
Abstract
Background:
Chinese Herbal Medicines (CHMs), as an important and integral part of a larger system of medicine practiced in China, called Traditional Chinese Medicine (TCM), have been used in stroke therapy for centuries. A large body of studies suggest that some Chinese herbs can help reverse cognitive impairment in stroke patients, while whether these herbs also exert therapeutic benefits for Alzheimer’s disease remains to be seen.
Objective:
To address this issue, we selected four types of CHMs that are commonly prescribed for stroke treatment in clinical practice, namely DengZhanXiXin (D1), TongLuoJiuNao (T2), QingKaiLing (Q3), and HuangQinGan (H4), and tested their effects on amyloid-β protein precursor (AβPP) processing in vitro.
Methods:
AβPP, β-secretase (BACE1), and 99-amino acid C-terminal fragment of AβPP (C99) stably transfected cells were used for the tests of AβPP processing. The production of Aβ, activity of BACE1, neprilysin (NEP), and γ-secretase were assessed by ELISA, RT-PCR, and western blot.
Results:
By upregulating BACE1 activity, D1 increased Aβ production whereas decreased the ratio of Aβ42/Aβ40; by downregulating BACE1 activity and modulating the expression of γ-secretase, T2 decreased Aβ production and the ratio of Aβ42/Aβ40; by downregulating BACE1 activity, Q3 decreased Aβ production; H4 did not change Aβ production due to the simultaneously downregulation of BACE1 and NEP activity.
Conclusion:
Our study indicates that these four anti-stroke CHMs regulate AβPP processing through different mechanisms. Particularly, T2 with relatively simple components and prominent effect on AβPP processing may be a promising candidate for the treatment of AD.
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disorder, characterized by the progressive formation of insoluble amyloid plaques and vascular deposits consisting of the amyloid-β peptide (Aβ) in the brain. Amyloid-β protein precursor (AβPP) is cleaved by two enzymes: β-secretase (BACE1) and γ-secretase to generate excessive Aβ. Despite that sheer amount of evidence indicating the abnormal processing of AβPP as one of typical pathological features of AD [1, 2], one is still unsure whether AD primarily results from the abnormality of the AβPP or from the dysregulation of the inflammation system [3, 4].
AD and stroke are both common in the elderly, but their association is rather complex and has not been fully defined. Previous studies showed that stroke patients might be more likely to suffer from memory decline [5, 6]. Additionally, stroke increases the risk of AD [7]. Stroke leads to a hypoxic environment in the brain, which facilitates AD pathogenesis by upregulating BACE1 gene transcription and markedly increases Aβ deposition and neuritic plaque formation, as well as potentiates the memory deficit in AβPP transgenic mice [8, 9]. Our previous study showed that the death of brain cells caused by stroke or head injury may induce generation of Aβ via the aggregation of BACE1 protein [10], raising the perspective that therapeutic intervention in this process may lower the risk of stroke-associated AD.
Traditional Chinese Medicine (TCM) has long been used in stroke therapy and its therapeutic efficacy has been identified by extensive clinical studies [11–15]. Considering the complicated interaction between stroke and AD, the possibility that anti-stroke CHMs may also have an effect on AβPP processing and Aβ production is intriguing. Here, we selected four types of CHMs from the ginsenoside family: DengZhanXiXin (D1), TongLuoJiuNao (T2), QingKaiLing (Q3), and HuangQinGan (H4). In clinical practice, these CHMs are commonly administrated by injection, and several studies suggest that they could be prescribed both in stroke and dementia treatment [16–23]. In addition, evidence also suggests they could alleviate plaque burden and Aβ pathology in AD animal models [24–31].
With the anti-Aβ agent officially approved by the FDA, drug development is focusing on the generation and clearance of Aβ once again. Herein, using AβPP, C99, and BACE1 transfected cell lines, we investigated the effect of four anti-stroke CHMs on the AβPP processing and Aβ clearance, and revealed the possible mechanism in vitro.
MATERIALS AND METHODS
Drug preparation
D1 was purchased from Yunnan BioValley Pharmaceutical Co. Ltd., China (Coad No. by SFDA: Z53021569); T2 was provided by Tianjin Chase Sun Pharmaceutical Co. Ltd., China (Cat. No. 090602); Q3 was purchased from Shineway Pharmaceutical Group Ltd., China (Coad No. by SFDA: Z13020880); H4, Ginsenoside (Gin), Geniposide (Gen), and PNS were purchased from National Institute for the Control of Pharmaceutical and Biological Products, China (Cat. No. 110842, Cat. No. 110703, Cat. No. 110749, Cat. No. 110870). Gin and Gen were dissolved in DMEM to make 10μg/ml stock.
Cell culture and CHMs administration
293 cells were obtained from ATCC and maintained in 10%FBS DMEM, AβPP stable transfected cells were maintained in 200μg/ml G418. pcDNA-BACE1 stable transfected cells were maintained in 200μg/ml Zeocin. C99 expression vector was constructed, simply C99 DNA fragment was ligated to AβPP signal peptide, cloned into pFLAG-CMV vector. Transfection was performed by lipofectamine2000 according to the manufactory’s instruction. The toxicity threshold of each drug to 293 cells was tested, doses that lower than the threshold were used in the experiments (data not shown). CHMs were directly mixed with the medium containing 10%FBS DMEM and then added to the cells. 72 h post-administration, medium was collected for Aβ ELISA assay.
Aβ ELISA
Aβ40 and Aβ42 levels were measured by the ELISA kit (KHB3481 and KHB3544, Biosource, USA). The ELISA system has been extensively tested, and no cross-reactivity between Aβ40 and Aβ42 was observed (data no shown). Data are presented as means±SEM of the three experiments. Control group without drug treatment was defined as 100%and the relative level after treatment was calculated and shown as the percentage of the control group
BACE1 and NEP activities
For BACE1 activity, cells were homogenized in lysis buffer (10 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM Na3VO4, 10%Glycerol, 0.5%Triton X-100). BACE substrate (Calbiochem, Germany) was dissolved in DMSO and mixed with reaction buffer (50 mM HAc, 100 mM NaCl; pH 4.1) to make final concentration to 2.5μM. An equal amount of protein was mixed with 100μl of substrate, and fluorescence intensity was measured with a microplate reader at an excitation wavelength of 430 nm and an emission wavelength of 520 nm. Maximal enzymatic speeds were calculated and the samples without drug treatment were regarded as 100%, relative activity percentage was calculated.
NEP enzyme activity was measured by quenched fluorogenic substrate as described previously [32]. Cells were homogenized in 100 mM MES pH 6.5 buffer containing 200μM PMSF and proteinase inhibitor mix (Sigma, USA). The resultant homogenate was directly used in NEP activity assay. The hydrolysis of the fluorogenic substrate peptides Mca-RPPGFSAFK(Dnp)-OH (R&D systems) in 100 mM MES buffer, pH 6.5 at concentration of 5μM was measured by following the increase in fluorescence (excitation at 342 nm and emission at 420 nm) that occurred upon peptide bond cleavage. Fluorescence was read for 1 h, 5 min/reading. The max velocity of NEP activity was calculated by the first 20 min. Samples without drug treatment were regarded as 100%, relative activity percentage were then calculated.
Western blot
50μg of the total protein was separated on SDS-PAGE, and then transferred to PDVF membrane by semi-dry transfer equipment. The blot was probed by anti-BACE1 antibody [33]; anti-APH1 polyclonal antibody (kind gift from Dr. Yuming Li, Memorial Sloan-Kettering Cancer Center, New York); anti-Nicastrin polyclonal antibody (kind gift from Dr. Gang Yu, University of Texas Southwestern Medical Center); anti-PS1 N-terminus polyclonal antibody (kind gift from Dr. Sangram Sisodia, University of Chicago); anti-β-actin antibody (Sigma, USA).
RT-PCR
Total RNA was extracted drug treated 293 cells by RNeasy mini kit from Qiagen. For BACE1, forward primer is 5′ AGGGAGCATGATCATTGGAG, backward primer is 5′ CGTGGATGACTGTGAGATGG to amplify a 475bp fragment. For human NEP RT-PCR, primers 5′ GGA CTC GAC TGG AGA TCA GC and 5′ CCA AGT CGA GGT TGG TCA AT were used to amplify region from 83–686 nt from NEP mRNA. For β-actin RT-PCR, the primers are: forward 5′- GGACTTCGAGCAAGAGATGG, backward 5′ GAAGCATTTGCGGTGGAG to amplify the β-actin mRNA coding region from 633 to 1125.
Statistical analysis
All data are presented as mean±SEM. Paired or unpaired Student’s t-test (for two group means) or one or two-way analysis of variance (ANOVA) with post-hoc Tukey test, Scheffe’s test, or Bonferroni test, as appropriate, were conducted using Prism. The significance level for the two-side analyses was set at p≤0.05.
RESULTS
Anti-stroke CHMs have different effect on Aβ production
293 human embryonic kidney cells stably transfected with AβPP695 carrying Swedish AβPP mutation and secreting high total Aβ were treated with different dose of four CHMs. The Swedish mutation makes AβPP a superior substrate for BACE1 and elevates the production of both secreted Aβ40 and Aβ42 in the medium, allowing an accurate quantification by sandwich ELISA. Therefore, this system is suitable to assess the influence of CHMs on Aβ production.
After 72 h treatment with different doses, we found that T2 reduced Aβ40 by 5–27%and Aβ42 by 30–50%, while Q3 reduced Aβ40 0–20%and Aβ42 by 0–19%. In contrast, D1 increased Aβ40 by about 18–37%and Aβ42 by 10–32%. All these effects were in a dose-dependent manner. H4 did not significantly change the Aβ40 and Aβ42 levels (Fig. 2A, B). In addition, we observed the ratio of Aβ42/Aβ40 after D1 and T2 administration were both significantly decreased (Fig. 2C).
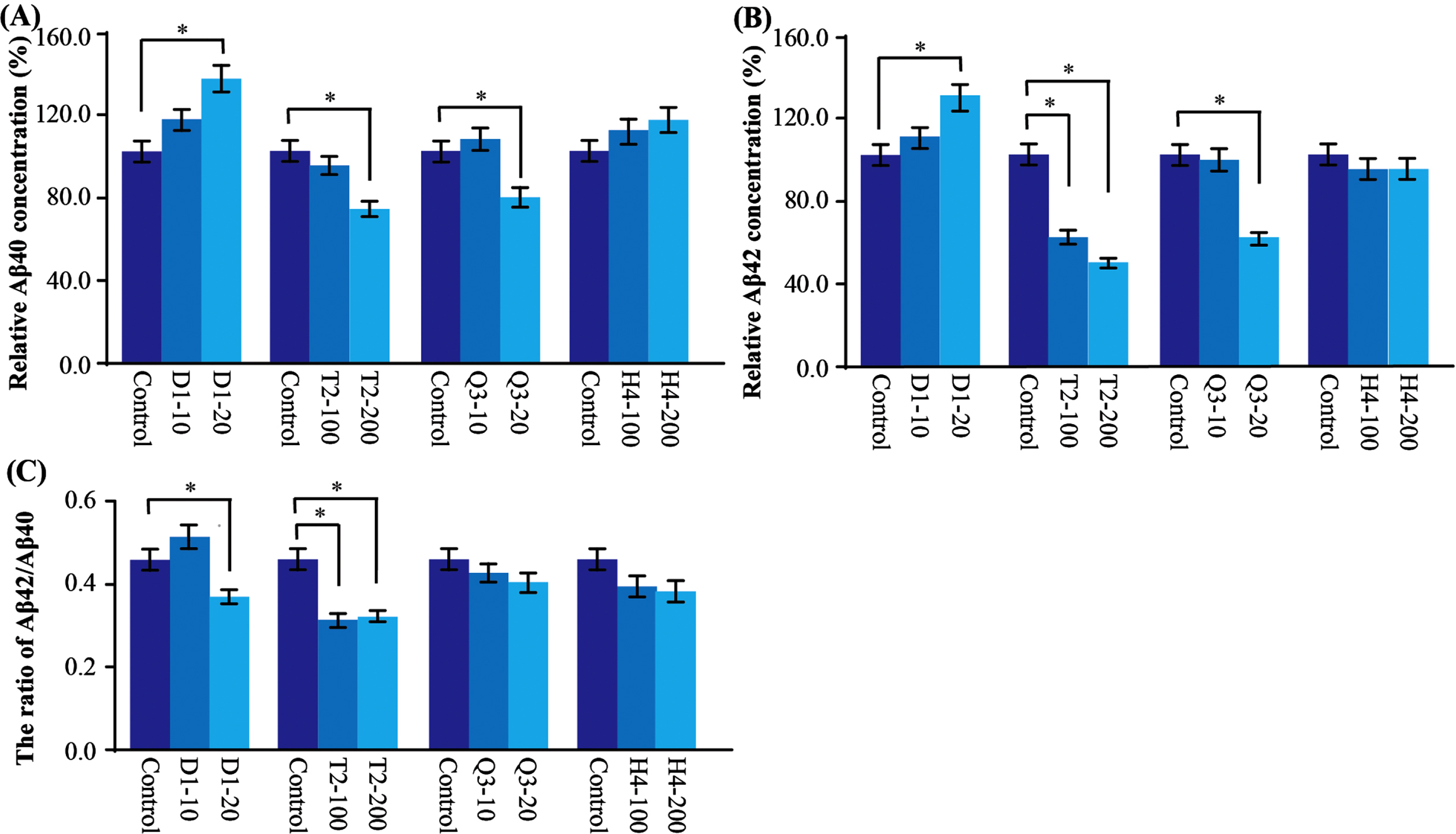
Relative Aβ40 and Aβ42 levels in AβPP stably transfected cells. A) The level of Aβ40 was significantly decreased after T2 and Q3 administration and increased after D1 administration in a dose dependent manner. B) The level of Aβ42 was significantly decreased after T2 and Q3 administration and was increased after D1 administration in a dose dependent manner. C) The ratio of Aβ42/Aβ40 was significantly reduced in D1 and T2 groups. Data are presented as mean±SEM. Compared with control, *p < 0.05.
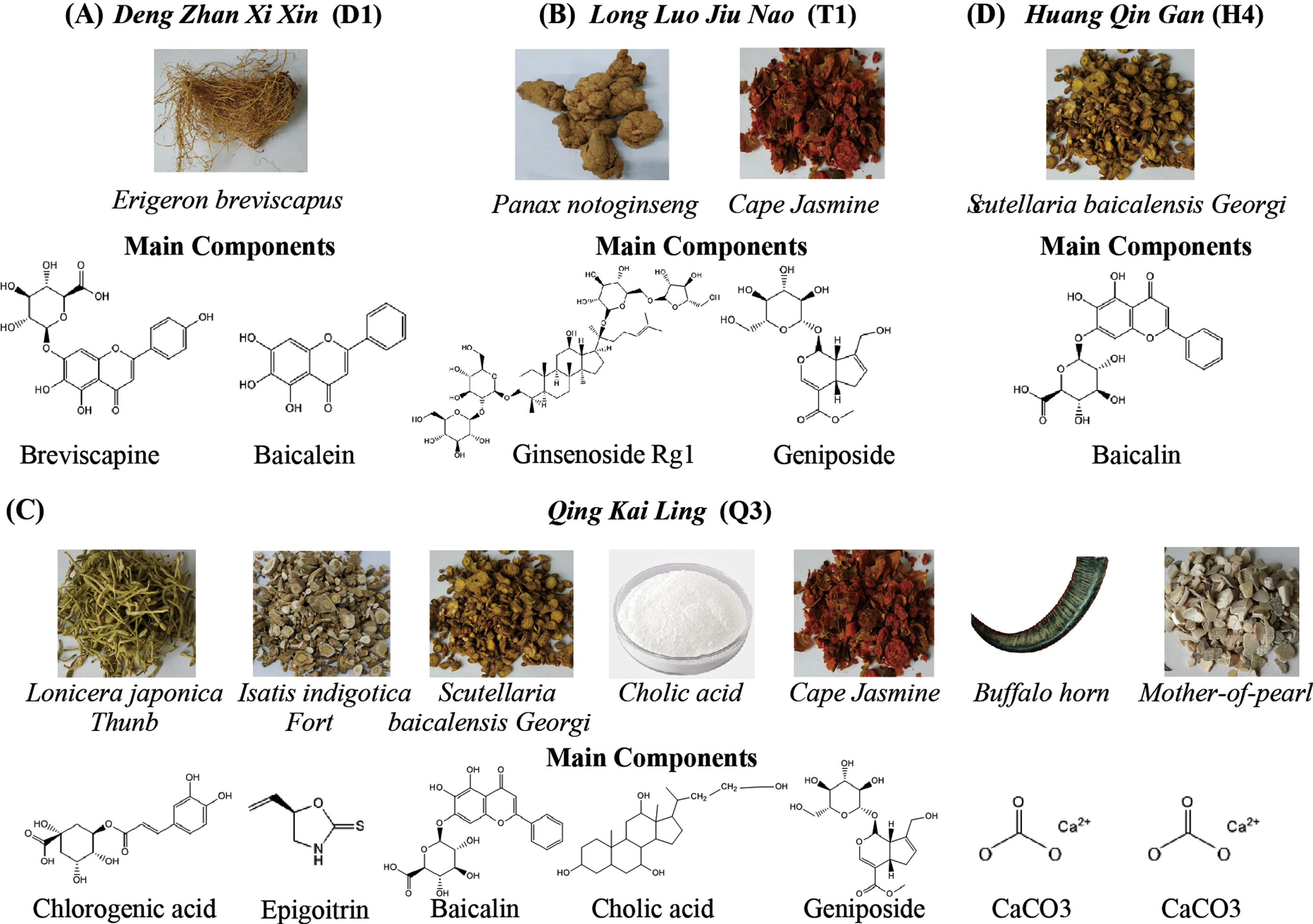
The major active components of the four anti-stroke CHMs. A) The components of DengZhanXiXin (D1). B) The components of TongLuoJiuNao (T2). C) The components of QingKaiLing (Q3). D) The component of HuangQinGan (H4).
Anti-stroke CHMs modulate AβPP processing: BACE1
Since we found that these CHMs showed different effects on Aβ production, we next tested whether this regulation is achieved through the modulation of AβPP processing.
Two key enzymes are involved in AβPP processing: β-secretase (BACE1) [34–36] and γ-secretase [35, 36]. BACE1 is well known to be implicated in the first step of producing Aβ peptide. Recently, it was reported that BACE1 activity was increased in the brain of AD [33, 37] and cerebrospinal fluid of mild cognitive impairment [38]. In this study, we first tested whether these CHMs could regulate the BACE1 activity in AβPP transfected cells. We found that three of the four CHMs, including T2, Q3, and H4, significantly decreased BACE1 activity despite that only T2 and Q3 decreased the Aβ production. Quantitatively, T2 and Q3 decreased BACE1 activity by about 25%, and H4 by about 34%. However, BACE1 activity was increased by about 14%in the D1 group (Fig. 3A), which is consistent with the result of increased Aβ by D1 (Figs. 3A 2).
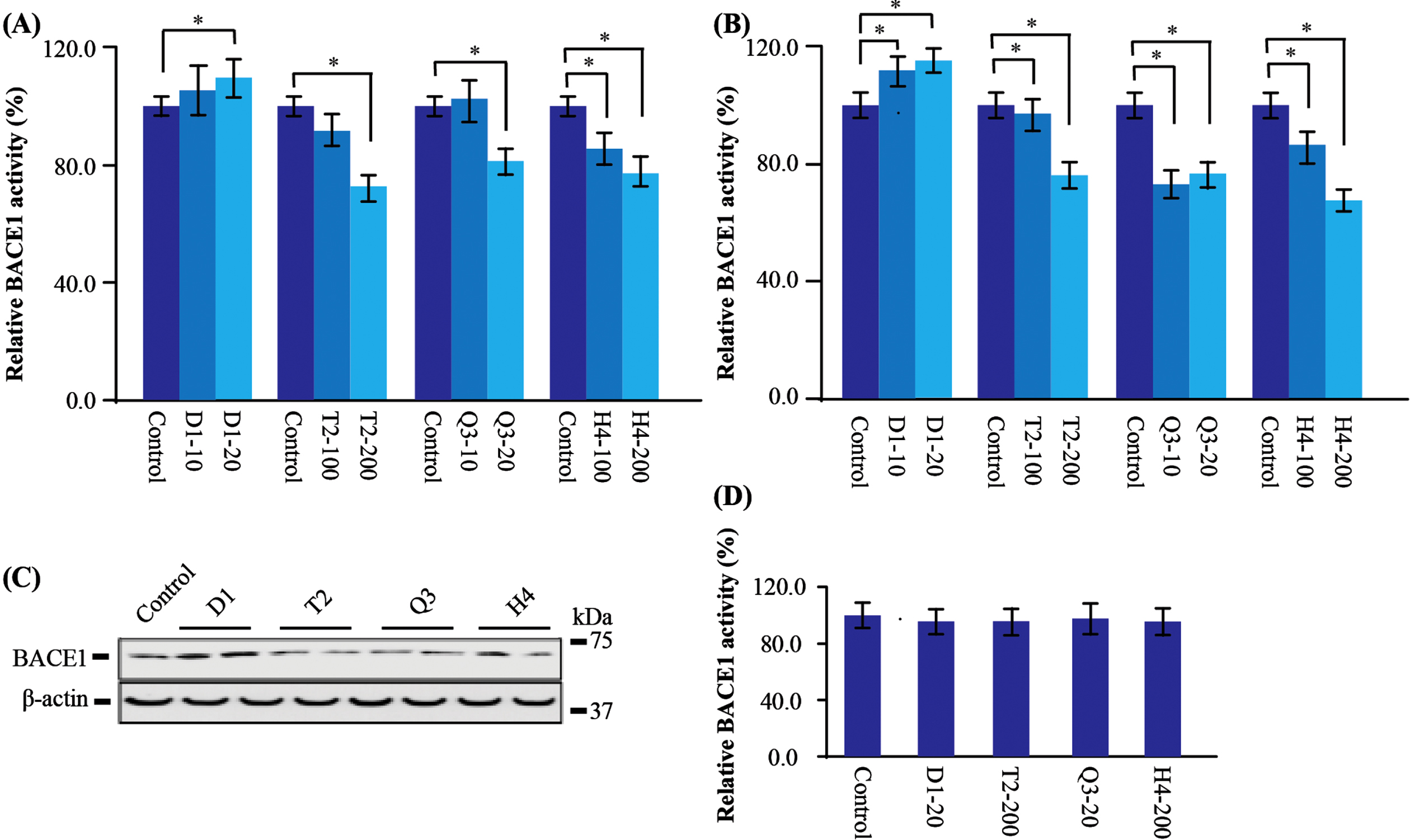
CHMs affect BACE1 expression and activity. A) In the AβPP stably transfected cells, the BACE1 activity was significantly increased after D1 administration, but was decreased after T2, Q3, and H4 administration. B) In the BACE1 stable transfected cells, the BACE1 activity was significantly increased after D1 administration, while decreased after T2, Q3, and H4 administration. C) In the BACE1 stably transfected cells, the protein level of BACE1 was higher after D1 administration but had a tendency to be lower after T2, Q3, and H4. D) Direct effect of CHMs on BACE1 activity. BACE1 activity assay was evaluated in the presence of CHMs in BACE1 stably transfected cells. There was no significant difference among all groups. Data are represented as mean±SEM. Compared with control, *p < 0.05.
In light of these results, we further found there was no change of BACE1 at mRNA level (data not shown). With regard to the protein level, BACE1 stably transfected cell expressing high level of BACE1 protein was used and BACE1 activity was evaluated again. Consistent with results in AβPP transfected cells, we found D1 increased the activity of BACE1 by about 13%. In contrast, T2 decreased BACE1 activity by about 26%, Q3 by about 27%and H4 by about 34%(Fig. 3B). Besides, the western blot analysis was coincided with the activity assay that D1 increased BACE1 protein expression, while the others decreased BACE1 expression (Fig. 3C). This indicates that D1 could enhance BACE1 activity by upregulating its protein level, which eventually leads to the accumulation of Aβ production.
To rule out the possibility that these CHMs could be interact with BACE1 and inhibit its activity, We directly mixed CHMs within the extracted purified BACE1. The concentration of the CHMs was same as the highest concentration used in the cell treatment. We did not find any direct regulation of BACE1 activity by mixing the drugs to the reaction (Fig. 3D). These results indicate that these four anti-stroke CHMs are involved in the biological process in the cell model and modulate the BACE1.
Anti-stroke CHMs modulate AβPP processing: γ-secretase
AβPP can be cleaved by BACE1 to produce a short C-terminal fragment (CTF) C99, which is in turn processed by γ-secretase to produce pathogenic Aβ peptide [35, 36]. To further identify which step of the AβPP processing that the CHMs regulate, a C99 transfected cell containing only γ-secretase processing site, was used to examine the effect of CHMs on Aβ production. There was no significant change of Aβ40 and Aβ42 levels of cells treated with D1, Q3, and H4; however, significantly decreased Aβ40 and Aβ42 levels were observed in the T2 group (Fig. 4). Quantitatively T2 decreased Aβ40 by 23–41%and Aβ42 by 22–27%, suggesting T2 might regulate the expression or activity of γ-secretase.
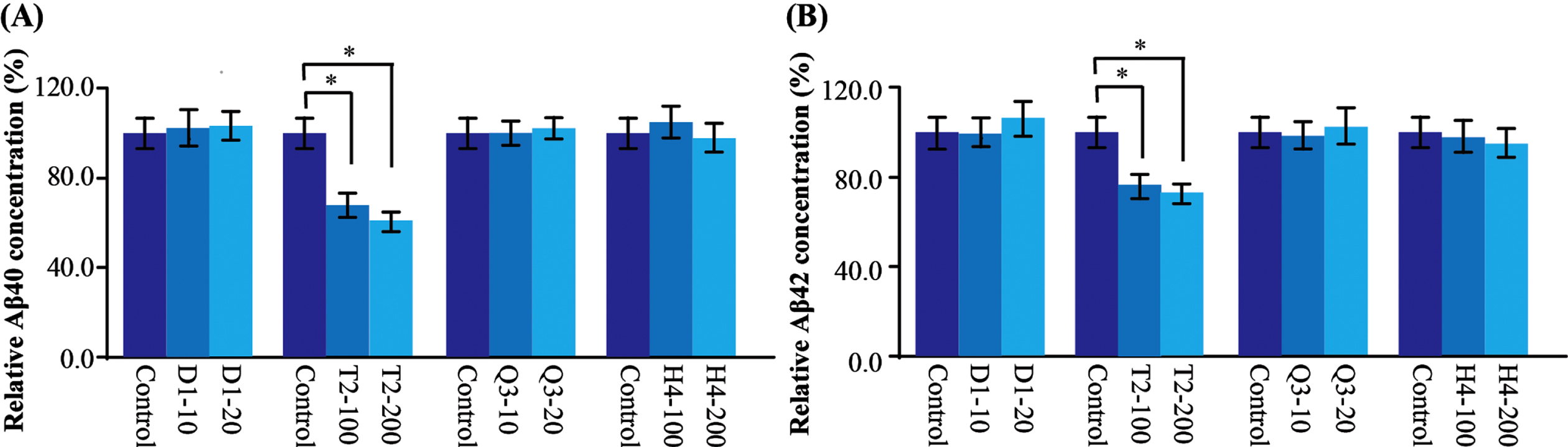
CHMs affect γ-secretase activity. A) In the C99 transfected cells, the production of Aβ40 was significantly decreased after T2 administration. B) In the C99 transfected cells, the production of Aβ42 was significantly decreased after T2 administration. Data are represented as mean±SEM. Compared with control, *p < 0.05.
Anti-stroke CHMs modulate clearance pathway: NEP
Now that H4 decreased BACE1 activity while the Aβ production was not changed, we inferred that H4 might affect the follow-up process after the generation of Aβ. One such process could be NEP, an Aβ degradation enzyme [39, 40]. NEP activity assay [41] was performed to evaluate the NEP activity in AβPP transfected cell. After H4 administration, NEP activity was significantly decreased (Fig. 5A). Considering the influence of H4 on BACE1 activity from earlier study, the unaltered Aβ level may be ascribed to the simultaneously downregulation of NEP and BACE1 activities.
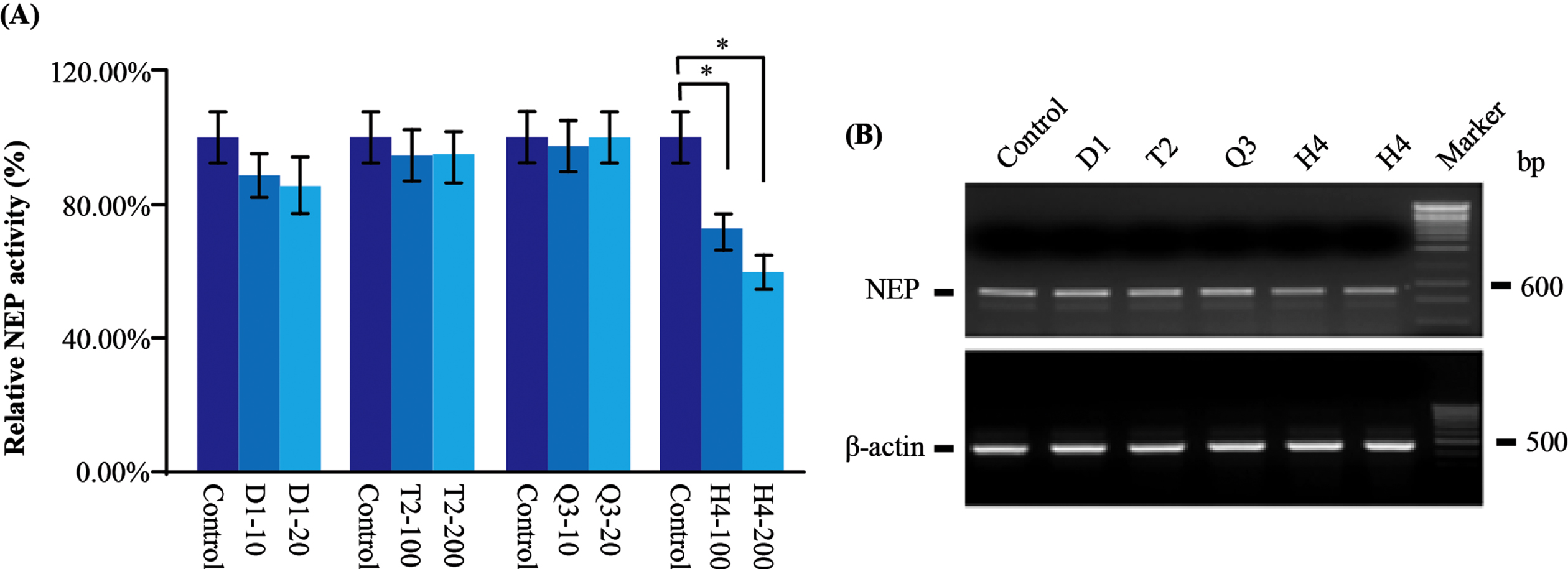
CHMs affect NEP activity and its mRNA level. A) In the AβPP stably transfected cells, only H4 significantly reduced the NEP activity. B) By the analysis of RT-PCR, again, the mRNA level of NEP was reduced after H4 administration. Data are represented as mean±SEM. Compared with control, *p < 0.05.
Additionally, there was a decreased tendency of NEP activity in D1 treated cells. Quantitatively D1 decreased NEP by about 15%, while no significant change was observed in T2 and Q3 treated cells (Fig. 5A). To further illustrate how CHMs regulate NEP activity, we conducted RT-PCR to test NEP at mRNA level. As expected, the level of NEP mRNA was reduced after H4 administration (Fig. 5B), indicating H4 could downregulate NEP transcription to reduce NEP activity.
T2 affects AβPP processing and Gin, which is the active component responsible for the reduced BACE1 level and activity
T2 and Q3 exhibited consistent results regarding BACE1 activity, protein, and Aβ production. Gin and Gen are reported to be the main components of T2. Q3 also contains Gin while its composition is rather complicated compared to T2. We focused on these two compounds and tried to distinguish which one is responsible for the reduced Aβ level in T2 treated cells.
Cells can be treated purified Gin as much as 1μg/ml without obvious toxicity (data not shown); for Gen, no obvious toxicity could be observed at the concentration up to 0.25μg/ml (data not shown). In AβPP stably transfected cells, a different dosage of Gin and Gen were administrated. In the Gin group, we found that Aβ40 level decreased by 34%and 50%in a dose-dependent manner. In contrast, Aβ40 in the Gen group increased by 4%and 16%in a dose-dependent manner (Fig. 6A).
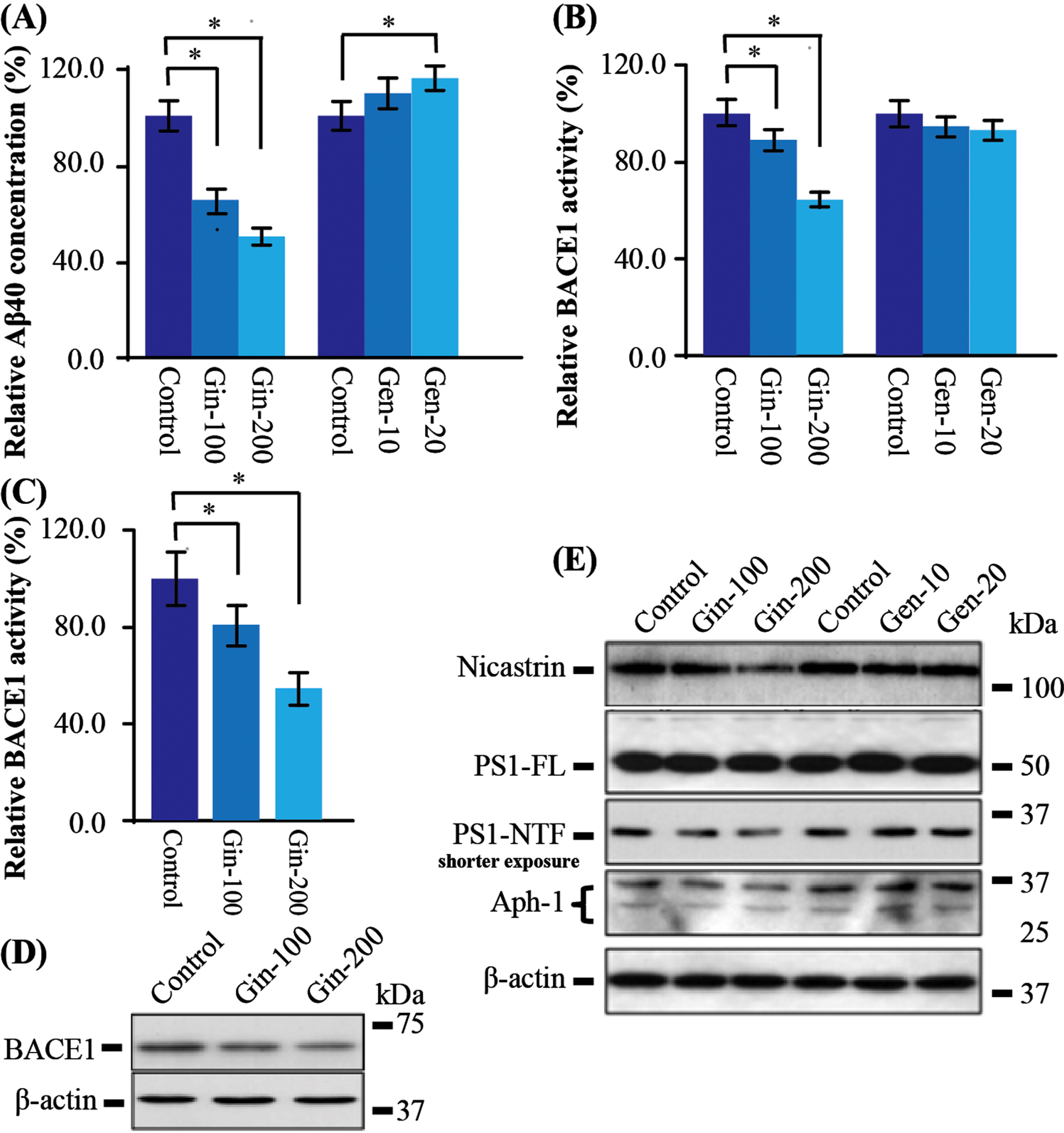
The major component of T2, Ginsenoside (Gin), mainly contributes to the modulation of the BACE1 activity and expression. A) In the AβPP stably transfected cells, Gin significantly reduced the production of Aβ40 in a dosage manner; Geniposide (Gen) significantly increased Aβ40 production in a dose-dependent manner. B) The BACE1 activity was significantly reduced after Gin administration, while there was no significant change after Gen treatment. C) In the BACE1 stably transfected cells, again, Gin significantly reduced the BACE1 activity. D) The protein level of BACE1 was significantly reduced after Gin administration. E) In the C99 stably cells, there was no significant change in western blot analysis of γ-secretase subunits. But Gin slightly reduced the level of Nicastrin, PS1-NTF, and Aph-1 in a dose-dependent manner. Data are represented as mean±SEM. Compared with control, *p < 0.05.
In order to find out whether Gin also affects BACE1 activity as T2, we further tested the BACE1 activity in these Gin-treated cells. We found that Gin reduced the BACE1 activity by 12%and 35%in a dose-dependent manner, while Gen did not alter BACE1 activity significantly (Fig. 6B). The results testified that Gin reduced Aβ production through downregulating BACE1 activity. Meanwhile, Gen showed no effect on BACE1 activity and could increase Aβ level.
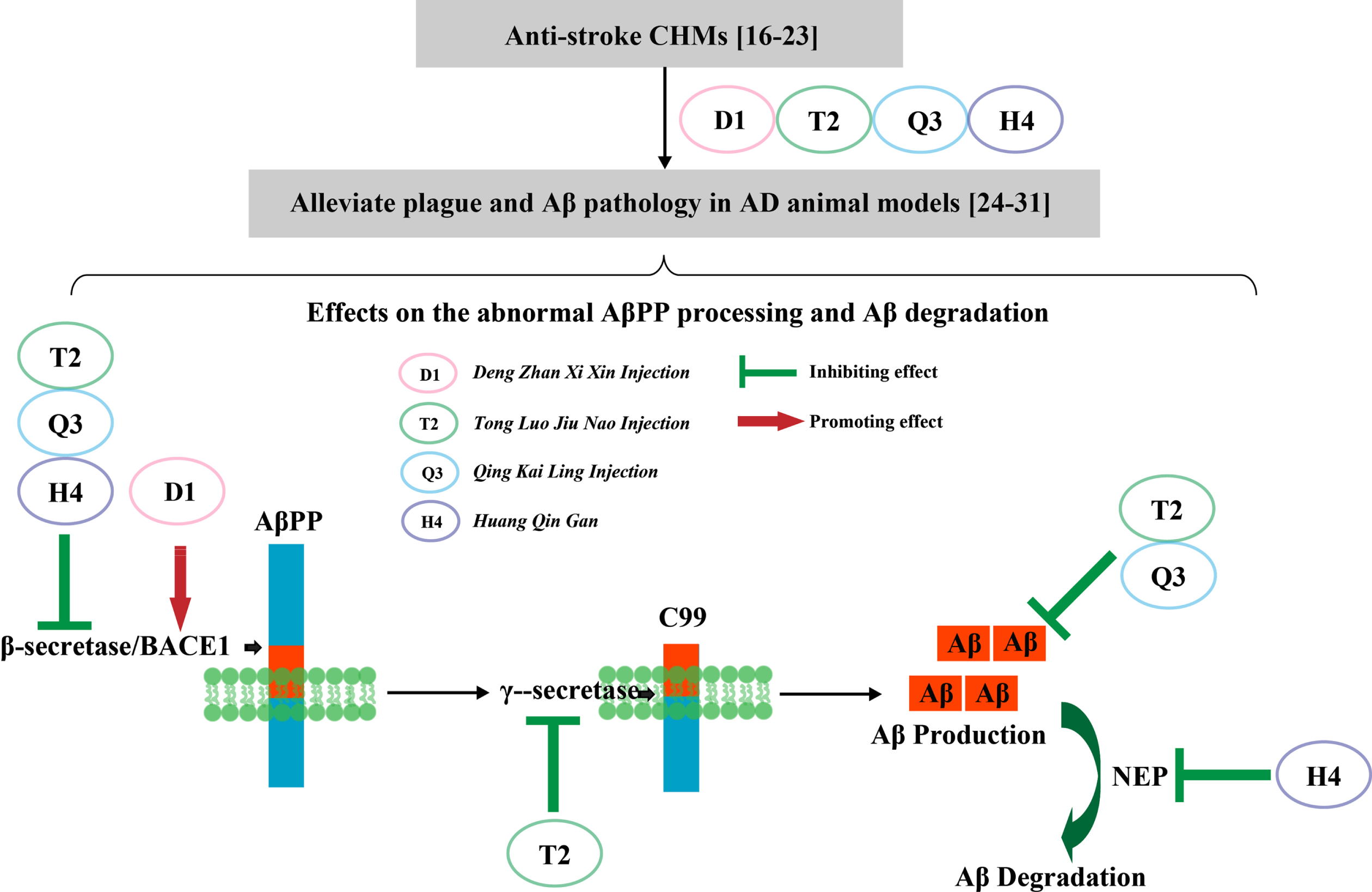
Four anti-stroke CHMs affect AβPP processing by different mechanisms. In clinical practice, studies supported that these CHMs could be prescribed both in stroke and dementia treatment. Related studies also suggested they could alleviate plaque burden and Aβ pathology in experimental AD animal models. In this study, we found that D1 increased Aβ production, while decreased the ratio of Aβ42/Aβ40 by upregulating BACE1 activity; T2 decreased Aβ production and Aβ42/Aβ40 ratio by downregulating BACE1 activity and modulating γ-secretase expression; Q3 decreased Aβ production by downregulating BACE1 activity; H4 did not change Aβ production due to the simultaneously downregulation of BACE1 and NEP activity.
It should be noted that there was no significant change of BACE1 at the mRNA level in Gin-treated cells (data not shown). To prove that Gin could modulate BACE1 at the protein level, we treated BACE1 stably transfected cells with Gin. As expected, BACE1 activity decreased by 21%and 45%in a dose-dependent manner (Fig. 6C). Western blot also showed that BACE1 protein was significantly decreased in a dose-dependent manner (Fig. 6D). These results are coincided with the results of T2-treated BACE1 stably transfected cell, indicating Gin is the active component of T2 to reduce BACE protein level and activity.
Since T2 also decreased Aβ level in C99 transfected cell, it suggests T2 could regulate γ-secretase pathway. We did western blot analysis of the three γ-secretase components, including Nicastrin, PS1, and APH1. The results showed that PS1 full length protein (PS1-FL) were not changed. However, after a higher concentration administration and shorter exposure, PS1 N-terminal fragment (PS1-NTF) was decreased. Similarly, PS1-NTF, APH1, and Nicastrin were also reduced (Fig. 6E). These results indicates that high concentration of Gin could decrease the major components of the γ-secretase to impair its function. To sum up, Gin plays the crucial role in AβPP processing through the regulation of BACE1 and γ-secretase.
DISCUSSION
In this study we tested the effects of the four CHMs that have been used in stroke treatment on AβPP processing. D1 is a preparation extracted from the herb Erigeron breviscapus, and was approved by the Chinese Food and Drug Administration and listed by the Chinese Pharmacopoeia in 2005 [16]. The benefits of D1 for ischemic stroke have been reported in clinical trials [17, 42]. T2 is a combined preparation of Gardenia jasminoides fruits and Panax notoginseng roots, and its major components are geniposide and ginsenoside. T2 has been well-regarded by the inspiring effectiveness in patients with vascular dementia and cerebral ischemic stroke [18]. Q3, also listed in the Chinese Pharmacopoeia, is modified from a famous ancient Chinese patent medicine Angong Niuhuang pills, which has great antipyretic and anti-inflammatory effects and is widely used for the acute stage of cerebrovascular disease [19]. The major active components of Q3 are cholalic acid, deoxycholic acid, jasminoidin, and baicalin [20]. T4, a flavonoid isolated from the root of herb Scutellaria baicalensis Georgiis, is an active component of several CHMs used for the treatment of ischemic stroke in the clinic [21]. It also is widely applied for the stroke management due to its anti-oxidant and anti-excitotoxic effects [22, 23].
Evidence suggests that these CHMs may also alleviate plaque burden and Aβ pathology in AD animal models [24–31]. In AβPP/PS1 mice, D1 could improve learning and memory, as well as reduce Aβ burden by downregulating BACE1, promoting Aβ-degrading enzyme IDE and suppressing receptor for advanced glycation end products (RAGE) [24]. The main active components of D1, caffeic acid ester fraction and scutellarin, have been reported to improve cognitive function and exert neuroprotective effect in animal model. Besides, in the SH-SY5Y cell line, scutellarin efficiently inhibited the oligomerization and fibril formation, as well as reduced Aβ oligomer-induced toxicity [25, 26]. The pharmacokinetics study using software and multiple data processing approaches systematically analyzed the chemical constituents of D1 and its prototypes and metabolites in vivo. HPLC showed that after D1 injected to rats over the dose ranges of 10–40 mg/kg, the plasma concentration-time curve of scutellarin was best conformed to three-compartment open model. The Area Under Curve (AUC) was proportional to dose indicating a linear pharmacokinetics characteristic of scutellarin after injection [43, 44]. For T2, it was reported that T2 reduced amyloidogenic processing of AβPP in AD mouse model [27]. In addition, T2 could improve spatial learning and memory function in 10-month-old AβPP V717I transgenic mice and could reduce anxiety [45, 46]. T2 is primarily composed of two active components: Gin and Gen. In terms of the pharmacokinetics, the AUC(0⟶∞) values of Gen were 6.99±1.27 h·μg/ml after injection with the dose of 10 mg/kg based on HPLC method in the rat model [47, 48]. Gin could penetrate the blood-brain barrier and blood-cerebrospinal fluid barrier, and then immediately distribute to learning and memory-related regions in rats brain [28]. Q3 is widely used in acute stage of cerebrovascular disease including ischemia stroke due to its regulation of several signal pathway or target such as toll-like receptor and PI3K-Akt [49, 50]. It offers the neuroprotective benefits for acute ischemia patients by inhibiting the adhesion of white blood cell and endothelia cell, relieving the damage of vascular endothelial cell as well as inhibiting the process of inflammation [51, 52]. In the SAMP8 AD mouse model, Q3 improved the learning and memory ability probably due to its function in increasing cerebral Ach and 5-HT and activating the PI3K/AKT pathway [29]. According to HPLC-UV analyzing the main components profiles of Q3, nine components (uridine, adenosine, guanosine, neochlorogenic acid, chlorogenic acid, caffeic acid, gen, baicalin, and 4,5-Di-O-caffeoylquinic acid) were identified [53]. By high-performance liquid chromatography with tandem mass spectrometry, 18 prototype compounds and 19 metabolites were identified in rat urine, providing essential data for pharmacokinetics of Q3 [54]. For H4, it attenuated cerebral ischemia via anti-oxidative and anti-apoptotic pathways [55]. Accompanied with jasminoidin and cholic acid, it further improved the cognitive function in ibotenic acid-induced rat models. In AD animal models, H4 ameliorated Aβ142 protein-related pathology and cognitive dysfunction [31]. The pharmacokinetics study showed that Baicalin was absorbed rapidly into the plasma and maintained its concentration at a certain range in plasma due to its high protein-binding rate, which was found to be 86–92%[56]. Together, these studies provide solid evidence that the four anti-stroke CHMs have the potential to treat AD; their main components could be distributed throughout the body and effectively alleviate plaque burden and Aβ pathology in vivo.
With an anti-Aβ agent officially approved by the FDA [57–61], drug development is focusing on the generation and clearance of Aβ once again. In our study, D1 increased Aβ level by elevating the expression of BACE1 protein and enhancing its enzymatic activity, while the ratio of Aβ42/Aβ40 was decreased. For H4, it reduced the protein expression and activity of BACE1, as well as the expression of the enzyme for Aβ clearance, NEP. The modulation of the BACE1 and NEP cancelled each other out, hence there was no obvious change of Aβ production. Due to the complex mechanisms of both stroke and AD, the result that the H4 was not effective on AβPP processing did not indicate that it could not relieve the symptoms. The exact mechanism of H4 in restoring memory needs to be studied further. Q3 reduced Aβ level by downregulating BACE1 activity and expression. T2 was the most effective one in reducing Aβ level and Aβ42/Aβ40 ratio in vitro, partially through downregulating BACE1 level and activity. Gin, a steroid-like compound, is the main component of Ginseng or Panax notoginseng saponin (PNS). Ginseng was shown to exert beneficial effects for amnesia induced by Aβ peptides in vivo. PNS is extracted from Panax notoginseng and has the effect of inhibiting platelet aggregation, increasing heart and cerebral blood flow as well as promoting angiogenesis in dentate gyrus areas of hippocampus. It also plays a protective role in cardiovascular and cerebrovascular diseases [63–65]. A bunch of clinical trials have demonstrated the efficacy of PNS in the treatment of hypertensive intracerebral hemorrhage and ischemic stroke (ClinicalTrials.gov identifier: NCT02999048, NCT01636154, NCT02544087). Gin itself also promotes learning and memory capability and reduces Aβ level of AD mice [66], as well as exerts a neuroprotective effect [67]. In this study, we demonstrated that Gin, rather than Gen, plays a main role in BACE1 modulation, while the exact mechanism remains to be explored in the future. Since reduced glucose with inhibited energy elevates the levels and activity of BACE1, and is potentially amyloidogenic in AβPP transgenic mice, and the energy deficits possibly also an early event in AD pathogenesis [68–70], the capability of Gin to enhance glucose uptake could be critical for its role in modulating BACE1 level and activity. It is worth to note that with a higher concentration, Gin showed effect on the modulation of γ-secretase expression. However, this effect was absent when cells were treated with T2 (Table 1).
A summary of anti-stroke CHMs affect AβPP processing and Aβ clearance
↑ or ↓, Significant change, compared with control; –, No significant change, compared with control.
CONCLUSION
In conclusion, our study suggests that all four CHMs used to treat stroke in the clinic show different effects and mechanisms on AβPP processing, among which T2 with relatively simple components could be a promising candidate for the treatment of AD.
Footnotes
ACKNOWLEDGMENTS
We are grateful for Dr. Louis Lei Jin of International Institute of Systems Medicine, Inc. (Milwaukee, WI) for the manuscript revision.
This research was supported by National Natural Science Foundation of China Project (Project No. 81973690, 81904049), Young Elite Scientists Sponsorship Program by CAST (Project No. 385CACM-2018-QNRC2-C06), Program for Innovative Research Team (in Science and Technology) in University of Henan Province (Project No. 21IRTSTHN026), and Leading Talents Program of Zhongyuan Science and Technology Innovation (Project No. 204200510022).