
Abstract
Alzheimer’s disease (AD) is a central neurodegenerative disease generally among the elderly; it accounts for approximately 50–75%of total cases of dementia patients and poses a serious threat to physical and mental health. Currently available treatments for AD mainly relieves its symptoms, and effective therapy is urgently needed. Deposition of amyloid-β protein in the brain is an early and invariant neuropathological feature of AD. Currently the main efforts in developing anti-AD drugs focus on anti-amyloidogenic therapeutics that prevent amyloid-β production or aggregation and decrease the occurrence of neurotoxic events. The results of an increasing number of studies suggest that natural extracts and phytochemicals have a positive impact on brain aging. Flavonoids belong to the broad group of polyphenols and recent data indicate a favorable effect of flavonoids on brain aging. In this review, we collect relevant discoveries from 1999 to 2021, discuss 75 flavonoids that effectively influence AD pathogenesis, and summarize their functional mechanisms in detail. The data we have reviewed show that, these flavonoids belong to various subclasses, including flavone, flavanone, biflavone, etc. Our results provide a reference for further study of the effects of flavonoids on AD and the progress of anti-AD therapy.
INTRODUCTION
Alzheimer’s disease (AD), the most common type of dementia among elderly people, can seriously affect a person’s ability to perform daily activities [1]. The pathogenesis of AD is complex, and its pathogenic factors have not been accurately determined yet. It involves a variety of cellular changes, including mitochondrial damage, the formation and accumulation of amyloid-β (Aβ), the formation and accumulation of hyperphosphorylated tau protein, synaptic damage, and neuron loss, etc. Among them, Aβ is widely believed to be the origin of AD. AD begins many years before the onset of symptoms. Amyloid-β protein precursor (AβPP) hydrolyzes to produce Aβ [2], then microscopic Aβ protein fragments start to form and slowly grow as chains of fibrils and form pleated sheets and plaques [3].
Aβ plaques evolve into AD through a variety of mechanisms, including the formation of reactive oxygen species (ROS) and stimulation of astrocytes and microglia [4]. As these accumulate, the plaques disrupt cell membranes and cause the death of the brain cells, resulting in neuron damage and memory and cognitive impairment.
Therefore, the study of Aβ clearance mechanism is critical to prevent the progression of AD disease. A large percentage of current therapeutic development efforts for AD focuses on therapies that target the Aβ peptide or Aβ aggregates [5]. Currently, the known clearance mechanisms include ubiquitin-proteasome system, autolysosome, protease, microglia phagocytosis, and the blood-brain barrier (BBB).
The majority of drugs currently on the market are compounds derived from natural product [6]. The role of natural products in the treatment of AD has already been summarized in the literature [7, 8]. Flavonoids are phytochemicals with a wide range of potential therapeutic activities, including anti-inflammatory, antioxidant, and anti-neurodegenerative activities, and the structures and therapeutic activities of flavonoids along with other secondary metabolites present in plants of the genus Iris, have been reviewed by our laboratory [9, 10]. In our research on the chemical constituents of the genus Iris, we reconstructed and modified the structures of the compounds isolated by our laboratory and acquired more than 70 flavonoids, including a new flavanone which has the effects of improving cognitive dysfunction and accelerating Aβ aggregation into senile plaques with lower toxicity in APP/PS1 mice [11]. Recent data from epidemiological studies and animal experiments indicated that flavonoid-rich diets may positively influence brain aging and decrease the incidence of neurodegenerative disorders such as AD and PD [12].
Based on these facts, we searched flavonoids that have been studied deeply at present and screened and summarized the research progress of the corresponding anti-AD mechanism and effect. Here we review how flavonoids act on anti-AD through three main strategies: 1) reduction of Aβ production, which has mainly been approached with secretase inhibition, 2) inhibition of Aβ aggregation and accumulation, and 3) protection against Aβ-induced neurotoxicity. The chemical structures and functional mechanisms associated with the relevant flavonoids are summarized in this article. General information on some natural flavonoids is presented in Table 1.
Natural flavonoids used in the treatment of Alzheimer’s disease
THE ANTI-AD MECHANISMS OF FLAVONOIDS
Reduction of Aβ production
Aβ is produced by sequential proteolytic cleavage of AβPP, a type of integral transmembrane protein consisting of 695–770 amino acids. The cleavage of AβPP takes place via amyloidogenic and non-amyloidogenic pathways. In the amyloidogenic pathway, Aβ is generated in a two-step proteolytic process initiated by β-secretase/BACE1, followed by γ-secretase. This cleavage process produces various Aβ peptides, including Aβ140 and Aβ142 [64]. Hydrophobic Aβ peptides spontaneously polymerize into neurotoxic oligomers in hydrophilic medium, and ultimately form plaques, the hallmark of AD. AβPP can also undergo the proteolytic processing through the non-amyloidogenic pathway. The first enzymatic cleavage in this pathway is mediated by α-cleavage, and occurs within the Aβ sequence domain, thereby precluding the generation of the Aβ peptide [65]. Since Aβ is generated from AβPP via sequential cleavage by β- and γ-secretases, and generation can be precluded by the pathway initiated by α-cleavage, so the secretases have become targets for the control of Aβ production and prevention of the progression of AD.
The vast majority of the reported inhibitors are plant-derived flavonoids, including flavones, and related phenolic compounds. Many of these natural products are non-competitive β-secretase (BACE1) inhibitors that show activity in FRET assays, with few demonstrating potency in cell or animal models [8]. It has been suggested that these compounds might bind to either the Aβ-secretase subsite or the regulatory domain [66], but more detailed mechanism work is needed to clarify these structure-activity relationship issues.
Myricetin (1; its structure is shown in Fig. 1) is a flavonoid found in various plants and wine. It was first separated from the bark of Myrica nagi Thunb 200 years ago [14]. Its oxidation product, dihydromyricetin (2), is a dihydroflavonol that is commonly found in plants, such as bayberry and grapes. Both of these compounds have been intensively studied and have shown great value for people’s health. They show promise in therapies for cancer and inflammatory disorders. Moreover, current research demonstrates their anti-AD effects. Myricetin protects neurons against Aβ-induced toxicity at various stages. It can significantly decrease Aβ1–40 and Aβ1–42 levels in culture medium, an effect that is based on two mechanisms: the activation and upregulation of α-secretase protein levels as indicated by fluorescence resonance energy transfer assay and immune blot analysis, and the direct binding and inhibition of β-secretase (BACE-1), as indicated by cell-free FRET assays [67]. BACE-1 inhibition was verified through a cell-free BACE-1 enzyme activity assay, and the results demonstrated that myricetin was a potent BACE inhibitor (IC50 = 2.8±0.3μM). In silico docking simulation, showed that myricetin forms two hydrogen bonds with Asp32 of BACE through the C3 OH of the C ring, two hydrogen bonds between C4′ and C5′ OH of the B ring and Trp198 and hydrogen bonds with Gln73 through the C7 OH of the A ring; thus, it interacts, directly with the BACE-1 catalytic core through these hydrogen bonds [68]. Myricetin and dihydromyricetin can inhibit the formation of β-sheet. Secondary structure analysis showed that the interaction between myricetin and Aβ inhibits the β-sheets formation of Aβ, which can prevent Aβ undergoing toxic changes. Myricetin can also bind with two additional sites in Aβ fibrils and inhibit the extension of Aβ [69]. Dihydromyricetin can combine with the three sites on the Aβ structure mainly by nonpolar interactions to block its molecular conformation and break its intramolecular hydrogen bonds. This not only blocks the β-sheet but also has a dismantling effect on the already formed Aβ fibrils in PC12 cells [70]. Transgenic mouse models of AD (TG2576 and TG-SwDI) treated chronically with dihydromyricetin 2 mg/Kg/day during 3 months decreases the accumulation of Aβ1–40 and Aβ1–42 [71]. Thus, myricetin and dihydromyricetin can be used in AD treatment.
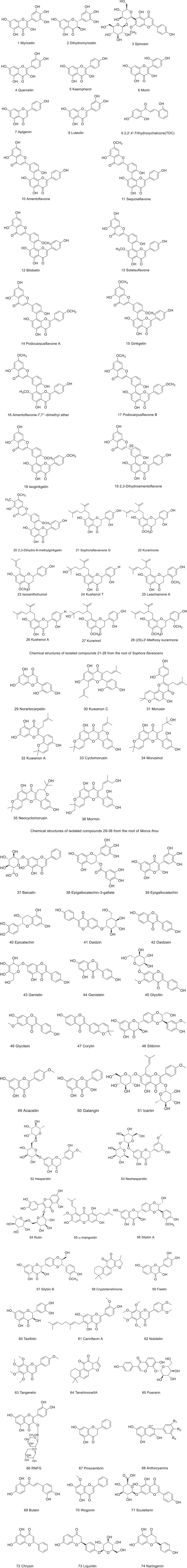
The chemical structure of natural flavonoids.
Spinosin (3), the major active C-glycoside flavonoid in Semen Ziziphi Spinosae (SZS), has been reported to be effective in the treatment of AD. Wild type mouse Neuro-2a cells (N2a/WT) and N2a cells stably expressing human APP695 (N2a/APP695) cells were treated with spinosin for 24 h. The levels of AβPP protein and secreted enzymes closely related to AβPP progression were examined by western blot analysis. The results showed that ROS induced the expression of ADAM10 and reduced the expression of BACE1, while spinosin inhibited ROS production by activating nuclear factor-erythroid 2-related factor 2 (Nrf2) and up-regulating the expression of heme oxygenase-1 (HO-1), which are oxidative stress related proteins. Additionally, spinosin reduced Aβ142 production by affecting the progression of AβPP. In conclusion, spinosin reduced Aβ142 production by activating the Nrf2/HO-1 pathway in N2a/WT and N2a/APP695 cells [16].
Three other flavonols, quercetin (4), kaempherol (5), and morin (6), and one flavone, apigenin (7), were studied in cell-free, cell-based and in silico studies to identify pharmacophore features for BACE-1 inhibition. The cell-free BACE-1 enzyme activity assay demonstrated that these flavonoids directly inhibit BACE-1 enzyme activity in a concentration- dependent manner: quercetin (IC50 = 5.4±0.5μM), kaempherol (IC50 = 14.7±1.3μM), morin (IC50 = 21.7±1.1μM), and apigenin (IC50 = 38.5±2.6μM). Like myricetin, they interact directly with the BACE-1 catalytic core through hydrogen bonds. Compared with myricetin, quercetin forms only one hydrogen bond with Asp32; kaempherol forms two hydrogen bonds through Trp198 and Asp32, morin interacts with Asp228 through a hydrogen bond with C2’OH of the B ring, and apigenin forms one hydrogen bond with Gln73. These results suggest that, in many cases, the BACE-1 inhibitory activity of flavonols is enhanced by the OH of C3 with the Asp catalytic residue [68].
Morin (2,3,4,5,7-pentahydroxyflavone) (6) is a natural bioflavonoid that is abundantly found in fruits, vegetables, green tea, red wine, and many oriental medicinal herbs [72]. Many studies have shown that morin exerts potent neuroprotectant activity via its anti-oxidant, anti-apoptotic, and anti-inflammatory properties in vitro and in vivo. The disease-modifying effects of chronic morin administration on the neuropathological and cognitive impairments found in APPswe/PS1dE9 double transgenic mice have been investigated. The results showed that chronic morin administration prevented spatial learning and memory deficits in APPswe/PS1dE9 mice. Morin treatment of APPswe/ PS1dE9 mice markedly reduced cerebral Aβ production and Aβ plaque burden by promoting the non-amyloidogenic AβPP processing pathway through increased ADAM10 expression, inhibiting the amyloidogenic AβPP processing pathway through decreased BACE1 and PS1 expression, and facilitating Aβ degradation through enhancement of the expression of Aβ-degrading enzyme [74].
Apigenin (4’, 5, 7-trihydroxyflavone) (7) is a flavonoid rich in the ligulate flowers of the chamomile plant [75]. It can serve as a pharmacologically active agent and is considered to have no toxicity even at high doses. A study by Zhao et al. examined the effects of apigenin on cognitive function in APP/PS1 double transgenic AD mice and explored its mechanism(s) of action. Oral administration of apigenin to APP/PS1 mice for three-month rescued learning deficits and relieved memory retention. Apigenin also affected AβPP processing and prevented the Aβ burden due to the downregulation of BACE1 and β-CTF levels, causing relief of Aβ deposition, and a decrease in insoluble Aβ levels.
Luteolin (8), a Citrus bioflavonoid, was isolated from a methanolic extract of Perilla frutescens var. acuta, and was shown to have BACE1 inhibitory activity with an IC50 value of 5.0×10-7 M. It inhibited BACE1 in a non-competitive manner with substrate as shown by Dixon plots and was a relatively specific inhibitor of BACE1 [66]. Another study conducted by Kavon Rezai-Zadeh demonstrated that luteolin exerts its anti-amyiloidogenic effects through down-regulation of γ-secretase activity. Correlated with the decreases in total Aβ generation, γ-secretase cleavage activity was decreased by luteolin in a concentration- and time-dependent manner. The specific mechanism through which luteolin modulates γ-secretase activity and subsequent Aβ generation is suggested that luteolin reduces GSK-3α/β activation in SweAPPN2a cells and Tg2576 mouse-derived primary neuronal cells [76]. In another study, luteolin was identified as a potent inhibitor of BACE1 transcription in human embryonic kidney 293 (HEK293) and human neuroblastoma (SH-SY5Y) cell lines. Luteolin can suppress activation of the BACE1 promoter by NF-B signaling. The cited study further showed that luteolin interferes with NF-B signaling by both directly and indirectly disrupting p65 complex formation. In addition, the investigators discovered that the estrogen receptor mediates luteolin’s inhibitory effects on NF-B signaling and BACE1 transcription. Interestingly, the beneficial effects of luteolin may be related to the selective activation profiles of luteolin to different estrogen receptor subtypes. Their study reports luteolin as a potent BACE1-inhibiting compound, providing useful information in understanding estrogen receptor- and NF-B-mediated signaling in regulating BACE1 expression [24].
The natural product 2, 2’, 4’-trihydroxychalcone (TDC) (9) from Glycyrrhiza glabra inhibits BACE1 with an IC50 of 2.45μM and was shown to be a specific non-competitive inhibitor of the enzyme. TDC exhibited no effects on either α- or γ-secretase activity, and an in vivo assay showed that TDC inhibits BACE1 activity, thereby reducing Aβ production in Tg APP-PS1 mice [27]. The above results suggest that TDC may inhibit AβPP cleavage and thus reduce Aβ production in mice by functioning as a specific BACE1 inhibitor.
Biflavonoids are flavonoid dimers connected by C-C or C-O-C bonds and are well known as constituents of gymnospermous plants. The structure-activity relationships of a variety of biflavonoids isolated from different plants were studied, amentoflavone (10) and its monomethoxy analogues (11–14) showed strong inhibitory activity against BACE-1, with IC50 values of 1.54, 1.40, 2.02, 1.58, and 0.99μM, respectively. Compounds 15–18 showed lower activities than compounds 10–14 with IC50 values of 4.18, 6.25, 4.21, and 3.01μM, respectively. Compared with other amentoflavone-type biflavonoids, compound 19 (IC50 = 0.75 μM) and compound 20 (IC50 = 0.35 μM) showed stronger inhibition of BACE-1. These results indicate that the presence of two apigenin molecules linked at the C3’- C8” position is important for the BACE-1 inhibitory activity of amentoflavone-type biflavonoids and that more than two hydroxyl groups at the R1-R4 position are needed for inhibitory activity. The results obtained for 2,3-dihydroamentoflavone (19) and 2,3-dihydro-6-methylginkgetin (20) show that the presence of a flavanone moiety in the amentoflavone biflavonoid is advantageous for inhibitory activity. Moreover, the presence of a methyl group at the C6 position increases the inhibitory effect [77].
Lavanduluylated flavanones achieved excellent BACE-1 inhibitory activity both in vitro and in a cell-based assay. Lipophilic alkylated (C10-C5) flavanones from Sophora flavescens were examined for their inhibitory effects against BACE-1. Lavandulyl flavanones 21, 22, 25, 26, and 28 showed potent BACE-1 inhibitory activities with IC50 values of 5.2, 3.3, 8.4, 2.6, and 6.7μM, respectively, while no significant activity was observed for the corresponding hydrated lavandulyl flavanones 24 and 27 or for prenylated flavanone 23. In transfected human embryonic kidney (HEK-293) cells, lavandulyl flavanones reduced Aβ secretion dose-dependently and in kinetic studies, all compounds screened were shown to be noncompetitive inhibitors [78]. Moreover, a study of the structure-activity relationships of prenylated flavonoids from S. flavescens showed that the prenyl group at the C-8 position plays a predominant role in BACE1 inhibition. In addition, prenylated flavonols and prenylated chalcones exhibited potent BACE1 inhibition, indicating that BACE1 inhibition may be attributed to specific C-ring types of flavonoid skeletons [79]. In summary, the results indicate that the prenyl group, rather than the lavandulyl group, and flavonols and chalcones, rather than flavanones, might make predominant contributions to BACE1 inhibition.
Eight flavones, norartocarpetin (29), kuwanon C (30), morusin (31), kuwanon A (32), cyclomorusin (33), morusinol (34), neocyclomorusin (35), and mormin (36), were isolated from the stem bark of Morus lhou. They showed dose-dependent, reversible inhibition of BACE-1 activity. Among them, kuwanon C (IC50 = 3.4μM) showed the strongest inhibitory activity against BACE-1 and was 20 times more potent than the corresponding parent compound norartocarpetin (IC50 = 60.6μM). BACE1 inhibitory potency was significantly increased in the presence of two free hydroxyl groups in the resorcinol moiety (B-ring) and isoprenyl functionality at C3 in ring A of the flavone. Kuwanon C (IC50 = 3.4μM) and kuwanon A (IC50 = 5.3μM) in which both hydroxyl groups in the resorcinol ring (ring B) are unsubstituted, exhibited significant activity. Compounds bearing alkyl substitutions therein (31, 33–36) showed lower activities (IC50 values of 59.4, 101.2, 135.9, 146.1, and 103.5μM, respectively), suggesting a crucial role of the free hydroxylgroups of the resorcinol ring of flavones in BACE inhibitory activity. Furthermore, ring-closure of the prenyl group onto C7-OH to form pentacyclic structures also led to lower potency, indicating that a free C7-hydroxyl is essential for the activity and/or that rings A and C adopt a conformation in the active site, which is not achieved in pentacyclic analogues. In summary, the best inhibitors possessed both a free resorcinol group and at least one prenyl motif at C3 in the A-ring. The OH group of the resorcinol moiety of the B-ring interacted with Thr 231, Thr 329 and Asp 228 of the enzyme by forming hydrogen bonds. These two prenyl groups established hydrophobic interactions with the surrounding amino acid residues Gly 34, Ser35, Val 69, Pro 70, Tyr 71, Ile 126, Tyr 198, Lys 224, and Thr 320 [80].
Baicalin (37) is a bioactive flavone with anti-inflammatory and neuroprotective activities. The neuroprotective effects of baicalin on pathological changes and behavioral deficits were explored in a mouse model of Aβ1–42 protein-induced AD. Mice received a bilateral injection of Aβ1–42 protein into the hippocampus, and were then treated with baicalin (30, 50, or 100 mg/kg body weight, orally) or Tween 80. The therapeutic effects of baicalin were monitored using the Morris water maze test and the probe test. Treatment of the mice with 100 mg/kg baicalin for 14 days significantly ameliorated memory impairment and glial cell activation and increased tumor necrosis factor (TNF)-α and interleukin (IL)-6 expression induced by the Aβ1–42 protein. These results suggest that baicalin ameliorates Aβ1–42 protein-related pathology and cognitive dysfunction via its anti-neuroinflammatory activity and that it may be a potential candidate for the treatment of AD [23].
Catechins are a class of phenolic active substances extracted from natural plants such as tea. Epigallocatechin-3-gallate (EGCG, 38), a major bioactive catechin has been found to have promising neuroprotective effects. It can significantly regulate AβPP processing, and decrease brain Aβ production and plaque burden by enhancing the synthesis of α-secretase (ADAM10) in APPswe/PS1dE9 mice [81]. It also possesses anti-inflammatory properties. Pre-administration of EGCG (1.5 and 3 mg/kg for 3 weeks) prevented lipopolysaccharide-induced memory impairment in ICR mice and suppressed the increase in cytokines and inflammatory proteins seen in untreated controls [82]. Epigallocatechin (39) and (-)-epicatechin (40) also belong to the catechin family, and studies have demonstrated that oral administration of (-)-epicatechin to 7-month-old TASTPM mice for 21 days can reduce Aβ pathology and Aβ levels. Furthermore, in vitro mechanistic studies suggest that this is likely due to indirect inhibition of BACE1 [83]. Taken together, the results suggest that orally delivered catechin may be a potential prophylactic agent for AD.
Soy isoflavones, including daidzin (41), daidzein (42), genistin (43), genistein (44), glycitin (45), and glycitein (46) [84, 85], have attracted much interest in recent years due to their estrogen-like effects and their role in influencing sex hormone metabolism. Estrogen exerts anti-AD effects through several mechanisms such as reducing Aβ production [86], antagonizing the toxicities of Aβ [87], and promoting synaptic growth and the expression of nerve growth factor and its receptor [88]. Mie Hirohata’s three-dimensional fluorescence spectroscopic analyses demonstrated that glycitein interacted with Aβ monomers, oligomers and fibrils in vitro using a nucleation-dependent polymerization model, indicating specific binding of glycitein to these Aβ species. Glycitein also interacted with various Aβ fragments (Aβ1–42, Aβ1–40, Aβ1–16, and Aβ25–35), exhibiting the most potent affinity for Aβ25–35. They speculated that glycitein’s anti-amyloidogenic properties are specifically mediated by its binding to Aβ monomers, oligomers and fibrils [63]. Genistein is a promising agent for the treatment of AD pathogenesis. A recent study showed that administration of genistein enhanced Aβ-induced cognitive impairment by attenuating synaptotoxicity, hyperphosphorylation of tau, and inactivation of extracellular signal regulated protein kinase (ERK) in Aβ-infused rats [89]. Due to their low systemic toxicity, soy isoflavones show potential treatment promise for AD therapy.
Corylin (47), an isoflavone extracted from Cullen corylifolium (L.) Medik., has been widely used in Asian countries for its anti-inflammatory and anticancer activities [90]. Recent studies have shown that corylin exerts a protective effect on the expression of Aβ in PC12 cells. At 10.1μmol/L corylin can significantly reduce the expression of AβPP, BACE-1, and Aβ [91].
Silymarin, the major component of which is silibinin (48), an extract of milk thistle, is frequently used as a medicinal herb in the treatment of liver diseases. However, recent studies have demonstrated that it shows promise in the treatment of AD. The effect of silymarin on passive avoidance learning was examined in a rat model of AD that was induced by intracerebroventricular injection of Aβ peptide (Aβ1–42) into Wistar rats. The rats were treated with 70 or 140 mg/kg of the milk thistle extract, once a day, for 4 weeks. Memory function improved and the number of brain amyloid plaques also decreased upon administration of the extract. Furthermore, comparison of APP gene expression in the treated and untreated groups showed that silymarin suppressed AβPP expression [92].
Acacetin (49) extracted from Agastache rugosa has anti-BACE-1 and behavioral activities in Drosophila melanogaster AD models, and its functional mechanism has been elucidated. Acacetin (100, 300, and 500μM) rescued AβPP/BACE1-expressing flies and kept them from developing both pathological eye morphological changes (dark deposits, ommatidial collapse and fusion, and the absence of ommatidial bristles) and behavioral (motor abnormalities) defects. Reverse transcription polymerase chain reaction analysis revealed that acacetin reduced both human AβPP and BACE-1 mRNA levels in the transgenic flies, suggesting that it plays an important role in the transcriptional regulation of human BACE-1 and AβPP. Western blot analysis revealed that acacetin reduced Aβ production by interfering with BACE-1 activity and AβPP synthesis, resulting in a decrease in the levels of the APP carboxyterminal fragments and the AβPP intracellular domain. In summary, acacetin exerts its effects on inhibiting Aβ production by transcriptional regulation of AβPP and BACE-1 and AβPP synthesis, resulting in a decrease in the number of amyloid plaques [32].
Galangin (50), isolated from the rhizome of Alpine officinarum Hance, has a significant lowering effect on β-secretase, Aβ42, and p-tau levels. Investigation has shown that epigenetic mechanisms, such as histone acetylation and DNA methylation, are involved in the galangin-induced transcriptional regulation of BACE1. The data showed that galangin induced a decrease in acetylated H3 in the BACE1 promoter regions through the upregulation of endogenous HDAC1-mediated deacetylation, a modification that is independent of DNA methylation status. It can also protect PC12 cells from okadaic acid-induced cytotoxicity and inhibit autophagy via the Akt/GSK3β/mTOR pathway [33]. The above findings suggest a novel mechanism for the neuroprotective effect of polyphenols on neurodegeneration and suggest that galangin may be a potential drug candidate for AD therapy [93].
Icariin (51), a prenylated flavonol glycoside derived from the traditional Chinese herb Epimedium brevicornum, offers protection against AD. A number of studies indicate that icariin decreases the expression of both AβPP and BACE1 and thereby decreases the expression of cytotoxic Aβ142 in AD-treated mice. Some previous studies have reported that icariin enhances spatial learning and memory in various models, including ten-month-old APP/presenilin1 (PS1) transgenic mice over-expressing human APP (APP695swe) and PS1 (PS1-dE9) [94], APP transgenic (Tg) mice [95], Tg2576 mice [34], and triple-transgenic (3×Tg) mice. A potential mechanistic investigation demonstrated that icariin is effective in improving cognitive deficits and hippocampal morphological alterations caused by bilateral common carotid arteries. Inhibition of TGF-β1 signaling and Smad2/3 phosphorylation are also involved in the course [96].
Hesperidin (52), a flavonoid that is particularly abundant in oranges and lemons, exerts anticarcinogenic, antihypertensive, antiviral, antioxidant and anti-inflammatory effects [36]. Li and coworkers found that oral administration of hesperidin (100 mg/kg body weight) to mice for 10 days repaired deficits in non-cognitive nesting capability and social interaction and that it attenuated Aβ deposition, plaque associated AβPP expression, microglial activation, and TGF-β1 immunoreactivity in both the cerebral cortex and the hippocampus of APP/PS1 mice [37]. A recent study also found that hesperidin can antagonize the memory loss caused by aluminum. It exerts this effect by lowering acetylcholine esterase activity and the expression of Aβ biosynthesis related markers [97]. Hesperetin, which is produced from hesperidin, has also been shown to be valid in AD treatments. Studies have shown that, the expression of AβPP, BACE-1, and Aβ in Aβ142–treated mice can be moderated by administration of hesperetin. Moreover, hesperetin-treated mice significantly enhanced memory function compared with untreated mice [98].
A natural flavanone glycoside, neohesperidin (53), commonly found in citrus food, has been reported to dually inhibit BACE1 and Aβ aggregation. Multi-spectroscopic approaches, force microscopy, and molecular modelling were used to validate its potency in SH-SY5Y neuroblastoma cell lines. Steady-state and time-resolved fluorescence revealed that neohesperidin binds close to the catalytic aspartate dyad. This binding conformationally restricts the protein in closed form, which possibly precludes AβPP recognition and thereby inhibits BACE1 activity. Neohesperidin also dose-dependently inhibits amyloid fibril formation, as is evident from ANS, ThT assay, and AFM. Neohesperidin ameliorates aggregated Aβ25–35 induced ROS generation and mitochondrial dysfunction in the SH-SY5Y cell line. The amyloid induced apoptosis was significantly inhibited, and normal neuronal morphology was rescued. These findings suggest that neohesperidin is an inhibitor of the pathogenic conversion of Aβ to fibrillar amyloid assembly [18].
Inhibition of Aβ aggregation and accumulation
Great efforts have been made to identify a growing number of natural flavonoids that block the enzymatic activity involved in the process of Aβ production. In fact, the activity of both enzymes was proven to be essential for neuronal functioning. Lack of γ-secretase activity can even trigger the onset of AD [57]. To date, contributions have been made to discover drugs inhibiting Aβ aggregation.
Aβ peptides self-assemble into neurotoxic intermediates, such as soluble oligomers and protofibrils, and this is considered another key event in the pathogenesis of AD. All of these Aβ species may differentially affect neuronal function and viability in vitro and in vivo [99, 100]. Aβ fibrils induce neurodegenerative changes, including neuronal cell death, apoptosis, oxidative stress, and calcium deregulation [101]. Accumulating evidence indicates that β-sheet formation triggers more neurotoxicity [102]. That is, spherical intermediate and small-sized soluble Aβ oligomers that are formed during peptide aggregation are cytotoxic while mature fibrils are inert. Therefore, prevention of Aβ aggregation and/or decomposition of existing oligomers and fibrils by small molecule compounds are promising strategies for the prevention and treatment of AD. Many compounds have been found to reduce Aβ aggregation or neurotoxicity in vitro; these include oligopeptides [103], polyphenols [104], antibodies [105], antioxidants [106], and non-steroidal anti-inflammatory drugs [107]. Here, we review the anti-Aβ aggregation effects of flavonoids from natural plants.
Epigallocatechin-3-gallate (EGCG,38) and quercetin (4) both of which were mentioned above have been demonstrated to interfere with Aβ42 aggregation and neurotoxicity [108, 109]. EGCG suppressed fibril elongation and changed the fibril structure in such a way that it displayed less seeding activity in secondary nucleation in vitro [110]. Studies of the mechanism of action of EGCG show that it efficiently inhibits fibrillogenesis of Aβ by directly binding to natively unfolded polypeptides and preventing their conversion into toxic amyloid oligomers and protofibrils in PC12 cells. Instead of β-sheet-rich amyloid, nontoxic and highly stable off-pathway, EGCG-containing spherical oligomers are formed, leading to lower Aβ42 production and a decreased aggregation rate. EGCG is a generic modulator of amyloid formation that binds to unfolded polypeptide chains independently of the primary amino acid sequence and converts them into nontoxic, unstructured, off-pathway oligomers of a new type [111]. Accumulating studies have proven that EGCG reduces the thioflavin T fluorescence of Aβ42 aggregates in a concentration-dependent manner. A recent study in vitro demonstrated that EGCG affects the amount of Aβ42 fibrils in two ways. While low concentrations of EGCG delayed the formation of Aβ42 fibrils, high concentrations of EGCG promoted fibril formation [112]. These findings may provide a new strategy for the treatment or prevention of AD.
Quercetin (4) exhibits properties that decrease fibril formation, including inhibition of Aβ aggregation and cytotoxicity [108, 113]. Quercetin and rutin (54) inhibited the formation of Aβ fibrils and disaggregated Aβ fibrils and they appeared to exert anti-amyloidogenic effects in vitro by reversibly binding to the amyloid fibril structure of Aβ oligomer and to Aβ monomers [38]. In a comparison of the structures of several structurally related flavonoids in vitro, the 3’,4’-dihydroxyl groups of quercetin were found to be essential for its inhibitory effect on Aβ fibril formation [114]. In vitro aggregation studies have illustrated that quercetin inhibits the formation of Aβ fibrils and destabilizes preformed mature fibrils by reinforcing the hydrophobic interaction between the aromatic rings and the β-sheet structures of Aβ by hydrogen bonds. In addition, the 3’,4’-dihydroxyl group of the B ring plays a key role in its anti-aggregation effect [38]. Quercetin inhibits Aβ fibril formation and destabilizes mature fibrils at IC50 concentrations of 0.72 and 0.73 mM [113].
Morin (6), which is widely distributed in plants and foods of plant origin, has been shown to suppress Aβ aggregation. In atomistic, explicit-solvent molecular dynamics simulations, morin was shown to inhibit the early stages of Aβ peptide aggregation by altering tertiary and quaternary interactions to produce “off-pathway” structures even at low concentrations. However, morin does not affect the secondary structure of Aβ peptides, except in the presence of a large molar excess of morin. The regions on the peptide that gave rise to tertiary and quaternary structural changes are the CHC and polar bend region, thus the main effect that morin has on the structure of Aβ40 and Aβ42 in the monomeric and dimeric states is to alter tertiary and quaternary interactions. These results provide molecular insight into the nature of so-called “off-pathway” aggregates that may have reduced toxicity, thus triggering the course of overcoming principal toxic entities associated with AD [19].
α-Mangostin (α-M) (55), a polyphenolic xanthone derivative of mangosteen (Garcinia mangostana L.), has been reported to be a potential multi-factorially acting anti-AD agent with a certain hepatoprotective effect. Molecular docking and dynamics simulations demonstrated that α-M can potentially bind to Aβ and stabilize its α-helical conformation. α-M was found to directly dissociate Aβ140 and Aβ142 oligomers by blotting with oligomer-specific antibodies. ThioflavinT fluorescence assay and electron microscopy imaging further demonstrated that α-M blocks the fibril formation and disturbs the preformed fibrils in primary rat cerebral cortical neurons [115]. According to WHO recommendations, α-M is safe for human consumption, and shows promise in anti-AD therapy.
A cell level study showed that the phenolic compound myricetin (1) could block early Aβ assembly processes and its toxicity. Myricetin binds to the localized or specific monomeric Aβ region Arg-5, promotes NMR chemical shift changes and lowers cellular toxicity and synaptic dysfunction caused by the Aβ oligomers [116]. Another study showed that myricetin has a strong affinity for native and partially folded protein conformers that is mediated by H-bonds and hydrophobic interactions and that it is capable of promoting the disassembly of amyloid fibrils. Discoveries in vitro have revealed that the myricetin molecule has the potential to suppress amyloid formation and this inherent property may contribute to the development of myricetin-based anti-amyloid drugs [117].
Silibinin (48), a flavonoid derived from the herb milk thistle (Silybum marianum), is a diastereoisomeric mixture of two flavonolignans, silybin A (Sil A, 56) and silybin B (Sil B, 57) in a ratio of approximately 1:1. Silibinin is a candidate for attenuating cognitive deficits induced by the Aβ2535 peptide, and its inhibitory effect on the formation of Aβ is mentioned above. Based on ThT assays and electron microscopic imaging, silibinin appears to inhibit Aβ aggregation dose-dependently [118]. Low amounts (4μM) of Sil B entirely abolished amyloid aggregation, and further toxicity assays revealed that Sil B significantly protects transgenic C. elegans from the toxicity induced by Aβ, proving that it can interact with toxic oligomers in vivo [119]. The result suggests that Sil B is a promising inhibitor of fibril and oligomer production.
Cryptotanshinone (CTS) (58), an abietane diterpenoid extracted from the medicinal herb Salvia miltiorrhiza, has been shown to improve learning and memory in several pharmacological models of AD [120, 121], and its effects on Aβ aggregation and toxicity have been studied. In thioflavin T fluorescence assays and transmission electron microscopy studies of SH-SY5Y cells, CTS inhibited Aβ aggregation. Researchers have suggested that the most likely molecular mechanism through which CTS can inhibit Aβ self-assembly, is that the dynamics of the process in which soluble Aβ undergoes a conformational change to a high β-sheet content, rendering it prone to aggregation into soluble oligomers and larger insoluble high-molecular-weight assemblies are affected by CTS treatment [122].
Fisetin (3, 3’, 4’, 7-tetrahydroxyflavone) (59) has been found to be neuroprotective, and to induce neuronal differentiation, enhance memory, and inhibit the aggregation of the Aβ protein in mice [123, 124]. The structure-activity relationships of fisetin were studied, and the results showed that the 3’, 4’-dihydroxyl group of the B ring is essential for the inhibitory effect on Aβ fibril formation, while the 3-hydroxyl group or 7-hydroxyl group is not necessary. Moreover, hydroxylation at the 5-position may partly contribute to the inhibition of Aβ fibril formation [114].
Taxifolin (60), a flavonol with strong anti-oxidative and anti-glycation activities, is present in various plants, such as herbs, and has been shown to inhibit Aβ fibril formation in vitro and to improve cerebral blood flow, facilitating Aβ clearance in the brain and suppressing cognitive decline in a mouse model of cerebral amyloid angiopathy [43]. The functioning mechanism is related to the chemical structure, 3’,4’-dihydroxyl groups on the B-ring. Its catechol structure can transfer into an oxidized form and react with Aβ42, causing the production of Aβ42–taxifolin adducts. The taxifolin targeting site of Aβ42 is located in the intermolecular β-sheet region and contributes to the inhibition and destabilization of Aβ42 aggregation, and to suppression of the elongation phase rather than the nucleation phase during the process of Aβ42 fibril formation. Based on the results, it is proposed that catechol-type flavonoids can inhibit the aggregation of Aβ42, in which a catechol structure could be autoxidized to form the o-quinone on the B-ring, followed by the formation of the o-quinone-Aβ42 adduct targeting Lys residues at positions 16 and 28 of Aβ42, but not be originated from the antioxidative activity [125].
Luteolin (8) treatment of murine N2a cells transfected with SweAPP and of primary neuronal cells derived from SweAPP overexpressing mice resulted in a significant reduction in Aβ generation. The mechanism appears to involve selective inactivation of the GSK-3α isoform, which in turn increases the phosphorylation of PS1, the catalytic core of the γ- secretase complex, thereby reducing PS1 APP interaction and Aβ generation [76]. Luteolin treatment is effective in suppressing Aβ accumulation induced by traumatic brain injury (TBI), which is regarded as a main risk factor for AD [126]. Luteolin treatment reduces amyloid pathology in Tg2576 mice by blunting the increase in soluble Aβ40,42 levels induced by TBI [127].
The novel geranylated cannabis-derived flavonoid, cannflavin A (61) is one of several flavonoids which have been identified in the Cannabis sativa L. plant. Cannflavin A demonstrated intrinsic hormetic effects on cell viability, increasing viability by 40%at concentrations ranging from 1 to 10μM but displaying neurotoxicity at higher concentrations (> 10–100μM), that is, it exhibited a biphasic effect. 10μM cannflavin A inhibited the neurotoxicity elicited by Aβ1–42 (0–2μM), reducing Aβ aggregate adherence to PC12 cells and associated neurite loss [17].
The study of structure-specific novel protein sumoylation inhibitors, 2′, 3′, 4′-trihydroxyflavone (2-D08) has effective suppressing capacity in PC12 cells and can compensate for the limitations of natural flavonoids nobiletin for the extension of B ring hydroxylation with methoxyl groups, resulting in an incremental loss of anti-fibrillar and neuroprotective activity. The extensive hydroxylation in the B ring of 2-D08 is the most important determinant of activity against Aβ within the flavone scaffold. These findings provide further structural insights into the anti-amyloid bioactivity of flavonoids in addition to demonstrating a novel and efficacious anti-aggregation and neuroprotective effect of semi-synthetic flavones. Such modified flavones may contribute to a better understanding of neurodegenerative disease and promotion of drugs that target multiple pathways [128].
Protection against Aβ-induced neurotoxicity
High Aβ production levels could contribute to neurotoxicity and to the neuronal death observed in AD [129]. Although the precise mechanism is unclear, several possible mechanisms of Aβ-induced neurotoxicity have been proposed: one of them is oxidative stress induced by Aβ. Recent studies have shown that Aβ oligomers and aggregated Aβ can induce oxidative stress which plays an important role in AD pathogenesis [130]. Higher levels of oxidative DNA damage in mitochondria of the frontal, parietal, and temporal lobes suggest that mitochondrial oxidative stress may be an important contributor to the pathogenesis of AD [131]. Transient or sustained mitochondrial dysfunction can deplete ATP levels, increase ROS generation, and initiate apoptosis, leading to neurodegeneration [132]. The other neurotoxic mechanism associated with Aβ is the destabilization of calcium homeostasis. Aβ appears to induce Ca2 + influx into the cytoplasm from extracellular. Ca2 + overloads in the cytoplasm, then triggers neurotoxicity by enhancing the release of glutamate, eventually leading to cytotoxicity [133]. Therefore, inhibition of Aβ-induced neurotoxicity would be an efficient therapeutic strategy for AD.
Nobiletin (62), 3’, 4’, 5, 6, 7, 8-hexamethoxyflavone, is found in the peel of Citrus depressa. It can rescue Aβ-induced memory deterioration and ameliorate oxidative stress in animal models of AD. Nobiletin affects oxidative stress levels in cells such as PC12 cells, HT22 cells, and SK-N-SH cells. It exerts its effect on PC12 cells through hydrogen peroxide (H2O2)-induced apoptosis, and this is associated with inhibition of the H2O2-induced decrease in GSH levels and superoxide dismutase activity [45]. It can also ameliorate cytotoxicity in PC12 cells indirectly by increasing intracellular GSH levels through the upregulation of glutamate-cysteine ligase, a rate-limiting enzyme for GSH synthesis [134]. There are also ways on gene levels. Nobiletin blocks H2O2-induced enhancement of the expression of the pro-apoptotic protein Bax, while expression of the anti-apoptotic protein Bcl-2 is triggered by nobiletin in HT22 cells [44]. Nobiletin exerts its anti-neuroinflammation effect by suppressing the lipopolysaccharide (LPS)-induced production and secretion of proinflammatory mediators, such as nitric oxide (NO), TNF-α, IL-1β, and IL-6, in the mouse microglia BV-2 cell line [135–137]. A growing number of studies have provided a solid basis for the development of AD therapy.
Tangeretin (63) and nobiletin (62) are polymethoxy flavonoids that are abundantly present in the pericarp of Citrus unshiu (family Rutaceae) and in the fruit of Citrus depressa (family Rutaceae). Tangeretin has been shown to have neuroprotective capacity both in vitro and in vivo. It could mitigate cholinergic deficits, decrease the abnormal accumulation of neurotoxic Aβ peptides, reverse N-methyl- D-aspartate (NMDA) receptor hypofunction, ameliorate ischemic injury, inhibit hyperphosphorylation of tau protein, enhance neprilysin levels, regulate several signaling cascades, and protect against 1-methyl-4-phenylpyridinium (MPP(+)) and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) toxicity [138].
Hesperidin (52), a bioactive flavonoid, has antioxidant, anti-inflammatory, and anti-apoptotic actions that reduce Aβ-induced neurodegeneration and memory impairment. Oxidative stress in the hippocampus, cortex, and in vitro HT22 cells was decreased based on the expression of Nrf2/HO-1 on the results of LPO and ROS assays. Activated glial cells were modulated by comparing the expression of GFAP and Iba-1. The expression of TLR4, P-NF-κB, and downstream targets was also analyzed. The results suggested that hesperidin restored the expression of these markers. Hesperetin has also been shown to have strong antiapoptotic effects [98]. The functional mechanism through which it protects against Aβ-induced neurotoxicity has been partly elucidated and shown to involve the mitochondrial voltage-dependent anion channel 1 (VDAC1)-regulated mitochondrial apoptotic pathway in PC12 cells. VDAC1 is involved in the release of apoptotic proteins that may be relevant to AD neuropathology, and it plays an important role in maintaining mitochondrial function and integrity. By reversing Aβ-induced mitochondrial dysfunction, including mitochondrial permeability transition pore opening, increase in intracellular free calcium and reactive oxygen species production, treatment with hesperidin significantly inhibited Aβ2535-induced apoptosis [139].
Tanshinone IIA (64) protected neurons against the neurotoxicity of Aβ2535, elevated the viability rate of neuron cells, decreased the expression of p-tau in neurons induced by Aβ2535, and improved cellular impairment on ultramicrostructure. These protective effects may be due to its inhibition of the cleavage of P35 into P25, Cdk5 translocation from the nuclear to cytoplasm and its hyperphosphorylation activity induced by Aβ2535. This hyperphosphorylation reduced the impairment of Aβ2535 in primary cortical neurons through the Cdk5/P35 pathway [140]. Tanshinone IIA inhibits the apoptosis of nerve cells in the rat model of neonatal hypoxic ischemia brain damage possibly by reducing the expression of Caspase-3 as assessed by Tunel staining, western blot, and other experimental methods [141]. Use of the MTT method, flow cytometry, western blot, and other experimental methods showed that tanshinone IIA improves the survival fraction of cultured HT-22 hippocampal neurons after exposure to high levels of radiation, reduces the apoptosis and enhances autophagy related proteins LC3-II expression. It suggests that tanshinone IIA can induce autophagy and improve radiation injury of stress, so as to exert its protective effect of hippocampal neurons [142].
Puerarin (65) is a phytoestrogen isolated from Pueraria lobata, and several studies have been conducted to assess its neuroprotective effects against neurotoxicity due to Aβ. It was demonstrated that puerarin protects Aβ-induced neurotoxicity in a manner that is related to the mitochondria mediated cell death process. Puerarin afforded protection by inhibiting neuronal apoptosis in PC12 cells [46]. Puerarin could be a potential therapeutic agent for neurodegenerative diseases involving oxidative stress according to a study of neuronal cultures from rat hippocampus [143]. It can attenuate Aβ140-induced NLRP3 inflammasome activation, the driving factor in the pathogenesis of chronic diseases by suppressing ROS-dependent oxidative stress and endoplasmic reticulum stress. Its anti-inflammatory capability is mediated by activation of Nrf2/HO-1 and suppression of the IRE1/PERK/ATF6 signaling pathway [48]. Puerarin protected neurons in the cortex and hippocampus of AD rats against apoptosis caused by Aβ2535 through downregulating Aβ140 and Bax expression in brain tissues, thereby alleviating the spatial learning and memory impairment of diseased animals [47, 144].
The inhibition of quercetin (4) on Aβ fibril formation has been described above. Studies conducted in vitro, in laboratory animals, and in humans have provided supporting evidence for the neuroprotective effects of quercetin, in a variety of models of neuronal damage and neurodegenerative disease. Quercetin almost completely decreased ROS generation in H2O2-treated APPswe cells and increased intracellular GSH content and the redox status. In addition, quercetin diminished the lipid peroxidation index in comparison with control APPswe cells at all concentrations tested [38]. The nanoparticles of quercetin are also a candidate for AD therapy through hindering the damaging effect of AlCl3 on hippocampal neurons [145]. It also protects HT22 murine neuroblastoma cells from Aβ oxidative attack [123]. Apart from its direct antioxidant effects, quercetin antagonizes oxidative stress through a general mechanism of neuroprotection, probably by two pathways: induction of Nrf2-ARE and induction of the antioxidant/anti-inflammatory enzyme paraoxonase 2 [146, 147]. Quercetin has been shown to activate sirtuins and to trigger autophagy. The protective effects of quercetin on Aβ-induced oxidative cell toxicity were investigated in cultured neurons, and pretreatment of primary hippocampal cultures with low doses of quercetin significantly attenuated Aβ-induced cytotoxicity, protein oxidation, lipid peroxidation and apoptosis [108]. Another study demonstrated that quercetin enhanced brain apoE levels and decreased Aβ levels in the cortex of amyloid model mice [148]. MAPT, PIK3R1, CASP8, DAPK1, MAPK1, and CYCS can act as biomarkers for AD diagnosis and as targets of quercetin treatment through bioinformatics analysis and experimental identification [149].
A Radix notoginseng flavonol glycoside (RNFG) (66) was isolated from the roots of Panax notoginseng, and various types of assays showed that it possessed a strong activity in preventing Aβ-induced cell death. RNFG reduced Aβ-induced cell death in a time- and dose-dependent manner with suppression of Aβ-induced DNA fragmentation and caspase-3 activation in cultured cortical neurons [49].
Pinocembrin (67), a flavonoid abundant in propolis, has been demonstrated to have neuroprotective effects on ischemic and vascular dementia in animal models [150]. Further investigations showed that pinocembrin improved cognitive function, preserved ultrastructural neuropil and decreased neurodegeneration of the cerebral cortex in Aβ2535-treated mice by RAGE-dependent transduction inhibition and mitochondrion protection [50]. Pinocembrin complex with lecithin remarkably enhanced its antioxidant activity and its scavenging capacity for hydroxyl radicals, and such complexes may become a novel and potential possibility for AD therapy [51].
The flavonol morin (6) can block the progression of AD through its neuroprotective effects. It was suggested that cell death caused by Aβ oligomers in neuronal cultures, which in turn attenuated oxidative stress, was decreased in the presence of micromolar concentrations of morin. The neuroprotective effects of antioxidants on Aβ are related to a reduction in Aβ-induced mitochondrial calcium load, decreased mitochondrial membrane depolarization, and decreased release of cytochrome c, a key promoter of apoptosis, from mitochondria. Furthermore, morin can also stimulate the endogenous enzymatic antioxidant system and regain normal levels of oxidized protein. Morin can alleviate the Aβ-triggered energy homeostasis caused by decreased respiratory ability in AD cell models [20].
Icariin (51) is a flavonoid glycoside derived from the traditional Chinese herb Epimedium brevicornum. Extensive studies have shown that it alleviates the neurotoxicity caused by H2O2, endoplasmic reticulum stress, ibotenic acid, and homocysteine. Icariin promotes learning and memory in both normal aging animals and in disease models, and this provides an insight into AD progression avoidance. One study in CA1 pyramidal neurons of neonatal rat hippocampal slice suggested that icariin inhibits Aβ-induced neurotoxicity in a dose-dependent manner, and that the neuroprotective effect of icariin against Aβ2535-induced neurotoxicity is acted through balancing intracelluar calcium homeostasis [151]. Moreover, icariin was found to inhibit Aβ42-induced neurotoxicity by upregulating cocaine-regulated and amphetamine-regulated transcripts in Aβ42-treated neurons [152]. Icariin also increases the expression of key mitochondrial proteins, including cyclooxygenase IV, the pyruvate dehydrogenase component α subunit, and post-synaptic density protein 95 in the hippocampus and cortex [153], and two proteins, cyclin D1 and p21 that induce an increase in the number of neural stem cells in the brain [154]. Intragastric administration of icariin reversed the learning and memory impairment observed in the SAMP8 mice model of early AD. Icariin can also ameliorate amyloid pathologies by inhibiting neuronal apoptosis and improving autophagy-lysosome function. These effects can be achieved through up-regulation of the expression of anti-apoptotic protein Bcl-2 and downregulation of the expression of pro-apoptotic protein Bax [35], and suppression of Aβ1–42-induced cell loss, mitochondrial damage, nuclear membranes unclear and abundant nucleus chromatin agglutinates in the hippocampus, thereby lessening the expression of cleaved-caspase-3, brain oxidative stress, and astroglial activation [155]. Taken together, these effects contribute to the preventive and/or therapeutic effect of icariin in AD.
Cryptotanshinone (CTS) (58) is an active compound of the medicinal herb Salvia miltiorrhiza. Through MTT and lactate dehydrogenase assays, the effect of CTS on Aβ-induced toxicity to human neuroblastoma cells was studied, CTS reduced the cytotoxicity induced by Aβ42 and dramatically reduced Aβ42-induced cellular apoptosis and increased the level of reactive oxygen species in these cells [122]. CTS alleviated memory deficits in Aβ142-injected mice in a dose dependent manner. Notably, the analysis of gliosis-related and neuro-inflammatory markers in hippocampal tissues revealed a remarkable reduction in the expression of GFAP, S100β, COX-2, inducible nitric oxide synthase (iNOS), and NF-kBp65 after CTS (10 mg/kg) treatment [156].
The neuroprotective function of the flavonoid silibinin (48) may offer another potential approach to AD therapy. It has been demonstrated that silibinin can attenuate memory impairment in mice through amelioration of oxidative stress and inflammatory response induced by Aβ2535 in the Y-maze and novel object recognition tests [29, 157]. Repeated treatment with silibinin attenuated the Aβ2535-induced accumulation of malondialdehyde and the depletion of glutathione in the hippocampus [29]. Silibinin also suppressed expressions of p53 and p-p53 in the hippocampus of Aβ2535-injected rats [30]. Silibinin also attenuated the neuronal damage in the hippocampus of Aβ142-injected rats. Silibinin-treatment up-regulated the function through BDNF/TrkB pathway and attenuated autophagy in the hippocampus [31].
The protective activities of kaempherol (5) against Aβ-induced cytotoxicity were studied in the neuronal models of rat pheochromocytoma cells (PC12) and Institute of Cancer Research mice. It was shown to have protective effects against oxidative stress-induced cytotoxicity in PC12 cells, and significantly reversed Aβ-induced impaired performance in a Y-maze test [52]. Recent research has shown that kaempherol attenuates neuroinflammation and blood brain barrier dysfunction to improve neurological deficits in cerebral ischemia/reperfusion rats, whose mechanism is related to the NF-κB pathway [53].
Anthocyanins (ANT) (68), plant pigments referring to a flavanol class of polyphenols, offer multifunctional benefits including antioxidative and anti-inflammatory properties, cardioprotection, and chemoprevention for cancer [158]. They are potent antioxidants that may modulate the free radical-mediated generation of Aβ in the brain. And its antioxidant activity is related to the structure, mainly including the position of the hydroxyl groups, glycosylation patterns, and the nature of the sugars [159]. ANTs cross the BBB and protect brain tissue from Aβ toxicity, mitochondrial dysfunction, and apoptosis induced by oxidiatve stress [54]. Cyanidin-3-glucoside can protect cells from oxidative stress-induced apoptosis [55]. Malvidin and its analogue were shown to have protective effects against Aβ-induced neurotoxicity in neuro-2A cells by blocking ROS formation, preserving Ca2 + homeostasis, and preventing Aβ-mediated perturbation of certain genes involved in Aβ metabolism and cellular defense [160].
Fisetin (59) and butein (69) are active flavonoids. It has been reported that Rhus vernciflua inhibits glutamate-induced neurotoxicity in primary cultured rat cortical cells. Further research narrowed it down to getting neuroprotective fractions from the plant yielding flavonoid-rich ethyl acetate fraction (PREF). Fisetin not only exhibited an inhibitory effect against LPS-induced neuroinflammation by suppressing iNOS and cyclooxygenase (COX)-2, but also displayed memory enhancing effects via reactivation of cAMP responsive element binding protein (CREB)–brain derived neurotrophic factor (BDNF) pathway in memory-impaired mice by scopolamine. A single intracerebroventricular injection of Aβ142 (3μl/5 min/mouse) markedly induced memory/synaptic deficits, neuroinflammation, and neurodegeneration. Fisetin prevents neuroinflammation by suppressing various activated neuroinflammatory mediators and gliosis and blocks the apoptotic neurodegeneration triggered by Aβ142 injections in the mouse hippocampus, proving that it has a potent neuroprotective effect against Aβ142-induced neurotoxicity [41]. Butein showed a similar activity to that of fisetin, although to a lesser extent. Neuroprotection by PREF and selected flavonoids may involve maintenance of antioxidant defense mechanisms including glutathione peroxidase (GSH-Px), glutathione reductase, and superoxide dismutase activities [42].
Wogonin (70), an active flavonoid-like compound found in Scutellaria baicalensis Georgi, is considered a good excipient [56], can also act as an inhibitor of AD progression. It has been shown that wogonin decreases the levels of some inflammatory regulators, including TNF-α, IL-1β, and NO in LPS-stimulated murine microglial cells, of which undue activities may cause neuron dysfunction in danger of AD [161]. Wogonin can also downregulate the expression of upstream genes including iNOS and nucleus factor-kappa B (NF-κB) that is of great significance for accommodating the production of inflammatory mediators in primary microglial cells [74]. It has been experimentally shown that wogonin exerts its neuroprotective effects both directly and indirectly. Some AD patients exhibit excitotoxicity caused by an excess of L-glutamate [162], resulting in secondary cascades involving ROS and mitochondria-mediated apoptosis. Wogonin (300μg/mL) has been shown in a number of studies to significantly inhibit glutamate or NMDA-induced excitotoxicity [163] and to attenuate the neural oxidation induced by H2O2, xanthine/xanthine oxidase, and DL-buthionine [S, R]-sulfoximine (a glutathione depleting agent) [164–166].
Scutellarin (71), baicalin (37), and chrysin (72) are flavonoids extracted from the stems and leaves of the Baical skullcap. A total flavonoids fraction of Baical skullcap stems and leaves protected hippocampal neurons against damage induced by injection of Aβ2535 into the hippocampus in rats. The underlying mechanisms were related to its actions of decreasing the accumulation of lipid peroxide and proliferation of glial cells induced by Aβ2535 [58]. Another study conducted by Ye et al. demonstrated that total flavonoids alleviated memory and learning injury and protected hippocampal neurons from morphological changes in AD rats induced by Aβ2535 injection [59]. These studies suggest that the total flavonoids of Baical skullcap stems and leaves possess potential efficacy against AD, a possibility that needs further study.
Liquiritin (73), an extract of the root of Glycyrrhiza uralensis Fisch [167], possesses neuroprotective effects [61]. The neuroprotective mechanisms of liquiritin are related to the regulation of metabolic disorders, activation of the Nrf2/ Keap1 pathway, attenuation of the ERK1/2/NF-κB pathway, and suppression of the mitochondrial apoptosis pathway in PC12 cells [168]. Liquiritin inhibited mitochondrial apoptosis by decreasing Ca2 + concentrations, improving MMP, upregulating Bcl-2, and downregulating Bax, cytochrome C, cleaved-caspase-3 expression levels. Yang et al. found that pretreatment with liquiritin on primary cultured rat hippocampal neurons for 6 h decreased the elevated intracellular Ca2 + concentration and neuron apoptosis caused by Aβ2535. Liquiritin is also capable of enhancing the effects of nerve growth factor on the extension of neuroaxons [169]. Liquiritin also specifically inhibits the activity of acetylcholinesterase and promote the differentiation of neuronal stem cells into cholinergic neurons [60, 169]. These neuroprotective and neurotrophic effects make liquiritin a promising agent against AD.
Naringenin (74), a natural predominant flavanone, possesses a broad range of biological and pharmacological activities [170]. A previous study examined the neuroprotective effect of naringenin from Citrus junos against oxidative cell death induced by the Aβ peptide in PC12 cells. They showed that pretreatment with naringenin prevented the generation of Aβ-induced ROS and decreased Aβ toxicity in a concentration dependent manner [62]. Naringenin in silico exhibits a robust affinity for AMP-activated protein kinase (AMPK) and upregulates AMPK-mediated autophagy signaling in neurons via inducing autophagy triggering proteins such as ULK1, Beclin1, ATG5, and ATG7 in Neuro2a cells and primary mouse neurons. It also ameliorates the Aβ content at nontoxic concentration in neuronal cells. It has been reported that naringenin maintains the mitochondrial membrane potential and attenuates ROS production, resulting in protection against Aβ1–42 evoked neurotoxicity [171].
Apigenin (7), has also been shown to have potent antioxidant, anti-inflammatory, and anti-carcinogenic properties [22]. It exhibited superoxide anion scavenging effects and improved the antioxidative enzyme activities of superoxide dismutase and glutathione peroxidase [21]. In addition, apigenin restored the neurotrophic ERK/CREB/BDNF pathway in the cerebral cortex. It was reported to protect rat cortical neurons against Aβ-induced neurotoxicity by inhibiting the caspase pathway, rather than through its anti-oxidative capacity [172]. In addition, investigation of the anti-amnesic and protective effects of apigenin against Aβ2535-induced toxicity and the underlying mechanisms in the cerebral cortex in mice demonstrated a coupling of modulation of microvascular function and neuronal activity, that might be due to the BBB protection, cortical cholinergic improvement and modification of BDNF-TrkB-pCREB levels [173]. An animal study demonstrated that apigenin pre-treated mice (10–20 mg/kg) subjected to contusive spinal cord injury had decreased levels of IL-1β, TFN-α, intercellular cell adhesion molecule-1 (ICAM-1) and caspase-3, and an increased Bcl-2/Bax ratio [174].
Rutin (3,3’,4’,5,7-pentahydroxyflavone-3-rhamnoglucoside) (54), a flavonoid with a wide range of biological activities, is found in many plants (such as buckwheat seeds), fruits (such as citrus fruits), and vegetables [175, 176]. Rutin has been shown to exert neuroprotective effects on AD transgenic mice [39]. Studies have shown that rutin can dose-dependently inhibit Aβ42 fibrillization and attenuate Aβ42-induced cytotoxicity in SH-SY5Y neuroblastoma cells [177]. Another study suggested an effect of rutin on the mitogen-activated protein kinase pathway, memory retrieval and oxidative stress in rats injected with Aβ. Rutin injection significantly increased the gene expression levels of ERK1, CREB, and BDNF in the hippocampus, augmented memory retrieval, and significantly decreased the malondialdehyde level compared to the control. The content of sulfhydryl groups in the rutin group was higher than that in the control group. The findings suggest that rutin may have beneficial effects against the neurotoxicity of Aβ on memory in rats [178].
Luteolin (8) also exerts its anti-AD effects by acting as anti-inflammatory agent. Some studies have also reported that the neuroprotective effect of luteolin is related to its antioxidant activity [179]. It can suppress amyloidogenesis, which contributes to increased inflammation in AD. By promoting inflammation, fibrillary Aβ can disrupt an in vitro BBB, an in vitro BBB model by co-culturing human brain microvascular endothelial cells and human astrocytes (hAs) under fAβ140-damaged conditions. However, treatment with luteolin (3–30μM) can preserve the function of the BBB by inhibiting the inflammatory response. Luteolin can also effectively reduce the levels of inflammatory mediators and cytokines (e.g., COX-2, IL-1β, and IL-6) induced by fibrillary Aβ. The anti-inflammatory action of luteolin is mediated by inhibition of the activation of p38 mitogen-activated protein kinase and NF-κB [25]. These discoveries provide a new line for therapeutic intervention in AD.
CONCLUSION
Flavonoids are the most important active ingredients among natural products of traditional Chinese medicine. In-depth research on the anti-AD effect of flavonoids can help elucidate the mechanism of TCM theory in treating AD with modern medicine. This review discusses some natural flavonoids, which play a role in slowing or halting the onset of AD through a variety of pathways, including Aβ production, Aβ oligomerization, and Aβ-induced neurotoxicity. In presenting the information, we expect to provide alternative and attractive targets for the discovery of natural therapeutic agents against AD.
Flavonoids belong to the broad group of polyphenols and, over 6,000 different flavonoids identified to date can be divided into several subclasses according to their different constituents, including flavanones, flavone, flavanols, flavonols, and others. The general flavonoid structure consists of a flavan nucleus in which, 15 carbon atoms are arranged in three rings (A, B, C). The various classes differ in their level of oxidation and in their pattern of substitution of the C-ring.
The inhibitory potency of BACE inhibition by flavonoids was significantly increased by the presence of free two hydroxyl groups in the resorcinol moiety (B-ring) and isoprenyl functionality at C3 in the ring A of flavone, and the presence of a C3-OH group plays a significant role in BACE inhibition. Furthermore, ring closure of the prenyl group onto C7-OH to form the pentacyclic structures also decreases BACE inhibitory activity, suggesting that a free C7-OH group plays a crucial role in BACE inhibitory activity. For amentoflavone-type biflavonoids, compounds consisting of two apigenin molecules linked at the C3’- C8” position are important for BACE-1 inhibitory activity and more than two hydroxyl groups at the R1-R4 position are needed for inhibitory activity. Studies have shown that flavonoids inhibit BACE by directly interacting with the BACE-1 catalytic core through hydrogen bonds, and that the interaction of flavonoids with catalytic Asp residues (particularly Asp 32) is more important than with other residues. Residues such as Trp198 and Gln73 can also help stabilize the docking conformations of flavonoids with BACE, which affects the inhibitory activity.
The following conclusions focus on the structural requirements for the inhibitory effect of flavonoids on Aβ fibril formation. The 3’, 4’-dihydroxyl group of the B ring is essential for the inhibitory effect on Aβ fibril formation, and loss of the 3’- or 4’-hydroxyl group converts the effect from inhibition to enhancement. Furthermore, myricetin inhibits Aβ fibril formation more potently than fisetin, suggesting that increasing the number of hydroxyl groups on the B ring potentiates the inhibitory effect. While neither the 3-hydroxyl group nor the 7-hydroxyl group is necessary for inhibition of Aβ fibril formation, loss of the 3-hydroxyl group has little influence on the inhibitory activity. Finally, hydroxylation at the 5-position may contribute to the inhibition of Aβ fibril formation, because a comparison between fisetin and quercetin demonstrates that 5-hydroxylation increases the inhibitory activity or suppresses the enhancing activity.
The molecular mechanisms underlying inhibitory effects of flavonoids on Aβ fibril formation remain unclear. This is the key point blocking the progress of available therapies. Although there are a number of hypotheses on AD pathogenesis, these hypotheses are still undergoing clarification. It is difficult to effectively control disease progression using a single drug that targets a specific target. Thus, flavonoids can be potentially invaluable as neuroprotective drugs delaying the progression of AD, due to their multi-target, multi-channel, and multi-system characteristics.
Extensive work has been done to demonstrate the multi-functional capacity of flavonoids in inhibiting cholinesterase activity and oxidative stress, scavenging free radicals, modifying tau protein, reducing Aβ accumulation, and inhibiting inflammatory cytokine expression. Apart from this, flavonoids are ubiquitous in nature and have low toxicity, thus, they are the most important component of active ingredients in traditional Chinese medicine. It will be necessary to further explore the pharmacological activity mechanisms and structure-activity relationships of flavonoids, and to focus on the development of new drug delivery systems, in order to explain the mechanism and to lay a solid foundation for their better application.
Footnotes
ACKNOWLEDGMENTS
This work was supported by grants from the National Major Scientific and Technological Special Project for “Food Safety” (No. 2019YFC1604604). C.Z. was responsible for the ideation of the article; Q.W., X.D. and R.Z. made contributions to the literature collection, data analysis, paper writing, and post-revision work. All authors have read and agreed to the published version of the manuscript.