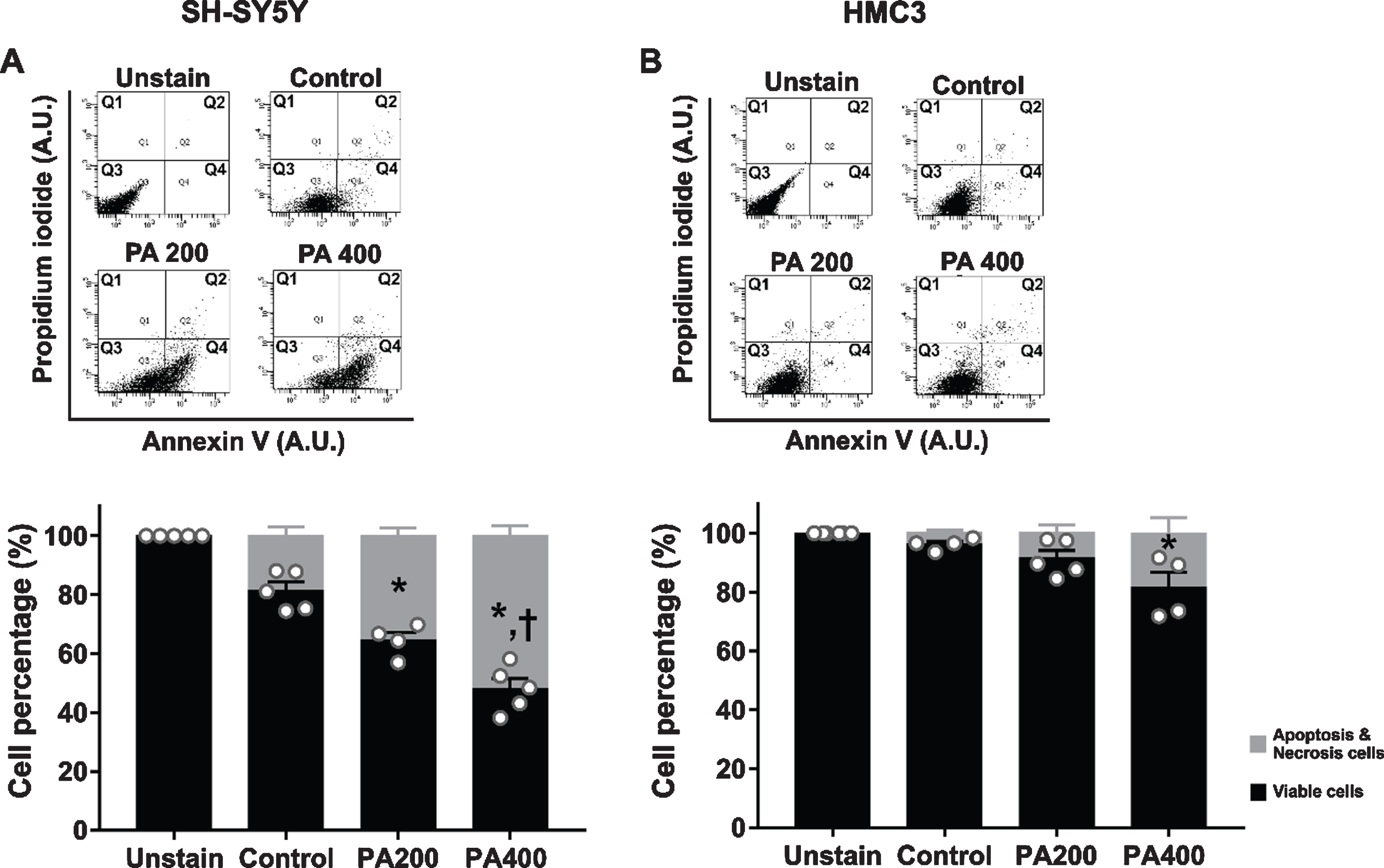
The effect of palmitic acid (PA) conjugated bovine serum albumin (BSA) on cell viability of SH-SY5Y and HMC3 cells. N = 4–6/group. Representative flow cytometry two-parameter dot-plots and average cell percentage of (A) SH-SY5Y and (B) HMC3 cells. Q1 quadrant representing necrotic cells (PI positive and Annexin V negative); Q2 quadrant representing late apoptosis (both PI and Annexin V positive); Q3 quadrant representing viable cells (both PI and Annexin V negative); Q4 quadrant representing early apoptosis (PI negative and Annexin V positive). All data are means±SEM. *p < 0.05 versus control and †p < 0.05 versus PA200. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of FcγR blockage pretreatment on the expression of FcγR signaling in SH-SY5Y and HMC3 cells in response to PA-conjugated BSA
The results showed that 200 and 400μM of PA-conjugated BSA had a significantly increased p-Syk/Syk ratio in both SH-SY5Y (Fig. 5A, B) and HMC3 cells (Fig. 5D, E). In addition, both SH-SY5Y and HMC3 cells pre-incubated with FcγR blocking reagent prior to exposure to the two experimental concentrations of PA-conjugated BSA resulted in significantly decreased p-Syk/Syk ratios (Fig. 5A, B, D, E). The percentage reduction of Syk following FcγR blocking of SH-SY5Y cells was 42%and 34%in 200 and 400μM PA-conjugated BSA, respectively. For HMC3 cells, the percentage reduction of Syk following FcγR blocking was 41%and 42%in 200 and 400μM PA-conjugated BSA, respectively. However, the levels of Syk in FcγR blocking group without PA-conjugated BSA were not significantly different from the control group both in SH-SY5Y and HMC3 cells. Furthermore, the results showed that 200 and 400μM of PA-conjugated BSA did not affect to the levels of p-SHIP1/SHIP1 ratio of both SH-SY5Y (Fig. 5A, C) and HMC3 cells (Fig. 5D, F). These findings suggested that this FcγR blocking reagent can inhibit the activation of FcγR from PA as shown by the reduction in phosphorylation of the Syk signaling protein.
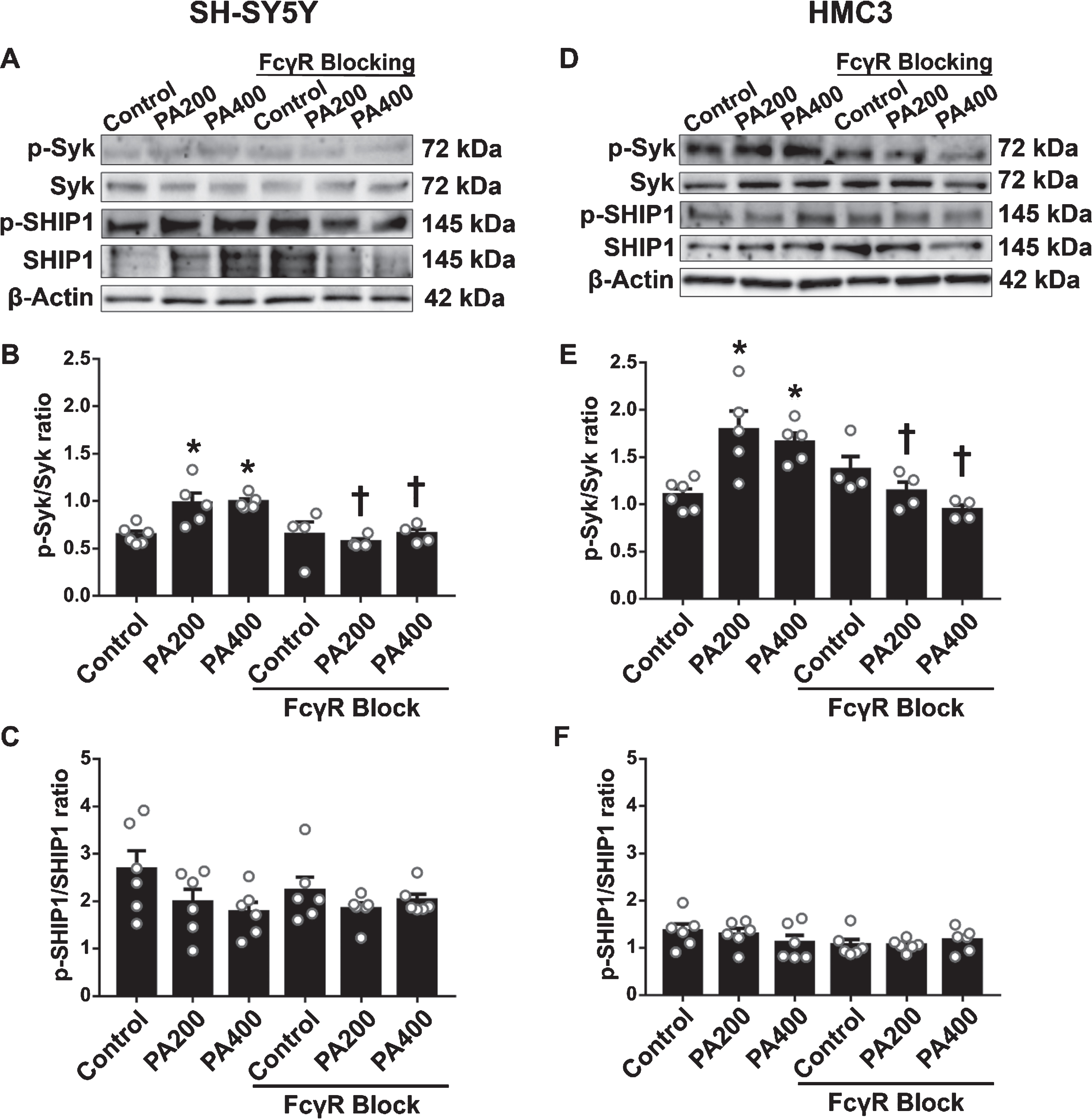
The effect of Fc gamma receptor (FcγR) blockage pretreatment on FcγR signaling proteins in SH-SY5Y and HMC3 cells following exposure to palmitic acid (PA)-conjugated bovine serum albumin (BSA) (N = 4–6/group). (A) Band representative of proteins in SH-SY5Y cells as follows (B) phosphorylated Syk (p-Syk)/Syk ratio and (C) p-SHIP1/SHIP1; (D) Band representative of proteins in HMC3 cells: (E) p-Syk/Syk ratio and (F) p-SHIP1/SHIP1. All data are means±SEM. *p < 0.05 versus control without FcγR blockage; and †p < 0.05 versus the same concentration of PA without FcγR blockage. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of FcγR blockage pretreatment on the expression of AD-related proteins by SH-SY5Y and HMC3 cells in response to the two doses of PA-conjugated BSA
After FcγR blockage pretreatment, SH-SY5Y cells showed significant decreases in the production of Aβ peptides and BACE1 when exposed to 200 and 400μM of PA-conjugated BSA (Fig. 6A-C and F-H). However, the expression of Aβ peptides and BACE1 in HMC3 cells tend to decrease in both 200 and 400μM of PA-conjugated BSA group. Both types of cells also showed increased expression of IDE (Fig. 6D, I) when compared to the cells without FcγR blockage at the same concentration of PA-conjugated BSA. In addition, only SH-SY5Y cells pre-incubated with FcγR blocking reagent had a significantly decreased p-Tau/tau ratio when exposed to 200 and 400μM PA-conjugated BSA, in comparison to outcomes from similar cells in response to the same concentration of PA-conjugated BSA without FcγR blockage (Fig. 6E). On the other hand, there was no change in the p-Tau/tau ratio in HMC3 cells pretreated with FcγR blocking reagent and exposed to 200 and 400μM PA-conjugated BSA (Fig. 6J).
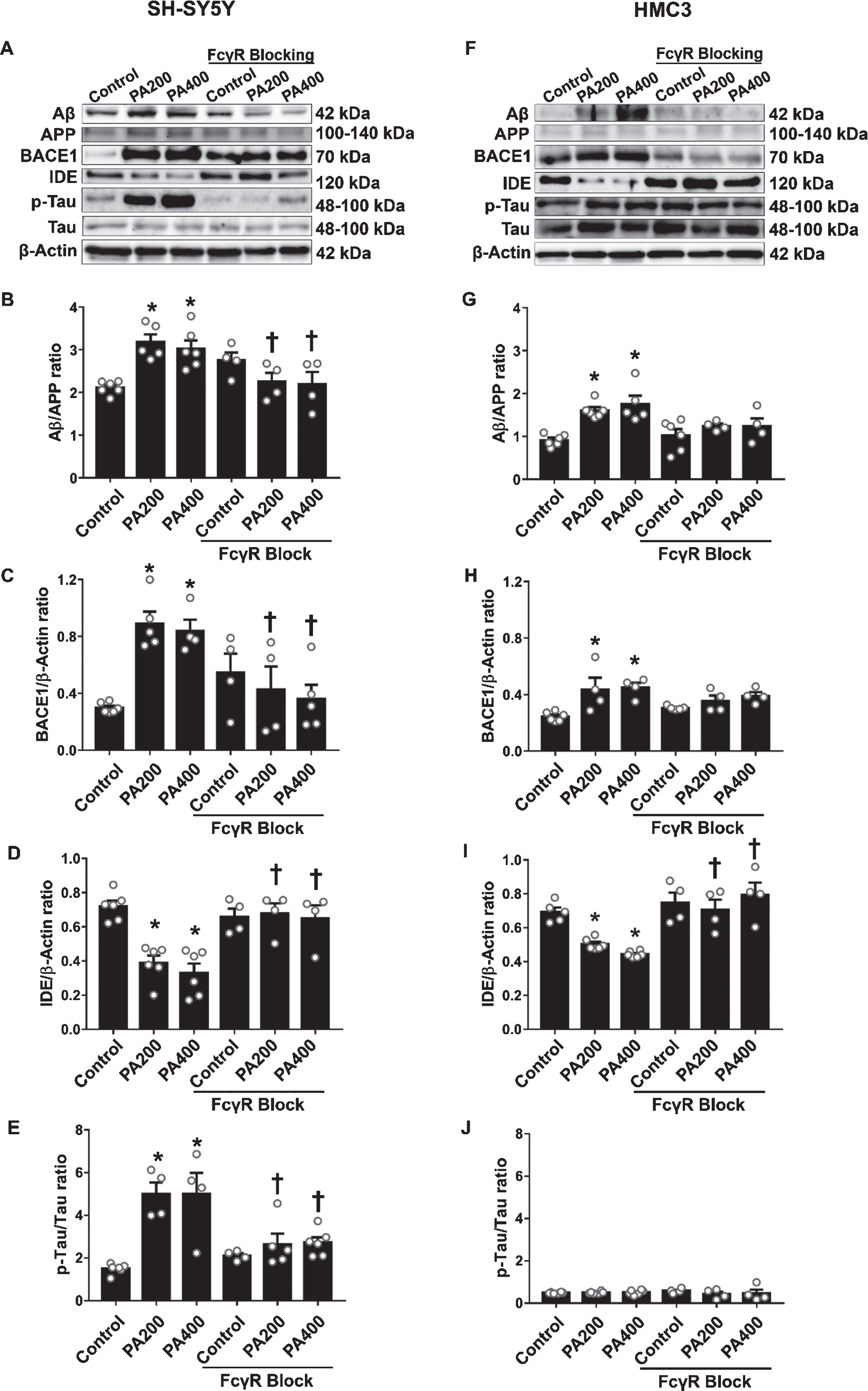
The effect of Fc gamma receptor (FcγR) blockage pretreatment on the expression of AD-related proteins and enzymes by SH-SY5Y and HMC3 cells in response to palmitic acid (PA)-conjugated bovine serum albumin (BSA) exposure (N = 4–6/group). (A) Band representative of proteins in SH-SY5Y cells as follows (B) amyloid-β (Aβ)/amyloid-β protein precursor (AβPP) ratio, (C) beta-site amyloid precursor protein cleaving enzyme 1 (BACE1), (D) insulin-degrading enzyme (IDE), and (E) phosphorylated Tau (p-Tau)/Tau ratio; (F) Band representative of proteins in HMC3 cells: (G) Aβ/AβPP ratio, (H) BACE1, (I) IDE, and (J) p-Tau/Tau ratio. All data are means±SEM. *p < 0.05 versus control without FcγR blockage; and †p < 0.05 versus the same concentration of PA without FcγR blockage. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of FcγR blockage pretreatment on expression of proinflammatory cytokines by mRNA in SH-SY5Y and HMC3 cells in response to the two doses of PA-conjugated BSA
In the FcγR blockage group, both SH-SY5Y and HMC3 cells exposed to 400μM of PA-conjugated BSA showed significant decreases in TNFα and IL-6 levels when compared with the same concentration of PA-conjugated BSA without FcγR blockage group (Fig. 7A, C and 7D, F). However, pretreatment with the FcγR blocking reagent did not reduce the levels of IL-1β in both SH-SY5Y and HMC3 cells when exposed to 400μM of PA-conjugated BSA, in comparison to the reaction by the same cells without FcγR blockage to the same concentration of PA-conjugated BSA (Fig. 7B, E).
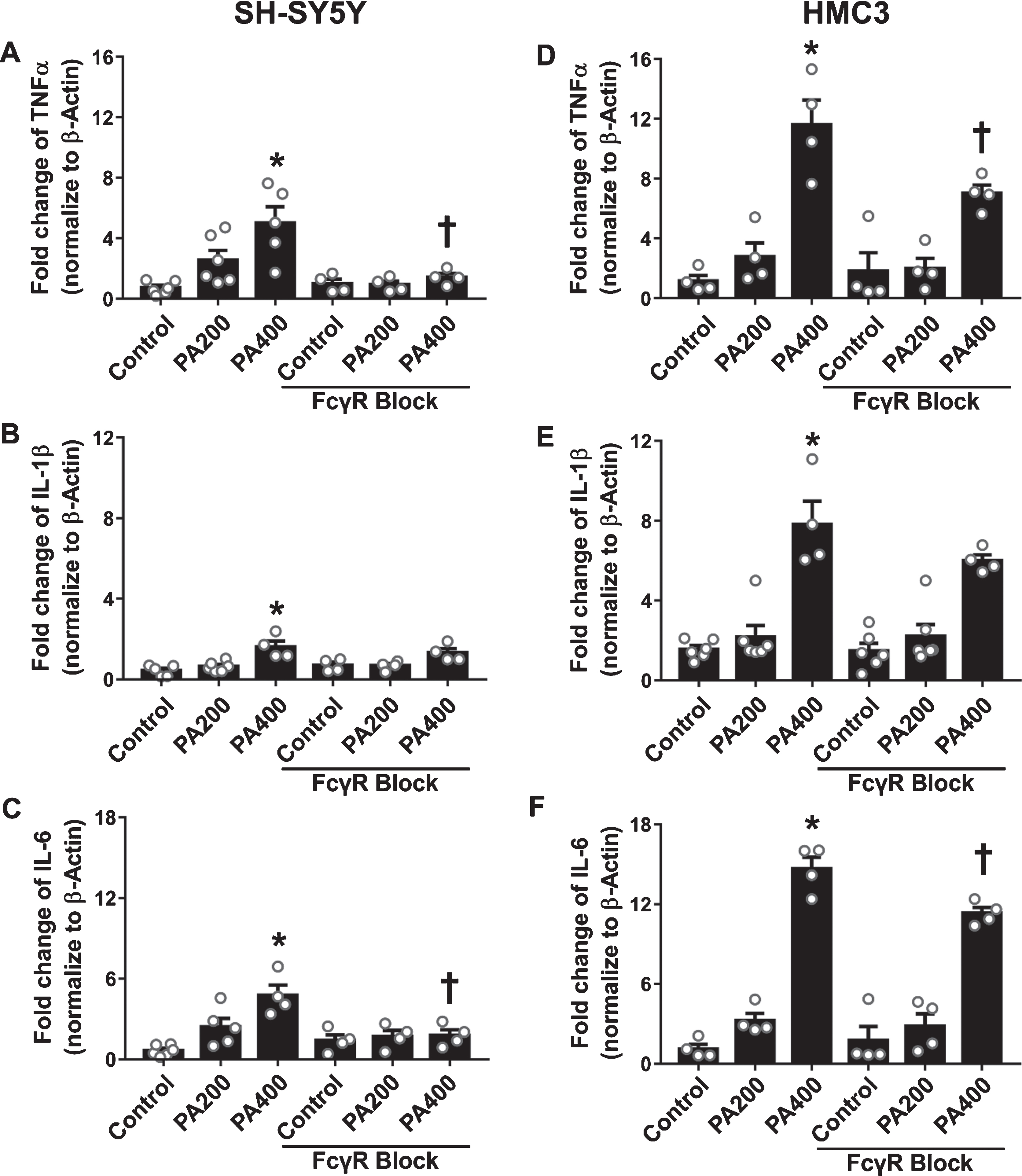
The effect of Fc gamma receptor (FcγR) blockage pretreatment on the production of inflammatory cytokines in SH-SY5Y and HMC3 cells in response to palmitic acid (PA)-conjugated bovine serum albumin (BSA) exposure (N = 4–6/group). (A) Tumor necrosis factor-α (TNFα), (B) interleukin (IL)-1β, and (C) IL-6 from SH-SY5Y cells, and (D) TNFα, (E) IL-1β, and (F) IL-6 from HMC3 cells. All data are means±SEM. *p < 0.05 versus control without FcγR blockage; and †p<0.05 versus the same concentration of PA without FcγR blockage. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of FcγR blockage pretreatment on the cell viability of SH-SY5Y and HMC3 cells after exposure to 200 and 400μM of PA-conjugated BSA
SH-SY5Y pre-incubated with FcγR blocking reagent showed significantly increased cell viability 21%and 24%when exposed to 200 and 400μM of PA-conjugated BSA respectively, comparing to the group without FcγR blockage in the same concentration of PA-conjugated BSA (Fig. 8A). The cell viability of HMC3 cells pre-incubated with FcγR blocking reagent was significantly increased for 64%only when exposed to 400μM of PA-conjugated BSA but did not change in 200μM of PA-conjugated BSA exposure group in compared to the same concentration of PA-conjugated BSA without FcγR blockage group (Fig. 8B).

The effect of Fc gamma receptor (FcγR) blockage pretreatment on viability in SH-SY5Y and HMC3 cells in response to palmitic acid (PA)-conjugated bovine serum albumin (BSA) exposure (N = 6–8/group). Bar graphs show the average percentage of cell viability of (A) SH-SY5Y and (B) HMC3 cells. All data are means±SEM. *p < 0.05 versus control without FcγR blockage; and †p < 0.05 versus the same concentration of PA without FcγR blockage. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
The effect of FcγR blockage pretreatment on the lipid accumulation of SH-SY5Y and HMC3 cells after exposure to 200 and 400μM of PA-conjugated BSA
Lipid accumulation were detected by Oil Red O staining and the results founded that high concentration of PA caused lipid accumulation in both SH-SY5Y and HMC3 cells (Fig. 9A, B). Lipid accumulation of SH-SY5Y cells was increased 45%and 53%by receiving 200 and 400μM of PA-conjugated BSA, respectively. Likewise, for HMC3 cells showed increased 38%and 45%lipid accumulation by receiving 200 and 400μM of PA-conjugated BSA, respectively. However, lipid accumulation between PA200 and PA400 group did not different among group. In addition, FcγR blockage did not affect the accumulation of lipid in both SH-SY5Y and HMC3 cells.
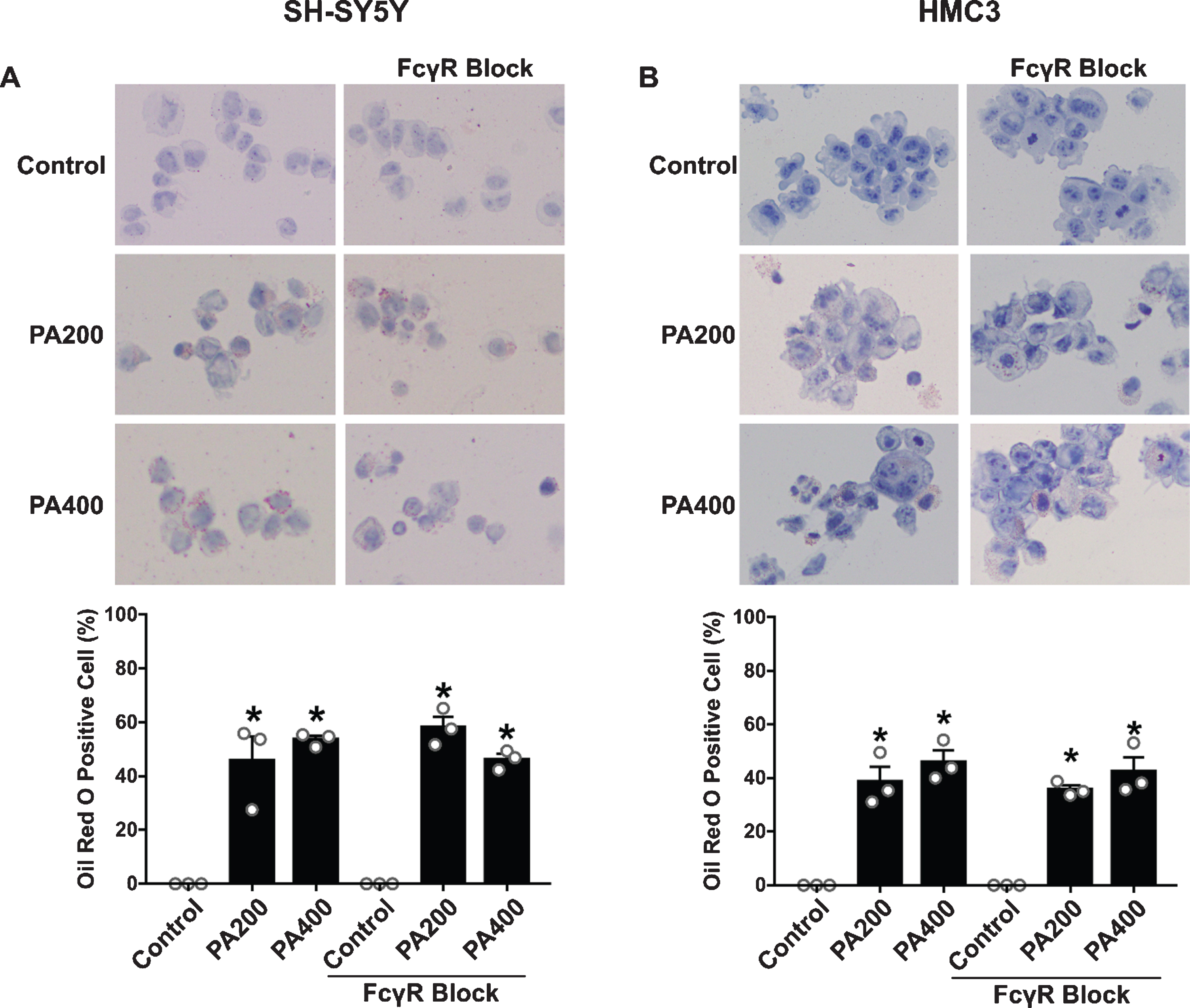
The effect of palmitic acid (PA)-conjugated bovine serum albumin (BSA) on lipid accumulation in SH-SY5Y and HMC3 cells (N = 3/group). Oil Red O staining of (A) SH-SY5Y and (B) HMC3 cells. Bright red staining represents lipids and blue represents nuclei at 40x magnification. All data are means±SEM. *p < 0.05 versus control without FcγR blockage and control of FcγR blockage group. PA200, palmitic acid 200μM; PA400, palmitic acid 400μM.
DISCUSSION
The major findings of this study are as PA-conjugated BSA caused the upregulation of Aβ peptides, and BACE1, and decreased IDE levels as well as increased proinflammatory cytokines including TNF-α, IL-1β, and IL-6 in neuronal and microglial cells. Only neuronal cells were seriously affected by PA-conjugated BSA as regards to the increment of p-Tau protein. PA-conjugated BSA increased the expression of FcγRII and FcγRIII by microglial cells while there was no alteration in the expression of these by neuronal cells. However, the activation of the FcγR signaling pathway as shown by increased phosphorylation of Syk was found in both neuronal and microglial cells in exposed to PA-conjugated BSA. As a result, PA-conjugated BSA caused neuronal and microglial cell death in a dose-dependent manner. Consequently, both neuronal and microglial cells which received FcγR blocking reagent was found to decrease the activation of the FcγR signaling protein, p-Syk. FcγR blockage pretreatment prior to exposure to PA-conjugated BSA also reduced the upregulation of Aβ peptides, BACE and proinflammatory cytokines of neuronal and microglia cells but only reduced p-Tau proteins in neuronal cells. Eventually, the cell viability of both cells treated with FcγR blockage was increased even in PA conditions.
Epidemiological studies have suggested that a diet with high-saturated fats is one of major risk factors for the development of AD in middle-aged people [38]. The previous findings showed that the level of saturated fatty acids, defined as PA, was found as the highest upregulation among other fatty acid in the hypothalamus of HFD-fed mice (60%fat) for either 4 or 8 weeks, when compared with ND-fed mice (13%fat) [39]. In addition, PA has been shown to be one of the major risk factors for the development of AD [4]. It has been shown that PA induces inflammatory signaling, abnormal proliferation, differentiation, and increases apoptosis of brain cells [35, 40–42]. Previous in vitro studies indicated PA causes lipotoxicity in various types of cell lines, including neuronal cell lines (SH-SY5Y) [4], human hepatoma cell lines [5], and astrocytes [6]. Furthermore, a previous study reported that PA at 200 and 400μM, which are the concentrations that we used in this study, represented a high physiological level of circulating PA in obesity [43]. To compare the levels of PA (200 and 400μM in this study) to brain PA levels, Melo et al., 2020 reported that the concentration of PA levels was significantly increased in the cerebrospinal fluid (CSF) of overweight and obese humans with amnestic mild cognitive impairment [44]. Their results showed that the levels of CSF PA in overweight and obese subjects was about 150–410μM and 380–590μM, respectively. In addition, their results showed that elevated levels of PA in CSF have been correlated with decreased cognitive performance in overweight individuals with diabetes, dyslipidemia, and/or hypertension, but not in overweight subjects without these conditions. Accordingly, these findings suggest that the concentration of PA used in our study are able to affect neuronal and microglial cells, as similar with that clinical study. Accordingly, these concentrations of PA were used in this study to investigate the effect of PA on the expression of AD-related proteins, proinflammatory cytokines, and FcγRs with their signaling proteins in SH-SY5Y and HMC3 cells. The results illustrated that PA was found to be a neurotoxic substance to neuronal and microglial cells as shown by the upregulated levels of proteins and enzymes associated with the development of AD pathology. Rising levels of Aβ peptides, BACE1, and tau proteins as well as decreasing levels of IDE were observed when SH-SY5Y cells were exposed to 200 and 400μM of PA-conjugated BSA. These results were consistent with a previous study which used the same neuronal model, SH-SY5Y cells, treated similarly with PA and found that Aβ peptides and BACE1 were increased [35]. In addition, a study in N2a neuronal cells given PA as a chronic supplement showed increases in Aβ peptides and p-Tau protein [45]. With regard to microglial cells, our study showed that increases in Aβ peptides and BACE1 levels, as well as a decrease in IDE were found, the exception being no impact on p-Tau proteins, when exposed to 200 and 400μM PA-conjugated BSA. In contrast, a previous study demonstrated that expression of Aβ peptides and p-Tau protein were unaltered while AβPP and total tau proteins were increased in BV2 microglial cells treated with PA [45]. The dissimilarity in these results may be due to the different strains of microglial cell used because the BV2 microglial cells were murine in origin not from humans as in the case of the HMC3 cells. Another reason of the variance could be the dose of the PA treatment. The previous study used 25μM of PA, which is significantly lower than the 200 and 400μM we used in our study. Interestingly, our findings demonstrated that the neuronal cells were more sensitive to PA than microglial cells as indicated by the upregulation of p-Tau proteins occurring only in neuronal cells. In this study, we determined the effects of PA on the expression of FcγRs, FcγR signaling proteins, AD-related proteins, proinflammatory cytokines, and cell viability of neurons and microglial cells. However, it should have the known controls such as stearic acid to compare the effects of PA on neuron and microglial cells that might lead to a deeper conclusion. We also demonstrated that FcγR blockage alleviated the emergence of AD pathologies in human neuronal and microglial cells following exposure to PA. Our results are substantiated by other literature, specifically that FcγR-/- mice had a reduction in tau hyperphosphorylation and Aβ accumulation both in the cortex and hippocampus of hypercholesterolemia models [46]. In addition, Fernandez-Vizarra and colleagues found that using FcγRIV blocking antibody attenuated p-Tau and Aβ in primary neurons from hypercholesterolemia mice [46].
Interestingly, our studies revealed that being in receipt of a diet high in this saturated fatty acid, as represented as PA-conjugated BSA, could induce inflammation in neuronal and microglial cells since these two brain cell models, SH-SY5Y and HMC3, received high concentrations of PA-conjugated BSA and showed significant increases in proinflammatory cytokine production, specifically TNFα, IL-1β and IL-6. In addition, the results indicated that the secretion of proinflammatory cytokines in HMC3 cells was obviously increased when compared to SH-SY5Y cells following exposure to 400μM of PA-conjugated BSA. A previous study has supported the idea that mRNA expression of TNFα and IL-1β was found to be significantly increased in BV2 microglial cells whereas it was not shown in N2a neuronal cells [37]. These findings are the reason behind microglia being the first and main form of active immune defense in the CNS [47]. Here, we also showed that FcγR blockage alleviated the inflammation which resulted in neuronal and microglial cells in response to these doses of PA. This result concurred with that published by a previous study which stated that TNFα was decreased in the cortex and hippocampus of gene deletion in FcγR mice under conditions of hypercholesterolemia [46].
However, the detailed mechanisms associated with the high-fat conditions which activate the upregulation of AD-related pathologies in association with FcγRs expression and activation in neuronal and microglial cells are not clearly understood. Therefore, this study provides evidence that giving high concentrations of PA-conjugated BSA for 24 h to microglial cells stimulates the expression of FcγRs levels, specifically FcγRII and FcγRIII. In human microglial cells, FcγRI, FcγRII and FcγRIII are expressed at very low levels under normal conditions [48]. It has been previously shown that an increase in level of both FcγRII and FcγRIII is present in the neurodegenerative condition [49]. Functionally, increased FcγRs in microglia in the high-fat condition could mediate the phagocytosis and release of inflammatory mediators [50, 51]. Our findings were consistent with a previous in vivo study which showed the upregulation of FcγRs gene expression in the hippocampus and cerebral cortex of hypercholesterolemic rats. However, in that study specific brain cell types such as neurons or microglia were not mentioned [46]. On the other hand, our findings clearly revealed that the microglial cell model receiving a defined concentration of PA-conjugated BSA led to an upregulation of FcγRII and FcγRIII levels, while in neuronal cells, FcγRII expression was not affected when exposed to the high-concentration of PA. Surprisingly, the activation of the FcγRs signaling protein, Syk, was increased in both neuronal and microglial cells in cases of high concentration of PA-induced AD progression. Moreover, the findings associated with FcγRs activation were shown to be associated with the upregulation of AD-related proteins, and proinflammatory cytokines in high concentration of PA treated neuronal and microglial cells in our study. Our study showed that the induction of FcγR blockage on neuronal and microglial cells reduced the activation of Syk resulting to decrease AD-related proteins and proinflammatory cytokines under PA-conjugated BSA exposure. Our study demonstrated that FcγR blockage could alleviate the activation of FcγRs which is associated with the reduction in the production of both AD-related proteins and proinflammatory cytokines in the PA condition. In addition to induce FcγRs proteins expression, PA also activated several signaling proteins including p-Syk and AD-related proteins in both neuronal and microglial cells. Moreover, previous study demonstrated that palmitate uptake by CD36 leads to the accumulation of deleterious lipids, such as diacylglycerol (DAG) which can activates protein kinase C (PKC) and NF-κB leading to increased inflammation [52]. Therefore, the inhibitor of these signaling proteins might be useful to inhibit the harmful effect of PA. However, this hypothesis still needs further investigation. Although our study demonstrated that PA is a risk factor of AD via the increased AD-related pathologies, inflammation and brain cell death associated with FcγRs activation, the other pathways might involve in this pathology. A previous study found that palmitic acid-BSA also enhances Aβ production through GPR40-mediated dual pathways in neuronal cells which involved with the Akt/mTOR/HIF-1α and Akt/NF-κB pathways [53]. Moreover, palmitic acid can induce neuron cell cycle G2/M arrest and endoplasmic reticular stress through protein palmitoylation in SH-SY5Y human neuroblastoma cells [35]. Therefore, other mechanisms implicated in the effect of palmitate remain to be further investigated.
The results from our study demonstrated that the high-saturated fat condition as caused by high concentration of PA is contributory to the pathology of AD and affected both neuronal and microglial cell death in a dose-dependent manner. In support of our findings, there is an accumulation of evidence to show that neuronal and microglial cell death occurs when they are exposed to PA at an appropriate dose and time of treatment [35, 54–56]. Nevertheless, there are some discrepancies between the findings associated with these two types of brain cell. When exposed to the same concentration of PA-conjugated BSA, the percentage of cell viability was lower in SH-SY5Y cells than in HMC3 cells indicating that neuronal cells were more susceptible to PA-conjugated BSA, especially at a high concentration. Markedly, only SH-SY5Y cells treated with PA-conjugated BSA had increased p-Tau protein expression while p-Tau protein expression was unaltered in HMC3 cells. These findings may explain the results associated with cell death, because neuronal cells were affected more severely than microglial cells in terms of cell viability as indicated by fewer viable cells when exposed to the same dose and time of PA-conjugated BSA. Our findings also indicated that the blockage of FcγR increased the percentage of cell viability in both human neuronal and microglial cells even if these cells are exposed to PA conditions.
In conclusion, our findings demonstrated that PA significantly increased AD-related proteins, inflammation, FcγRs expression, and their activation, as well as decreased cell viability in SH-SY5Y and HMC3 cells. Our findings imply that the viability of neuronal and microglial cells under high PA condition was associated with the presence of AD-related proteins, proinflammatory cytokines, and the activation of FcγRs signaling. It is possible that the activation of FcγRs might be due to Aβ clearance, and this clearance could result in the generation of inflammatory environments which could lead to cell death. Our results also indicated that FcγR blockage could alleviate the activation of FcγRs which is associated with the reduction in the production of both AD-related proteins and proinflammatory cytokines in the PA condition. Eventually, these events could lead to the reduction of cell death. These findings suggest that inhibition of FcγRs might be a promising approach to further investigate the way to attenuate neuroinflammation and AD in HFD-induced cognitive decline. However, this study still has a lot of limitation, since the present study used only human neuroblastoma SH-SY5Y cells and microglial HMC3 cells, which is not fully represented animal and clinical studies. Therefore, an in vivo and clinical studies are still need for future investigation to confirm the results of the present study.
Footnotes
ACKNOWLEDGMENTS
This work was supported by Senior Research Scholar Grant from the National Research Council of Thailand (SCC), a NSTDA Research Chair Grant from the National Science and Technology Development Agency Thailand (NC), the Chiang Mai University Excellent Center Award (NC) and Thailand Science Research and Innovation (TSRI)-CMU (SK). The authors acknowledge Prof. Dr. Watchara Kasinrerk, Biomedical Technology Research Center, Chiang Mai, Thailand, for providing isotype matched control antibody. The authors acknowledge Dr. Supansa Pata and Dr. Witida Laopajon, Biomedical Technology Research Center, Chiang Mai, Thailand, for the help with flow cytometry instruction. Ms. Sasiwan Kerdphoo and Ms. Thidarat Jaiwongkum are acknowledged for laboratory technical assistance.