
Abstract
Background:
The coexistence of magnetite within protein aggregates in the brain is a typical pathologic feature of Alzheimer’s disease (AD), and the formation of amyloid-β (Aβ) plaques induces critical impairment of cognitive function.
Objective:
This study aimed to investigate the therapeutic effect of proton stimulation (PS) targeting plaque magnetite in the transgenic AD mouse brain.
Methods:
A proton transmission beam was applied to the whole mouse brain at a single entrance dose of 2 or 4 Gy to test the effect of disruption of magnetite-containing Aβ plaques by electron emission from magnetite. The reduction in Aβ plaque burden and the cognitive function of the PS-treated mouse group were assayed by histochemical analysis and memory tests, respectively. Aβ-magnetite and Aβ fibrils were treated with PS to investigate the breakdown of the amyloid protein matrix.
Results:
Single PS induced a 48–87%reduction in both the amyloid plaque burden and ferrous-containing magnetite level in the early-onset AD mouse brain while saving normal tissue. The overall Aβ plaque burden (68–82%) and (94–97%) hippocampal magnetite levels were reduced in late onset AD mice that showed improvements in cognitive function after PS compared with untreated AD mice (p < 0.001). Analysis of amyloid fibrils after exposure to a single 2 or 4 Gy proton transmission beam demonstrated that the protein matrix was broken down only in magnetite-associated Aβ fibrils.
Conclusion:
Single PS targeting plaque magnetite effectively decreases the amyloid plaque burden and the ferrous-containing magnetite level, and this effect is useful for memory recovery.
Keywords
INTRODUCTION
Combined irradiation of hadron or protons with target-delivered medium-to-high Z nanoparticles induce a site-specific dose-enhancement effect (release of low energy electrons and X-ray fluorescence), as depicted in Fig. 1 [1–8]. The existence of magnetite in amyloid-β (Aβ) plaques [9–12] provides the unique opportunity to use proton stimulation (PS) therapy targeting endogenous magnetite in the Alzheimer’s disease (AD) brain without the need to deliver exogeneous nanoparticles for dose enhancement. With the Bragg peak behind the body [5, 8], a 100 MeV proton transmission beam is capable of traversing the entire mouse brain to selectively activate plaque magnetite, thus exerting a therapeutic effect only on the target plaque by proton fluence that is comparable with the Bragg peak dose [13, 14]. PS using a transmission beam for the treatment of AD is distinct from the spread-out Bragg peak used for conventional oncological proton treatment, in which the Bragg peak is placed inside the tumor volume.

Schematic diagram of proton stimulation with a Bragg peak behind the body (28) where Bragg peak was placed outside brain by high energy proton transmission beam compared with the depth of mouse head. Low-energy electron (LEE) and characteristic X-ray fluorescence (XFL) are emitted from proton-stimulated magnetite (Fe3O4) in AD protein aggregates via Auger cascades and the interatomic Coulomb decay path (right panel). The proton fluence during the transmission of brain tissues containing Aβ plaques with magnetite abruptly decays to zero at the Bragg peak position, resulting in the deposition of all energy, but the plateau dose is absorbed by the tissue along the beam path, resulting in the stimulation of nanoparticles with comparable Bragg peak dose (BPD) (left panel).
Amyloid plaques and neurofibrillary tangles comprise highly insoluble and densely packed filaments that contain high concentrations of iron in the AD brain [15–17]. Numerous studies have established that ferrous magnetite is localized within Aβ plaques [9–12]. Thus, it has been proposed that ferrous magnetite is a major source of toxicity caused by Fenton reaction-induced reactive oxygen species (ROS) production. Magnetite integrated in the protein matrix of Aβ plaques is relatively enriched Fe2+ over Fe3+ due to Aβ-mediated reduction by binding with Aβ, resulting in a redox-toxic state. Ferrous iron catalyzes the formation of hydroxyl radicals by reacting with hydrogen peroxide, which is formed by freely permeable mitochondrial superoxide ions in tissue. The surface of plaque magnetite, which presents both Fe2+ and Fe3+, may act as a catalyst for the Haber-Weiss cycle and the Fenton reaction [18, 19], leading to increased formation of ROS, such as hydroxyl radicals. Therefore, insoluble Aβ plaques containing ferrous magnetite, if not eliminated, are continuously neurotoxic. Previously, we performed a proof-of-concept study of the effect of PS on Aβ-magnetite fibrils in vitro, demonstrating that the treatment induces Aβ fibril and Aβ-bound ferrous magnetite degradation [20]. To investigate the in vivo role of single PS on Aβ plaques containing magnetite, we subjected AD mouse brains to PS to irreversibly and safely break down Aβ plaques in this study.
The coexistence of redox-toxic ferrous magnetite with diffusive immature plaques and the mature Aβ plaque core suggests that metal integration into the protein matrix at the onset of AD [9, 22] leads to significant cognitive defects [23–25]. An electromagnetic field treatment targeting plaque magnetite was shown to protect against and alleviate cognitive impairment in AD mice [26]. Considering that current AD therapies have failed despite the prevailing theory that soluble Aβ oligomers (soAβs) are toxic, recent discoveries related to the iron-dependent cell death process termed ferroptosis [27] and plaque magnetite have provided an emerging framework to understand iron-driven neurodegeneration, thus providing new opportunities to treat AD by targeting iron. Here, we demonstrate novel therapeutic strategies for AD targeting redox-toxic magnetite in Aβ plaques by deep-brain stimulation with proton transmission beams.
MATERIALS AND METHODS
Design of the in vivo PS experiment
In this study, early-onset APP/PS1 mice and aged APP/PS1 and 5xFAD mice were treated, and the average number of plaques was counted. First, seven cohorts of female early-onset APP/PS1 mice (n = 21, median age of 12 weeks) received single PS, and the plaque burden was measured at three different time points (separate mouse group for the 1st, 3rd, and 7th days) after PS to determine the optimal time point for examining plaque reduction and the effect of the proton dose on irreversible plaque removal. The effect of PS treatment on amyloid pathology and associated iron toxicity in the mouse brain was measured by histological methods. Second, aged APP/PS1 (n = 7, female, mean age of 41.2 weeks) and 5xFAD (n = 16, mixed gender, mean age of 31.8 weeks) (n = 23) received single or fractionated PS, and the cognitive function of all animals and Aβ-plaque/ferrous magnetite levels in those showing improvement of cognitive function were assessed. To investigate the safety and effects on memory recovery of PS, the performance of both aged AD mice and non-Tg WT mice (n = 4) in the RAM and MWM tests were tested prior to and one month after PS. Among the 23 aged AD mice, 10 mice were subjected to both pre- and post-PS memory tests, whereas the others were subjected to either a pre-PS or post-PS test only. The correlation between the histological results and the results of the memory tests was analyzed, and the results of Congo red staining were averaged for randomly selected mice that showed improvement in memory after PS and untreated mice.
Animals
All animal experiments were approved by the Institutional Animal Care and Use Committee (DCIAFCR-181029-22Y). All applicable international and institutional guidelines for the care and use of animals were followed. 5xFAD [B6SJL-Tg(APPSwFlLon,PSEN1*M146L*L286 V)6799Vas/Mmjax] and APP/PS1 [B6. Cg-Tg(APPswe,PSEN1dE9)85Dbo/Mmjax] transgenic mice were purchased from the Jackson Laboratory (Bar Harbor, ME, USA).
Proton irradiation
Proton irradiation was performed at KOMAC TR102 (Kyungju, South Korea). Intraperitoneal injections of 100 mg/kg ketamine and 20 mg/kg xylazine cocktail were administered to mice for anesthesia. A proton transmission beam with a pristine Bragg peak behind the body [5, 28] was applied to the entire brain (PS), as shown in Fig. 1. The irradiation energy was 100 MeV, which is sufficient to penetrate the mouse head, and a single entrance dose of either 2 Gy or 4 Gy, as measured by a TM30013 Farmer chamber (PTW), was applied to the frontal surface of the sample. The dose rate was 0.02 Gy/s. For fractionation, irradiation was performed at one-month intervals; for 2 + 2 Gy fractionation, 2 Gy PS was administered twice at a one-month interval for a total of 4 Gy.
Sample preparation
The animals were sacrificed by a lethal dose of ketamine and xylazine cocktail and perfused intracardially with normal saline and then with ∼50 ml of fixative containing 4%(wt/vol) paraformaldehyde in PBS (pH 7.4). The brains were harvested, postfixed in fresh fixative overnight, and subsequently placed in PBS. The samples used for histological staining were embedded in paraffin, sectioned at a thickness of 10μm and mounted on glass slides.
Histological staining of plaques and tissue damage
Sections with a thickness of 10μm from AD mice were stained with acidified 1%(wt/vol) potassium ferricyanide for Turnbull staining of Fe2+ iron, with 5% potassium ferrocyanide for Perl’s staining of Fe3+ iron, or with 0.2%Congo red in 50%alcohol for staining of amyloid plaques using a modified Highman’s Congo Red staining protocol [29]. The sections were stained with Congo red for approximately 10 min, and after the tissues were washed to remove unbound Congo red, they were stained with hematoxylin for 2 min. The stained sections were examined with a microscope. The area and number of stained plaques were determined per section either manually for the early-onset cohort or by automated analysis for the aged cohort, which contained mice with a relatively large number of plaques and averaged for each section or mouse. ImageJ software, which is available at https://rsbweb.nih.gov/ij/, was used for analysis of amyloid plaques and Fe2+/Fe3+ particles. For automated counting, the intensity of red, green and blue staining within the identified plaque margin was determined in microscopic images. Using the MATLAB, pixels that were not in the colored region of the plaque were assigned a color of black in a pixel-by-pixel manner. The processed image was saved, loaded in ImageJ, and converted to 8 bits, resulting in a grayscale image. Typical plaque regions not containing noise were chosen in a standardized manner from typically 3-4 plaques by adjusting the threshold value to exclude noise pixels (typically 1 or 2) in the plaques using the built-in threshold function. An assigned threshold value was applied to select all the plaques within an imaging slice and set them to black with a white background, resulting in black and white images. Plaques were counted using the analyze particle function by setting the pixel size to 30 and the circularity to 0.3–15.0. To determine Aβ plaque or iron load, data were obtained from 15 or 4–6 mouse brain sections including the frontal cortex and hippocampus from early-onset or aged mice, respectively, at×200 magnification and used for statistical analysis. Sections from PS-treated mice were stained with hematoxylin and eosin (H&E) for the assessment of erythrocyte extravasation, edema or vascular damage and with cresyl violet (Nissl staining) for the assessment of neuronal degeneration.
Radial arm maze test
Untreated and non-Tg wild-type mice were subjected to the RAM test to assess neurocognition before and after PS. Spatial working memory was tested using an eight-arm radial maze as previously described [30]. Briefly, the maze consisted of a central chamber with eight equally spaced arms extending outward. One reward cup was located on a platform at the distal end of each arm. Each mouse first underwent 10-min habituation training trials in the radial arm maze on 3 consecutive days. After acclimation, the mouse was given free access to all open arms for the duration of the testing session (two arms separated by 135° were baited), allowing it to find the baited arms. The trial was terminated when the animal located the baited arms and consumed the food reward. The number of arms that the mouse visited before visiting the two baited arms, including revisits, was counted. A choice was counted as correct only if the mouse approached and ate from the baited food cup. A visiting error was considered a spatial working memory error and occurred when a mouse re-entered an arm that was unbaited or had been previously baited. These performance measures were acquired before and after PS treatment and analyzed by analysis of variance (ANOVA).
Modified morris water maze (MWM)
The MWM test was performed using a similar apparatus as that described previously [31]. Briefly, the water maze apparatus consisted of a circular pool (1.2 m in diameter and 0.47 m in height) that was filled to a depth of 20 cm with water (24–25 °C), and the water was made opaque by the addition of nontoxic white paint. During conventional reference memory training in the MWM, an escape platform (10 cm in diameter) was submerged 0.5 cm under the water surface in the same location of the pool (NE quadrant). Mice were released into water from the same point in all trials. For each mouse, the differences in the escape latency and the time spent on the escape platform between two consecutive trials were assessed.
Aβ-magnetite-targeted PS
Magnetite was synthesized using a standard coprecipitation method [32]. Aβ42 peptide was obtained from a commercial manufacturer (AnyGen, AGP-8338, Seoul, Korea). Aβ-magnetite fibrils were prepared in a glovebox fume hood in an atmosphere of N2 to prevent changes in the oxidation state of the iron. Monomeric synthetic Aβ42 was dissolved in 0.1 M NaOH to obtain a 220μM Aβ stock solution. This Aβ stock solution was left standing for 30 min to ensure complete peptide dissolution prior to mixing with the 80μg/mL magnetite solution. Aβ-magnetite solutions containing thioflavin T (ThT) were incubated at 37°C for 144 h, and the fluorescence of ThT in a series of aliquots was used as a measure of fibril formation over time. Aβ fibrils were prepared as previously described [33]. A stock solution of Aβ peptides was prepared as follows: 1 mg of peptide was dissolved in 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP) at a concentration of 500μM. Fresh Aβ stock solution was diluted in 20 mM HEPES and 100 mM NaCl, pH 7.4, containing 3.3μM ThT to a final concentration of 5μM. A total of 400μl of each sample was incubated at 40°C. Fibrillization was monitored using ThT fluorescence, which was measured at 480 nm with an excitation at 445 nm. A 10-μL aliquot of each Aβ-magnetite or Aβ solution was pipetted onto carbon/Formvar-coated copper transmission electron microscopy (TEM) grids (TED PELLA Inc.), and the excess liquid was removed to allow the deposition of Aβ-magnetite or Aβ aggregates. Each TEM grid with bound Aβ-magnetite or Aβ fibril complexes was attached to the bottom of each single well and irradiated using a 100 MeV proton transmission beam with a plateau dose of 2 or 4 Gy. The dose rate was measured as 0.1 Gy/s in the front of the sample.
Synchrotron X-ray spectromicroscopy
X-ray spectromicroscopy experiments were performed at BL-13A of PF in Tsukuba Japan. Energy-specific images were created by raster scanning the focused beam (the X-ray spot size was ∼30 nm) across the area of interest in the sample with sample exposure times < 2 ms per pixel for each image and recording the transmitted X-ray intensity. An iron L3-edge absorption image was obtained at 710 eV, and the background at 700 eV was subtracted. X-ray absorption spectra were obtained from 700 eV to 720 eV for the iron L-edge. The STXM data were processed using the aXis 2000 software package available at http://unicorn.mcmaster.ca/aXis2000.html. For the iron L2,3 X-ray absorption spectra, a three-point smoothing filter was applied to the raw data.
Infrared spectroscopy of Aβ-magnetite
Infrared spectroscopy experiments were performed at the 12D beam line of the Pohang Accelerator Laboratory (PAL) in Pohang, Korea. Mixtures of Aβ42 and magnetite nanoparticles were incubated under anoxic conditions for one week at 37°C, whereas Aβ42 monomer was prepared just prior to data acquisition. PS was performed on a TEM mesh on which Aβ-magnetite fibrils were mounted. FTIR spectra were obtained using a Bruker Vertex 80/v FTIR spectrometer equipped with a Hyperion3000 at the 12D IRS beamline of PAL. FTIR spectra were recorded at a resolution of 4 cm–1 using a mercury cadmium telluride detector in transmission mode with a 15×objective. The initially obtained spectra were subjected to deconvolution of the amide I band using Gaussian curves with fityk-1.3.1 available at https://fityk.nieto.pl.
Statistics
The data are presented as the means±SEMs. One-way ANOVA followed by post hoc tests was performed using SAS software (SAS Institute, NC, USA). Repeated-measures ANOVA was also used to evaluate the RAM test data. Student’s t-test was used to compare the difference between two groups. Differences were considered statistically significant at p < 0.05.
RESULTS
A single PS reduces the amyloid plaque load and the level of ferrous-containing magnetite in early-onset transgenic AD mice
In this study, 21 Aβ plaque-forming early-onset APP/PS1 transgenic mice [34, 35] were subjected to a single PS. The PS-treated mice were sacrificed a few days later (at three time points after treatment; 1, 3, and 7 days) for histological analysis.
We next performed Congo red, Perl, and Turnbull staining to assess amyloid plaques, ferric iron, and ferrous iron/magnetic nanoparticles dispersed in Aβ deposits (Fig. 2A-C). We analyzed the area of each plaque and iron foci and the number of plaques and iron foci in selected sections from each mouse (a total of 15 sections per mouse) and found that the cortical and hippocampal area occupied by plaques and ferrous iron foci was reduced by 66%and 85%, respectively (Fig. 2A, B), and that the average number of plaques and ferrous iron foci per mouse at 7 days after PS was reduced by 72%and 87%, respectively, in the proton-treated mice compared with the untreated mice. The intraplaque staining density was greatly reduced in the proton treatment group compared with the untreated control group, as shown in Fig. 2D and 2F. In Fig. 2A-C, the dependence of the reduction in the plaque/iron load on the day after the onset of treatment demonstrated an indicative of irreversible change in the plaque burden.

Graph showing the gradual reduction of Congo red-stained Aβ plaques, p value for * or ** was < 0.0001, < 0.0001, respectively (a) and Turnbull-stained ferrous, p value for † or †† was 0.0288, 0.0290, respectively (b) and ferric magnetite foci, p value for ‡ or ‡‡ was 0.0534, 0.0217, respectively (c) in early-onset AD (3-month-old APP/PS1) mouse brains at different time points (1, 3, and 7 days) after PS treatment. For counting iron staining, number density of intraplaque iron foci was analyzed and averaged per section and mouse. The analysis of the morphological features of typical Aβ plaques (d, f) and ferrous iron (e, g) untreated and 7 days after PS treatment showed the change in density from dense in the untreated tissue to degraded and sparse in the treated tissue, and the size gradually decreased over time after treatment. Areas in graph was displayed as number of pixels in microscopic image.
Correlation between the reduction in Aβ plaque burden and improvement in cognitive function in aged AD mice
To assess the correlation between the reduction in the Aβ plaque/ferrous magnetite burden and improvement in cognitive function, untreated and PS-treated (4 Gy, single or 2 + 2-Gy fractionation) aged AD mice (n = 23, aged 5–12 months) and non-Tg WT (n = 3) and untreated early-onset AD mice (n = 6, aged 1–4 months) were subjected to two spatial memory tests, namely, the radial arm maze (RAM) and modified Morris water maze (MWM) tests, and the results were compared. Among the mice exposed to 4 Gy PS (n = 16), 81%showed improvement in one memory test, and 54%showed improvement in both tests. Randomly selected mice that showed improvement in one test and nontreated aged AD mice were sacrificed for histologic analysis of the remaining Aβ plaques and ferrous magnetite levels. Compared with untreated AD mice, all the mice that showed improvement demonstrated a marked reduction in plaque burden (68–82%), as shown in Fig. 3 and Table 1 (p < 0.0001).
Congo red staining images of cortical and hippocampal tissues from typical untreated (left panel) and 4Gy PS-treated (right panel) aged APP/PS1 mice (A) and 5xFAD mice (B). Turnbull staining image (5xFAD) (C) showed clearly a retained intraplaque distribution of iron foci in untreated one (left panel) compared with a reduction and more likely scattered distribution of both cortical and hippocampal ferrous iron without apparent protein matrix in 4-Gy PS-treated one (right panel) (iron foci appeared within dotted circle).
Results from Congo red and turnbull staining of untreated and PS-treated brain tissues of the memory test-positive aged mice
4 Gy-PS treated mice showed 82%reduction in plaque burden compared with untreated mice (p < 0.0001). Hippocampal magnetite level in a PS-treated 5xFAD mouse was reduced as much as 97 %compared with untreated one. *Number of individual Aβ plaque or Fe foci was counted manually from microscopic image. #Aβ plaque was counted with automatic image program from microscopic image of the haploid brain (averaged over three mice per each group where 4–6 sections were measured in each mouse).
Single and 2 + 2-fractionated irradiation induced similar improvements in cognitive function. The results of the cognitive tests, including the accumulated error frequency (AEF) in the RAM test and the difference in escape latency (DEL) in two consecutively cued MWM test trials before and after PS for mice, are summarized in Fig. 4. All the tested groups showed significant improvement over time after treatment and exhibited intact learning acquisition and spatial short-term working memory (see Supplementary Figure 1). The AEF was significantly different before and after PS, as shown in Fig. 4. The non-Tg WT and early-onset AD mice showed a lower AEF than the aged AD mice. The early-onset AD mice made fewer errors after training than the aged AD mice (p < 0.01). The daily average error frequency was different before and after PS (Supplementary Figure 1) but the difference was not statistically significant, as determined by repeated measures ANOVA, due to large fluctuations between mice. Compared with aged mice prior to PS, PS-treated AD mice showed a significant reduction in error rate in the RAM test one month after treatment (p = 0.0019), and this error rate was on similar to that of the non-Tg WT mice. However, no significant change in error rate was observed one week after treatment, which suggested a delay in the recovery of neurocognitive function despite extensive removal of Aβ plaques. Moreover, augmented DELs were observed for the aged AD mice prior to PS treatment, as shown in Fig. 4. Therefore, the results suggest that impairment of memory function in AD mice was alleviated one month after PS. However, lack of improvement in the RAM test was observed several months after treatment for a few mice that had previously shown improvement in the RAM test; In case 1, AEF of RAM test was shown from 27 to 39 at four month after PS while showing still MWM+ result at that time: In case 2, DEL of MWM test was shown from -82 to 23 six months later after PS, while showing still RAM+ at that time: In case 3, AEF of RAM test showed a change from 27 to 64 four months later and further change to 19 another one month later.
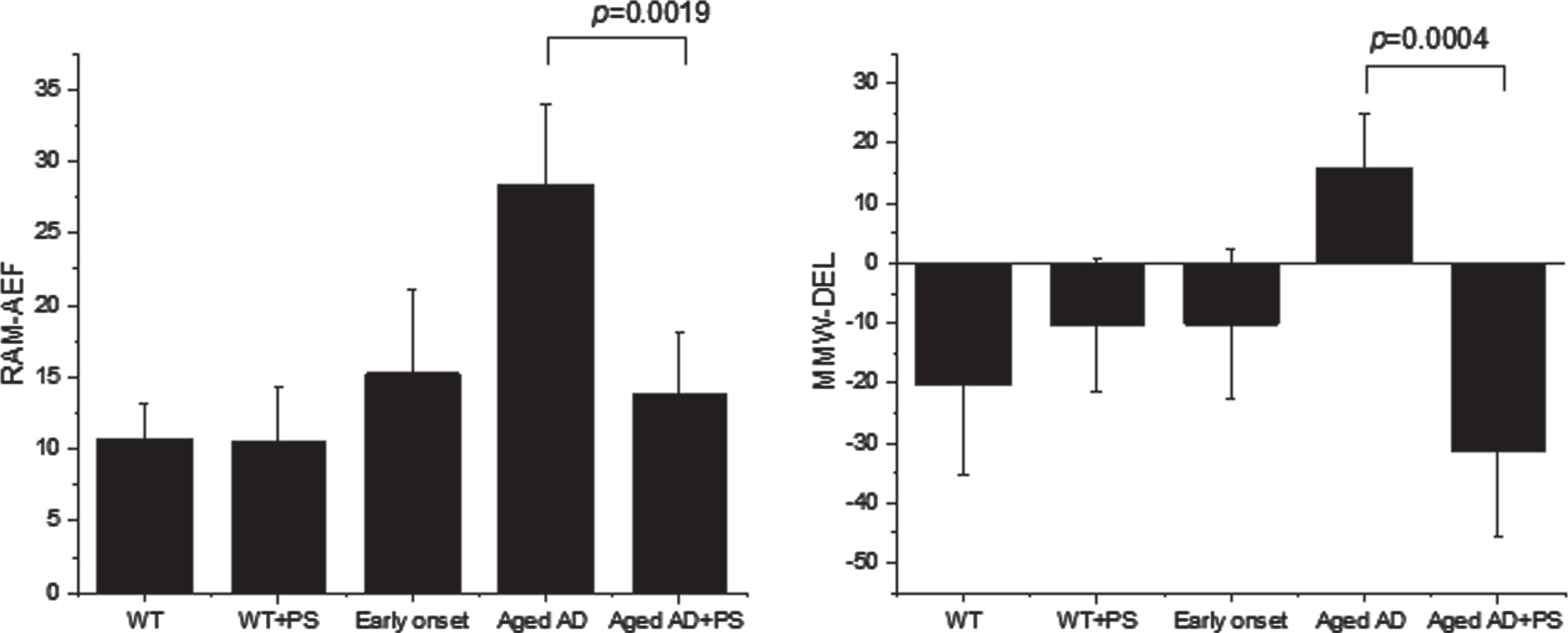
The RAM (a) or water maze (b) test was performed both before and after PS. AEF, accumulated error frequency; DEL, difference in escape latency from consecutively cued tests. A higher cumulative error in the RAM test was obtained with aged AD mice, and a decreased cumulative error was obtained after PS treatment (a). The differences in the escape latency (DELs) between two consecutively cued MWM tests showed a contrast between the positive DELs obtained for untreated mice and the negative DELs obtained for PS-treated aged AD mice. Non-TG WT and untreated early-onset AD mice also showed negative DELs (b).
Iron detoxification in aged AD mice
The oxidation state of iron in selected iron foci from representative iron-containing deposits in the entorhinal cortex and hippocampus of the mouse brain was examined by nanoscale-resolution STXM-XAS and compared between untreated and PS-treated mouse brain tissue. Fe2+ ions with a stronger peak at ∼708 eV and a weaker peak at ∼710 eV, representing dominant ferrous iron [Fe(II)] were identified in untreated brain tissue, and Fe3+ ions with a stronger peak at ∼710 eV and a weaker peak at ∼708 eV were identified typically in PS-treated brain tissue, suggesting the conversion of ferrous iron to ferric iron [Fe(III)] (Supplementary Figure 2). The cortical and hippocampal area that this effect occurred was analyzed by Turnbull staining, as shown in Fig. 3C and summarized in Table 1. Hippocampal magnetite levels in PS-treated mice were decreased by more than 95%compared with those in untreated AD mice.
PS did not induce overt damage
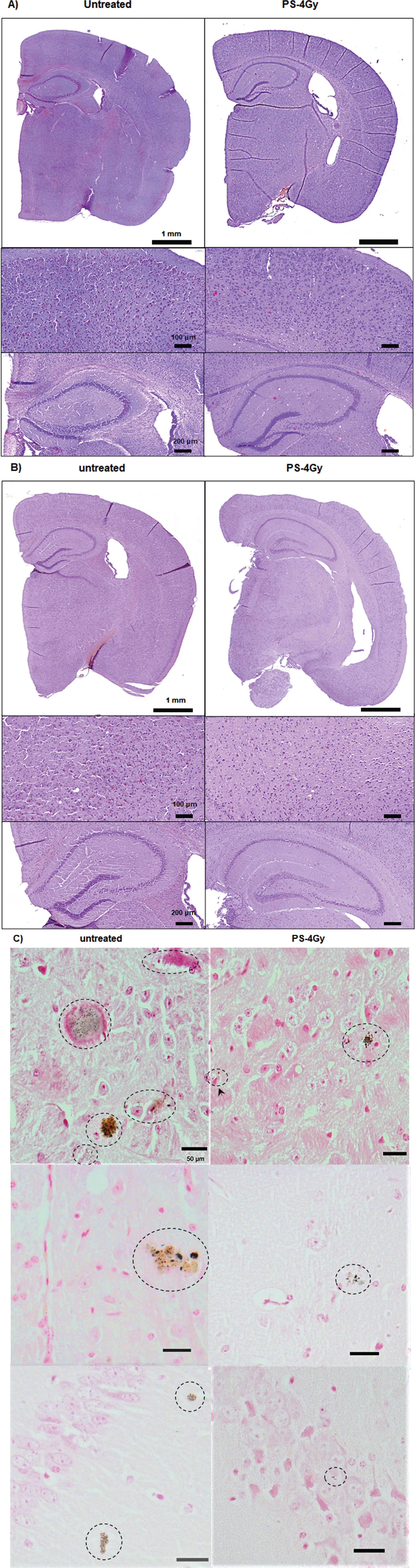
A low linear energy transfer (LET) plateau dose in the surrounding nontarget area did not cause degeneration of neurons in the AD brain one week after PS at a dose of either 2 or 4 Gy, as shown by Nissl staining (Fig. 5A). In addition, neither edema nor erythrocyte extravasation was observed, as shown by H&E staining (Fig. 5A), which suggested that PS did not induce any attributable ROS-mediated damage. No similar damage was observed in mice several months after PS (Supplementary Figure 3). In addition, the treatment of non-Tg WT mice with 2- to 4-Gy PS did not induce any significant histochemical changes related to degeneration or edema, as shown in Fig. 5B, which suggested that PD absorption after proton transmission did not cause overt damage to normal brain tissue.

A) Nissl (left-hand panel) or H&E staining (right-hand panel) of brain tissues from untreated and PS-treated mice. The results did not reveal any significant degeneration or vascular damage. A relatively long-term (more than one month) assay of the effects of PS treatment on AD mouse did not show any morphological changes in the surrounding normal tissue, as shown in the histological analysis based on H&E staining (panel 4-Gy PS). B) Results of Nissl and H&E staining of hippocampal sections from non-Tg WT mice after 4-Gy PS treatment demonstrated no significant damage (degeneration, edema, microvessel damage) from 4-Gy PS.
Degradation of the protein matrix by PS
Formation of Aβ fibrils was strongly promoted by magnetite with a more complex network, as shown in Fig. 6 [36]. TEM analysis revealed degradation of the protein matrix in the magnetite-associated Aβ fibrils after PS, as shown in Fig. 6. Aβ-magnetite fibrils were exposed to a low-dose (2–4 Gy) proton transmission beam, and degradation increased with the proton dose; however, no degradation was observed in Aβ-only fibrils, suggesting that PS has a magnetite-sensitizing effect [8]. The results of synchrotron-based IR spectroscopy shown in Fig. 7 and HPLC-eluting analysis (Supplementary Figure 4) demonstrated a major conformational change of Aβ from β-sheets to α-helices/random coils and fragmentation-like damage to Aβ upon PS of Aβ-magnetite fibrils, respectively.

TEM images of Aβ fibrils (upper panels, a–c) and Aβ-magnetite fibrils (lower panel, d-f) for untreated (a and d) and after PS (2 Gy: b, e; 4 Gy: c, f). Scale bar: 1μm. All images were obtained with 100.0 kV and 8.0μA. Aβ-magnetite fibrils were critically disrupted and degraded after PS compared with Aβ fibrils.

IR spectral features of a) an Aβ monomer, b) untreated Aβ-magnetite, and c) 4-Gy PS-treated Aβ-magnetite identified by decomposition of the raw spectra (see SM-4) into secondary structural components. The identified features included the alpha helix of Aβ alone (a), the introduction of a dominant β-sheet structure after Aβ-magnetite fibril formation (b), and a mixture of random coils after PS-induced degradation (c).
DISCUSSION
The in vivo PS treatment studied here induced both the detoxification of ferrous magnetite and the removal of Aβ plaques obtained from an Aβ fibril model. The marked reductions in plaque burden and redox-toxic magnetite levels may be ascribed to the selective dose-enhancement of relatively higher Z-numbered iron oxide nanoparticles in comparison to tissue elements by transmitting protons, which is in contrast to the effect of low-dose X-ray radiotherapy [37], during which both tissue and plaque magnetite are potentially exposed to the same dose. The overall number of magnetite-containing Aβ plaques in the AD brain, a limiting factor for PS treatment, is not known despite the numerous reports of iron/magnetite-containing Aβ plaques [9–12, 39]. Here, we estimated that more than 70%of Aβ plaques were magnetite-containing plaques by counting Aβ plaques and Fe foci following histochemical staining of hippocampal tissue, as shown in Table 1; this finding is comparable to that of a histochemical study in which Aβ plaques and iron were co-stained [21]. Given that ferrous iron-containing magnetite is formed by chemical reduction due to Aβ binding in either fibrils or plaques, the concomitant reduction in the number of Aβ plaques and levels of ferrous iron species in tissue sections as well as the conformational change of Aβ in PS-treated Aβ fibrils suggested local disruption of Aβ binding due to magnetite-derived electron transfer. This explanation is consistent with the dominant ferric magnetite after PS treatment of Aβ-magnetite fibrils, as reported previously [20]. The electron-transfer-mediated disruption of Aβ-magnetite bonding occurs very rapidly via picosecond-scale de-excitation (of ionized atoms [40] in magnetite before the emergence of ROS formation (μs∼ms scale), which curtails the damage caused to related protein aggregates in the surroundings. Changes in the oxidation state of magnetite occur upon dissociation from Aβ after PS, resulting in the potential uptake of unbound magnetite and degraded amyloid protein by brain microglia as an anti-inflammatory response [41–43]. Therefore, to investigate the oxidative conversion of iron and removal, histochemical analysis of iron rather than the time-consuming analysis of individual plaques by 30 nm-resolution STXM-XAS study was performed to assess the change in overall distribution. Multiple intraplaque distributions of iron foci were well delineated with apparent protein matrix in untreated AD brain as shown in Fig. 3C, whereas reduced number of iron foci were scattered without protein matrix in PS-treated AD brain. These results pertaining to plaque burden reduction indicated an irreversible change in the amyloid protein matrix, as suggested also by the results of IR spectroscopy and HPLC analysis. The irreversible degradation of plaque protein rather than the sonication-based transient disaggregation of insoluble Aβ plaques by PS would be a notable outcome [41]. The advantage of PS over multiple repeated sonications to achieve dissociation of plaques is also supported by the fact that it achieves simultaneous detoxification and removal of ferrous magnetite and Aβ plaques. This irreversible change suggests that reaggregation of Aβ or reformation of soAβs, is significantly inhibited, which, in turn, may lead to the effective diminution of Aβ-associated neurotoxicity and result in the enhancement of neurogenesis. However, the effect of PS on soAβs PS in the AD brain was not analyzed in this study. It is feasible that neurocognitive improvement after PS is ascribed to modulation of disturbed hippocampal neurogenesis [44, 45] and neuroinflammation upon reductions in both the number of Aβ plaques [46] and redox-toxic magnetite levels [43]. A decrease in ferrous iron levels indicates the transient emergence of soluble ferric iron by reoxidation of magnetite upon disruption of the Aβ-magnetite complex prior to elimination. This was evident from the ferric iron staining results, which demonstrated a latency to reduction due to the conversion of ferrous to ferric magnetite during the first 1-3 days of treatment. According to the results, PS, which decreases both the Aβ plaque burden and ferrous magnetite levels, effectively reduces sources of oxidative stress, neuroinflammation, and neuronal and synaptic loss [42, 46–48] and is a candidate therapeutic approach for AD.
Plaque regrowth is possible after treatment due to the partial removal of plaques and ferrous magnetite by single PS. In the cases of three PS-treated mice that showed improvement in the memory tests while demonstrating lack of improvement in the late phase, presently, we could not determine whether these findings resulted from short-lasting or delayed effects of single PS because we did not perform a series of studies on the same mouse. This as well as the mechanism by which PS improves cognition will be studied further. In early-onset AD mice, 2 + 2-Gy- or 1 + 1-Gy fractionated PS was observed to improve neurocognitive function without inducing cognitive deterioration. Further studies with repeated PSs are necessary to better evaluate dose schemes and the corresponding neurocognitive improvements in different AD stages.
The low LET plateau dose used in this study was less than 10 Gy of that required for the onset of radiation-induced cognitive dysfunction in normal tissues [49], as observed after typical oncological proton treatment (∼40 Gy) for glioma. Since even a 1-Gy proton dose induces a reduction in dendritic spine density in hippocampal tissue [50], it is necessary to monitor potential the long-term effect on cognitive impairment after PS. Owing to higher-energy protons, such as 300 MeV protons, which have a 50%lower Bragg-peak-to-plateau energy (BP/PD) ratio than 100 MeV protons, the effect of heavy ion stimulation with a higher BP/PD ratio at more than 300 MeV should be investigated and compared to the effect of PS at the same energy level as that required for transmitting the entire brain of the patient. The study underscores the potential of PS for the treatment of AD and other neurodegenerative diseases characterized by biomineralization of protein aggregates with iron oxide nanoparticles.