
Abstract
Background:
Tau hyperphosphorylation at several sites, including those close to its microtubule domain (MD), is considered a key pathogenic event in the development of tauopathies. Nevertheless, we recently demonstrated that at the very early disease stage, tau phosphorylation (pTau) at MD sites promotes neuroprotection by preventing seizure-like activity.
Objective:
To further support the notion that very early pTau is not detrimental, the present work evaluated the young rTg4510 mouse model of tauopathy as a case study. Thus, in mice at one month of age (PN30-35), we studied the increase of pTau within the hippocampal area as well as hippocampal and locomotor function.
Methods:
We used immunohistochemistry, T-maze, nesting test, novel object recognition test, open field arena, and electrophysiology.
Results:
Our results showed that the very young rTg4510 mouse model has no detectable changes in hippocampal dependent tasks, such as spontaneous alternation and nesting, or in locomotor activity. However, at this very early stage the hippocampal neurons from PN30-35 rTg4510 mice accumulate pTau protein and exhibit changes in hippocampal oscillatory activity. Moreover, we found a significant reduction in the somatic area of pTau positive pyramidal and granule neurons in the young rTg4510 mice. Despite this, improved memory and increased number of dendrites per cell in granule neurons was found.
Conclusion:
Altogether, this study provides new insights into the early pathogenesis of tauopathies and provides further evidence that pTau remodels hippocampal function and morphology.
INTRODUCTION
Neuronal protein inclusions are the hallmark of tauopathies [1]. Although the spectrum of tauopa-thies is vast (i.e., Alzheimer’s disease (AD), Pick disease, corticobasal degeneration, progressive supra-nuclear palsy, argyrophilic grain disease, chronic traumatic encephalopathy, and aging-related tau astrogliopathy), abnormally phosphorylated tau protein (pTau) aggregation remains as their common feature [1–3]. Tauopathies include complex syndromes such as neurofibrillary tangle dementia, frontotemporal dementia, corticobasal syndrome, Richardson syndrome, parkinsonism, akinesia with gait freezing syndrome, and cerebellar ataxia [1, 3]; however, the current consensus argues that tauo-pathies are a continuum of asymptomatic prodromal stages leading to symptomatic dementia stages [4]. One neuronal signature of the prodromal tauopathy stages is the disruption of neuronal activity and connectivity between brain regions [4, 5]. Recent studies performed in rTg4510 transgenic mice, which overexpresses the mutant human Tau-P301L gene and recapitulate the pathology with accumulation of neurofibrillary tangles (NFTs), showed that: 1) pTau aggregation in the entorhinal cortex is sufficient to disrupt the coordination of local field potentials between its efferent regions [6]; 2) pTau inclusions are correlated with early weakening of electroencephalogram theta oscillations and coherent activity between the prefrontal cortex and hippocampal CA1-CA3 networks [7]; 3) intracellular pTau aggregation results in theta-gamma oscillations uncoupling [7]; and 4) by changing the activity patterns of a subpopulation of affected neurons, pTau aggregation reduces the activity of the neocortical network [8]. In line with these findings, in 5-week-old triple transgenic AD mice (3xTgAD), we recently reported that pTau at sites S396 and T231, which are located close to the microtubule domain (MD) region, accumulates within hippocampal pyramidal neurons and some parvalbumin-positive (PV+) interneurons correlating with alterations in the hippocampal theta activity [9]. Considering that pTau disrupts ongoing network activity, all the data pointed to the deleterious roles of pTau. However, non-deleterious roles for pTau have also been reported [9–12]. For example, it was reported that pTau inhibited amyloid-β induced neuronal death and offered protection from excitotoxicity [12]. This finding coincides with our previous report that pTau protein at MD sites prevents N-methyl-D-aspartate receptor (NMDAR)-mediated overexcitation [10, 11]. Indeed, we demonstrated that pTau at MD sites counteracts overexcitation by facilitating long-term depression (LTD) [10, 13]. Long-term potentiation (LTP) and LTD provide a compelling cellular model for learning and memory [14–18]. We also reported a dramatic reduction in 4-aminopyridine-induced seizure-like activity in 5-week-old 3xTgAD mice [9, 19] and found that this reduction in the induction of epileptiform activity correlated with pTau’s MD accumulation in hippocampal pyramidal neurons and PV + interneurons [9]. Further suggesting a non-deleterious role for pTau, in 9-week-old Tau-P301L mice, increased LTP in the dentate gyrus was observed in parallel with improved cognitive performance [20]. Altogether, the data suggested that pTau plays a non-deleterious role during the early stage of disease.
In the present work, we sought to study the role of pTau during the early stages (5 weeks of age) of tauopathy-driven neurodegeneration in the young rTg4510 mice that express the human tau isoform P301L [21]. In addition, we evaluated the early effects of pTau accumulation on neuronal function by questioning whether pTau accumulation induces structural and functional hippocampal changes.
Our results showed that very young rTg4510 mice have no detectable changes in either hippocampal-dependent tasks or locomotor activity. Interestingly, the hippocampal pyramidal neurons and some PV + hippocampal interneurons accumulate pTau protein and this accumulation relates to changes in oscillatory activity recorded in the CA1/subiculum of very young rTg4510 mice. Importantly, enhanced learning and memory was associated with improved dendrites of granule neurons in the 5-week-old rTg4510 mice, despite a reduction in their somatic area.
METHODS
Animal strain
rTg4510 tetO-MAPT*P301L transgenic mice allow Tet-Off/Tet-On expression of a P301L mutant variant of human four-repeat microtubule-associated protein tau (Tau P301L), Jackson Labs. rTg4510 mice (male and female) between 30 and 35 days of age and age-matched non-transgenic (No-Tg) mice were housed in groups of 2–4 animals and maintained under a 12:12 h light cycle with water and food available ad libitum. Experimental procedures were approved by the Bioethics Committee of the Institute of Neurobiology (NOM- 062-ZOO-1999/UNAM). The genotype of each animal was confirmed by PCR (Jackson Labs. Protocol Standard PCR, at the INB Proteogenomic Unit).
Immunohistochemistry
Mice were anesthetized with pentobarbital and transcardially perfused with PBS followed by 4% PFA. Brains were stored in fixative for 24 h at 4°C, then in a sucrose solution (30% in PBS) for three days at 4°C. Next, the brains were frozen using dimethylbutane and stored at –80°C. Brains were sliced coronally at 40μm using a freezing microtome for light microscopy, or a vibrating microtome at 50μm for fluorescence microscopy, and free-floating sections were stored in a preservative solution (3:3:4 glycerol:ethylene glycol:PBS) at –20°C, in preparation for immunohistochemistry. Washes in PBS preceded all steps except primary antibody addition. All steps were performed at room temperature unless otherwise specified. For immunofluorescence, sections were incubated for 2 h with PBS containing 1% normal goat serum, 0.25% Triton X-100, and 0.45% gelatin. Sections were incubated overnight at 4°C with pS396 (Invitrogen), AT180 (Thermo Fisher), Tau-5 (Sigma-Aldrich), and anti-PV + (Synaptic Systems). Sections were incubated for 2 h with secondary antibody (Alexa 488–568 conjugated goat anti-rabbit at 1:1000 dilution, Invitrogen). Sections were then mounted on glass slides with Vectashield-DAPI Mounting Medium, Vector Labs. Fluorescence was visualized using an epifluorescence microscope (Axioplan 2, Zeiss). The hippocampal subfields were visualized with a 20X objective. A high magnification photomicrograph was obtained with a 40X objective.
Morphometry
Morphometric quantification in the areas was done on three microscopic fields from randomly chosen regions in the area of interest. Observations were conducted by bright field microscopy using a Zeiss microscope (Axioplan 2, Zeiss, Germany). Neuronal nuclei from the hippocampal region of interest were counted. Two to three slides were analyzed for each mouse. Cells in the region of interest were calculated per mouse and averaged per group. pTau-positive cells were identified and counted using 20x and 40x objective lenses. All cell counts were converted to a density value (pTau-positive cells/mm2) as previously described [2, 23]. For counting, and when critical observations to identify pTau-positive cells were necessary, low-magnification lenses were used. pTau-positive cells per square millimeter were registered from the CA1, CA2, CA3, DG, and subiculum. The fluorescent channel was imaged with an exposure time that was kept consistent across all slide imaging.
T maze
The T-maze task was designed to assess the cognitive ability of rodents in spontaneous alternation by allowing them to choose between two arms to explore a novel environment. This alternating choice depends on hippocampal functional integrity at the level of memory and learning [24–26]. For each trial, mice were placed into a T-shaped acrylic platform (40 cm long, 10 cm wide, and 15 cm high) located in a room with three space clues positioned in its walls. The T-maze consists of three basic zones: long arm (I) used as starting point, and two short arms for the alternative behavior (right and left). The long arm included a guillotine-like gate in its distal extreme used as the starting zone. The trial, which was repeated 3 times per day for 3 days, consisted of two phases: in the first phase, the mouse was placed in the start arm and allowed to choose one of the two short arms. The animal was left in the chosen arm for 20 s since the arm was blocked. For the second phase, the mouse was returned to the starting point and, once more, allowed to explore the maze and choose one of the two short arms. If the mouse did not choose an arm in either of the two phases of the task, the mouse was removed from the maze after 60 s and placed in its room box. This data was canceled from the analysis. The aim was to analyze whether the mouse would choose the same arm twice or alternate between arms. At the end of the study, the percentage of alternations between the novel and familiar arm was evaluated.
Nesting
The nesting test was performed to evaluate the general cognitive status of the animals by assessing the innate behavior of nest building [19, 27]. To do so, during the light period, 12 h prior to the night phase, each experimental subject was placed in an individual cage (26 cm×48 cm×20 cm) with a sawdust bed and a cotton square (5×5 cm). The quality of the nest made with the enrichment material (cotton) was evaluated according to the following scale: 0 = sawdust displacement by excavation, but intact cotton; 1 = sawdust movement to form a crater and minimal cotton shredding; 2 = clear sawdust crater formation and a substantial crumbling of the cotton piece leading to the formation of the nest with a concave shape; 3 = all the cotton material used for the construction of the nest, which has a spherical shape with a depth that completely covers the experimental subject.
Open field
The open field was used to assess exploratory behavior and locomotor activity. Every mouse was adapted to the room for 30 min and individually placed into the center of the open field (55 cm×32.5 cm), followed by a single 5 min session test, with the experimenter out of its view. During the open field test, a regulated light intensity of 30 lux was used. The floor was marked to visualize the separation between the peripheral and the central areas. A digital camera videotaped the whole field. The videos obtained from the tests were analyzed using automated tracking software (SMART, Panlab SL, Barcelona, España). The parameters included distance traveled, the time spent in each area (periphery and center), number of crosses and vertical exploration, which allowed to evaluate anxiety, exploratory behavior, and hyperactivity in the experimental subjects [28, 29].
Novel object recognition test
Mice were transferred to the testing room and acclimated for at least 2 h. Mice were tested in an open field. The open field was placed in a homogeneously illuminated room (30 lux at the level of the open field). On the first day, mice were habituated to the testing arena for 30 min. On the second day, two identical objects were placed in the same chamber for 10 min. Four hours later, the objects were replaced with one exact replica of the original object and a novel object of different shape and texture. The mice were then allowed to explore the objects and the chamber for an equal amount of time. Positions of the familiar and novel object in the chamber were changed semi-randomly between testing of different mice but were kept constant between training and test sessions for any given mouse. Frequency of interactions with the object and time spent exploring each object were recorded for subsequent analysis, and recognition memory was assessed at the final trial [30]. Arena and object were cleaned after testing of each mouse.
Morphology analysis
For reconstruction of individual neurons, immuno-fluorescence images were imported into Neurolucida neuron reconstruction software (MBF Bioscience, Williston, VT). The entire dendritic structure and soma of each neuron were manually traced. Reconstruction data was subsequently imported into NeuroExplorer software (MBF Bioscience). The enclosed area of each soma was determined. Number of dendrites per cell and total dendritic length were measured using a Scholl analysis, also in the NeuroExplorer software. Cell counting was done using the ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Electrophysiology
Animals were anesthetized with urethane (1.0 g / kg, i.p) and subsequently mounted on a stereotaxic apparatus for the placement of the recording electrode. The tip of the electrode reached the following coordinates (with respect to Bregma and dura): AP = –0.2 mm, ML =±2.0, mm and DV = 1.9 mm. Local field potentials were recorded using borosilicate electrodes filled with aCSF (2–6 MΩ). Signals were recorded with a differential AC amplifier (A-M Systems) and filtered (high pass 1 Hz, low pass 10 KHz). With custom MATLAB software, field traces were analyzed using Fourier transform. Power spectra were calculated using the multitaper method (Chronux toolbox, i.e., using seven tapers). Integrated delta, theta, beta, and gamma power was calculated in 5 s binds over a period of 2–10 min, and the mean spectrum was taken as the grand mean of each animal. Theta power was measured in μV2/Hz.
Data analysis
All data were compared statistically using GraphPad Prism 5 Windows, version 5.01 software. In all tests, values of p < 0.05 were considered significant unless otherwise specified. Bar graphs show experimental mean, with dispersion bars indicating standard error of the mean unless otherwise specified.
RESULTS
Hippocampal cells from young rTg4510 mice exhibit intracellular pTau accumulation
The immunohistochemical characterization of PN30-35 rTg4510 samples revealed the presence of intracellular pTau accumulation in neurons (Fig. 1). pTau (site S396) accumulation was detected in neurons from hippocampal CA1/CA2 (Fig. 1A, B), CA1/Subiculum (Fig. 1C), DG (Fig. 1D), and CA3 subfields (Fig. 1E). Of note, the number of pTau-positive neurons in PN30-35 rTg4510 mice was statistically increased when compared to PN30-35 No-Tg mice (No-Tg mean = 5.9±0.82 cells/mm2; rTg4510 mean = 36.20±4.9 cells/mm2, unpaired 2-tailed t test, t = 6.066, df = 10, p = 0.0001, No-Tg: n = 6; rTg4510: n = 6). Increases in pTau levels were clearly detected in somata (Fig.1A’, D’) and in the dendritic compartments of hippocampal cells (Fig. 1A’, arrow). The stratum pyramidale, which contains the hippocampal principal excitatory neurons, was revealed by high accumulation of nuclear 4’,6-diamidino-2-phenylindole (DAPI) labeling (Fig. 1B, F, blue-fluorescent stain). pTau accumulation was detected in principal neurons from the hippocampus (Fig. 1F, arrows). To further evaluate pTau at sites close to their MD, we evaluated the levels of pTau at sites T231-S235 labeled by AT180 (Fig. 1G, H, H’). The accumulation of AT180 was detected in cells from the PN30-35 rTg4510 samples (Fig. 1G, H, H’, see inserts). Some PV + hippocampal interneurons (Fig. 1I) accumulate pTau (Fig. 1J, see merge in Fig. 1K and arrows in Fig. 1L). Additionally, non-positive PV cells accumulate pTau (see asterisk in Fig. 1K, L). Detailed analysis of pTau-positive PV + cells in PN30-35 rTg4510 mice revealed diffuse pTau-positive staining within the cytoplasm, sometimes comprising small punctate regions (Fig. 1L, see insert). Around 50% of the cells detected by total tau (Fig. 1M) accumulated pTau (Fig. 1N, see arrows in Fig. 1O). Some cells detected by total tau antibody did not accumulate pTau (see asterisk in Fig. 1M-O). Importantly, the density of nuclei in CA1 and subiculum was statistically similar between groups (CA1No - Tg versus CA1rTg4510 unpaired 2-tailed t test, t = 0.06355, df = 10, p = 0.9506; Mann–Whitney U = 17, p = 0.9372, SubNo - Tg versus SubrTg4510 unpaired 2-tailed t test, t = 1.932, df = 10, p = 0.0821; Mann–Whitney U = 8, p = 0.132, Fig. 1P). When we analyzed the number of pTau-positive neurons, we found that pTau cells per mm2 were statistically similar between hippocampal areas (CA1 mean = 36.2±13.2 cells/mm2; CA2 mean = 36.37±12.7, CA3 mean = 29.39±4.84, DG mean = 29.74±22.56; CA1 versus CA2 unpaired 2-tailed t test, t = 0.0258, df = 10, p = 0.9799; Mann–Whitney U = 17, p = 0.9372, CA1 versus CA3 unpaired 2-tailed t test, t = 1.161, df = 10, p = 0.2726; Mann–Whitney U = 10, p = 0.2403, CA1 versus DG unpaired 2-tailed t test, t = 0.6702, df = 10, p = 0.5179; Mann–Whitney U = 13.5, p = 0.5211, Fig. 1Q). A planned comparison between Subiculum and CA1, CA2, and CA3 areas revealed lower levels of pTau-positive neurons per mm2 in the hippocampus (Sub mean = 19.78±5.72 cells/mm2, Sub versus CA1 unpaired 2-tailed t test, t = 2.957, df = 10, p = 0.0144; Mann–Whitney U = 4, p = 0.026, Sub versus CA2 unpaired 2-tailed t test, t = 3.015, df = 10, p = 0.013; Mann–Whitney U = 3.5, p = 0.0247, Sub versus CA3 unpaired 2-tailed t test, t = 2.344, df = 10, p = 0.0411; Mann–Whitney U = 5, p = 0.0446, Sub versus DG unpaired 2-tailed t test, t = 1.182, df = 10, p = 0.2644; Mann–Whitney U = 15, p = 0.6879, Fig. 1Q).
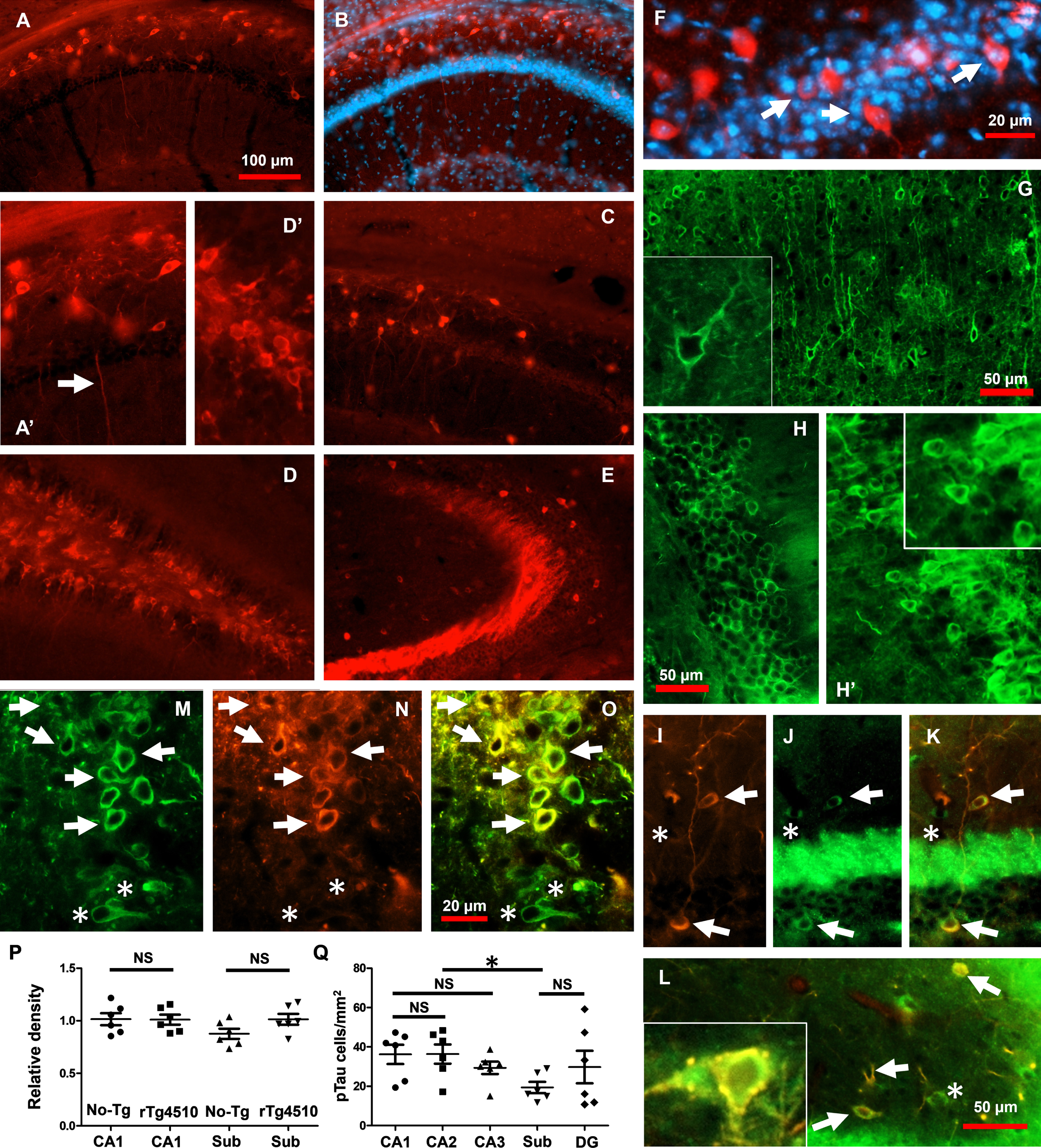
Intracellular pTau into hippocampal cells in very young rTg4510 mice. pTau protein, detected through anti-S396 antibody, was found accumulating within pyramidal cells from the hippocampal CA1/CA2 area in the rTg4510 group (A, B). pTau accumulation was detected also within cells from CA1/Sub (C), DG (D), and CA3 in the rTg4510 group (E). Early pTau accumulation was detected in both dendrites (A’, arrow) and somata (D’). pTau protein was clearly observed in the stratum pyramidale as revealed by DAPI (B, F). pTau at sites labeled by AT180 was found coexisting with neurons from cortex (G, see insert), and hippocampus (H and H’, see insert). Hippocampal interneurons (I) accumulate pTau (I-L, arrows). AT180-positive PV + interneurons revealed punctate accumulation of pTau (L, see insert). Cells detected by total Tau (M) accumulate pTau (N, see merge in O). Density of nuclei in CA1 and subiculum was statistically similar between No-Tg and rTg4510 groups (P). Quantification of pTau-positive cells per mm2 showed that almost all the hippocampal areas from young rTg510 exhibited the same number of pTau-positive cells per mm2 (Q). The subiculum showed that the number of cells exhibiting pTau was significantly smaller than those exhibiting the other hippocampal areas (Q). The graph shows experimental mean and standard error of the mean, * denotes a significant difference between groups p < 0.05.
Overall, and consistent with our previous findings [9], we showed that pTau at MD sites increases at very initial stages of tauopathy progression. Additionally, pTau at MD sites is expressed in different neuronal types in the entire hippocampus of the very young rTg4510 mice.
Working memory and hippocampal dependent function
Spatial working memory was analyzed through spontaneous alternation in the hippocampus-dependent T-maze [24–26]. In this test, control mice showed a choice accuracy of 71.30±5.54% (Fig. 2A), whereas the very young rTg4510 achieved a choice accuracy of 60.10±7.49% (Fig. 2A). The statistical comparison revealed no significant differences between genotypes (unpaired 2-tailed t test, t = 1.202, df = 18, p = 0.2449; Mann–Whitney U = 36.5, p = 0.3174, No-Tg: n = 10; rTg4510: n = 10, Fig. 2A). To further explore hippocampal-dependent function, we performed the innate nesting test [9, 27]. Innate nesting is significantly reduced upon hippocampal cytotoxic lesions and can be evaluated in animals at early post-natal developmental ages [9, 27]. Consistent with the T-maze test, the statistical comparison of nesting behavior revealed no significant differences between genotypes (No-Tg =2.05±0.21 nesting rating and rTg4510 = 1.65±0.15 nesting rating; unpaired 2-tailed t test, t = 1.518, df = 18, p = 0.1464; Mann–Whitney U = 31, p =0.1466, No-Tg: n = 10; rTg4510: n = 10, Fig. 2B). To identify memory deficits, we tested mice in the novel object recognition test [30]. Surprisingly, PN30-35-day-old rTg4510 mice spent significantly more time with the novel object than with the familiar one (unpaired 2-tailed t test, t = 3.234, df = 16, p = 0.0052; Mann–Whitney U = 9, p = 0.0044, No-Tg: n = 8; rTg4510: n = 10, Fig. 2C).
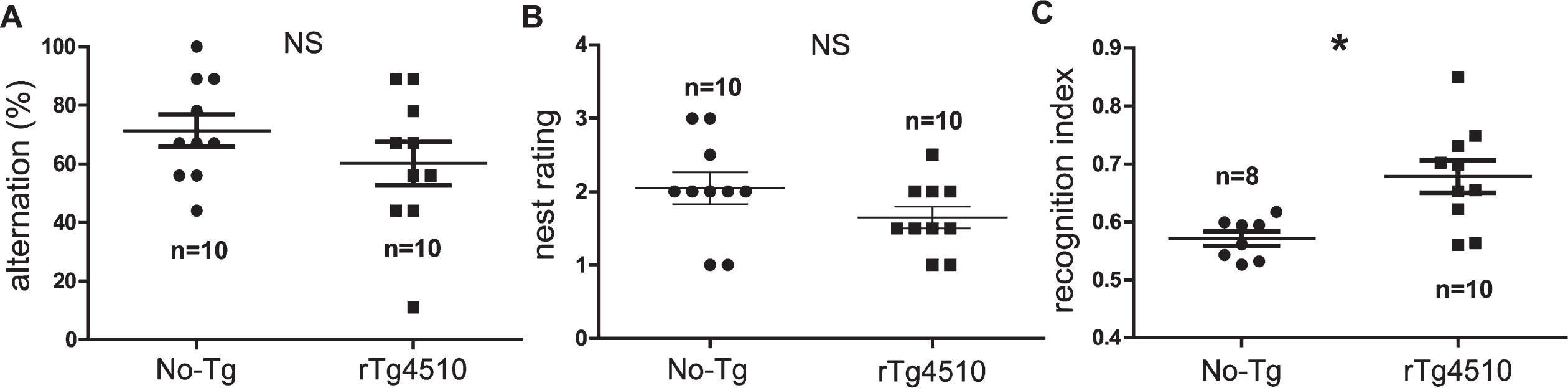
Working memory and nesting are not affected in the young rTg4510 mice. Alternation in the T-maze (working memory) showed no significant differences between No-Tg and rTg4510 mice (A). The nest rating revealed no significant differences between genotypes (B). rTg4510 mice spent significantly more time with the novel object than with the familiar one (C). All graphs show experimental mean and standard error of the mean. NS denotes no significant difference between groups p > 0.05 and * denotes a significant difference between groups p < 0.05.
In sum, the young rTg4510 mice showed no signs of hippocampal function deficit at the very early developmental age of PN30-35. Conversely, PN30-35-day-old rTg4510 mice displayed improved memory.
Motor activity and exploration in the young rTg4510 mice
Motor activity and exploration were assessed in the open-field arena [28, 29]. The PN30-35 rTg4510 mice were similarly active to age-matched No-Tg mice (Fig. 3A, B, mouse trajectories: black lines). During the 5 min of open-field spontaneous exploration, the total distance traveled showed no significant differences between No-Tg and rTg4510 mice (No-Tg = 2250 ± 125.1 and rTg4510 = 2314±157.4 cm, unpaired 2-tailed t test, t = 0.06893, df = 22, p = 0.9457; Mann–Whitney U = 71, p = 0.9770, No-Tg: n = 10; rTg4510: n = 10, Fig. 3C). The total number of crosses into the center area of the open-field arena showed no significant difference between No-Tg and rTg4510 (No-Tg = 54.30±4.81 and rTg4510 = 60.20±3.83; unpaired 2-tailed t test, t = 0.9580, df = 18, p = 0.3508; Mann–Whitney U = 36.5, p = 0.3254, No-Tg: n = 10; rTg4510: n = 10, Fig. 3D). However, the rTg4510 mice spent significantly less time in the center of the arena when compared to family age-matched No-Tg mice (No-Tg = 128.5±4.97 sec and rTg4510 = 104.5±9.89 s; unpaired 2-tailed t test, t = 2.167, df = 18, p = 0.0439, Mann–Whitney U = 28, p = 0.1051, No-Tg: n = 10; rTg4510: n = 10, Fig. 3E). Finally, regarding rearing display movements, both genotypes showed no significant differences in the number of rearing (No-Tg = 192.3±2.31 and rTg4510 = 189.6±3.00; unpaired 2-tailed t test, t = 0.7296, df = 15, p = 0.4769; Mann–Whitney U = 27.5, p = 0.4932, No-Tg: n = 10; rTg4510: n = 7, Fig. 3F).
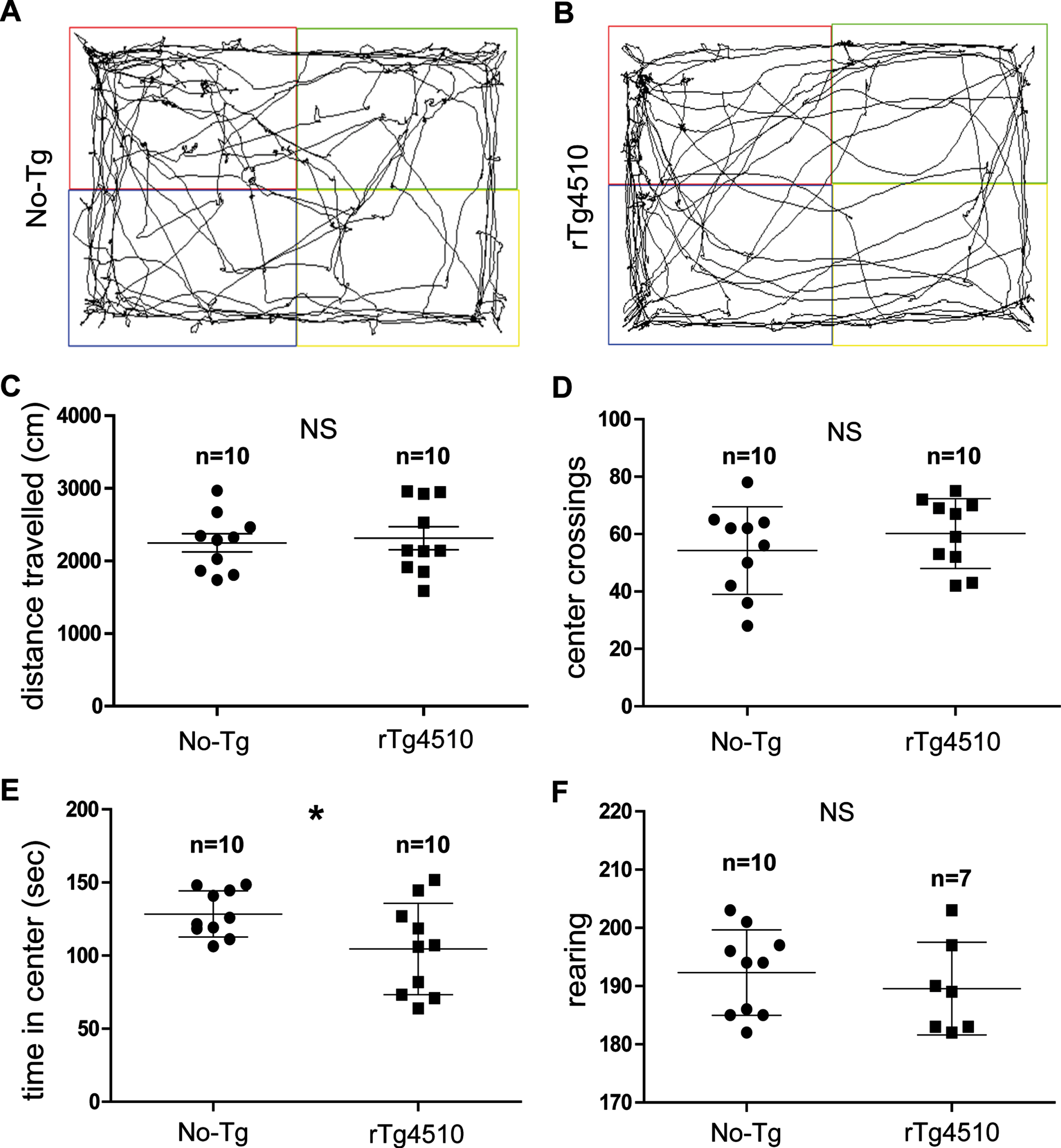
Exploration of motor activity showed no alterations in the young rTg4510 mice. Representative trajectories of No-Tg (A) and rTg4510 mice (B). The total distance traveled in the open field arena showed no significant changes between No-Tg and rTg4510 groups (C). The total number of squares crossed in the open field arena showed no significant difference between No-Tg and rTg4510 mice (D). In contrast, the rTg4510 spent significantly less time in the center when compared to No-Tg mice (E). Rearing display showed no significant differences between No-Tg and rTg4510 groups (F). All graphs show experimental mean and standard error of the mean, NS denotes no significant difference between groups p > 0.05 and * denotes a significant difference between groups p < 0.05.
Although the very young rTg4510 mice spent less time in the center of the open-field arena, their motor activity and motor coordination showed no alterations.
Hippocampal morphology in the young rTg4510 mice
To evaluate potential morphological changes, we investigated structural characteristics in several hippocampal subfields of rTg4510 mice in direct comparison with No-Tg mice of the same age. We focused the structural analysis on cells characterized by pTau accumulation (Fig. 4). Parameters such as the area of individual soma and number of dendritic ramifications were evaluated (Fig. 4). The somatic area (μm2) of the pyramidal cells was significantly decreased in the PN30-35 rTg4510 mice (No-Tg = 14.29±3.47% and rTg4510 = 6.66±0.98%; unpaired 2-tailed t test, t = 2.795, df = 20, p = 0.0112; Mann–Whitney U = 23, p = 0.0409, No-Tg: n = 7; rTg4510: n = 15, Fig. 4A). In accordance with the findings in the pyramidal layer, the somatic area of DG cells was significantly smaller in the PN30-35 rTg4510 mice (No-Tg = 20.00±4.14% and rTg4510 = 11.11±0.80%; unpaired 2-tailed t test, t = 2.790, df = 12, p = 0.0163; Mann–Whitney U = 9, p = 0.0829, No-Tg: n = 7; rTg4510: n = 9, Fig. 4B). Surprisingly, dendrites per cell in the DG cell layer (Fig. 4C) were significantly higher in the PN30-35 rTg4510 mice (No-Tg = 1.8±0.2 and rTg4510 = 2.66±0.24; unpaired 2-tailed t test, t = 2.750, df = 26, p = 0.0107; Mann–Whitney U = 26.5, p = 0.0003, No-Tg: n = 16; rTg4510: n = 12, Fig. 4D).
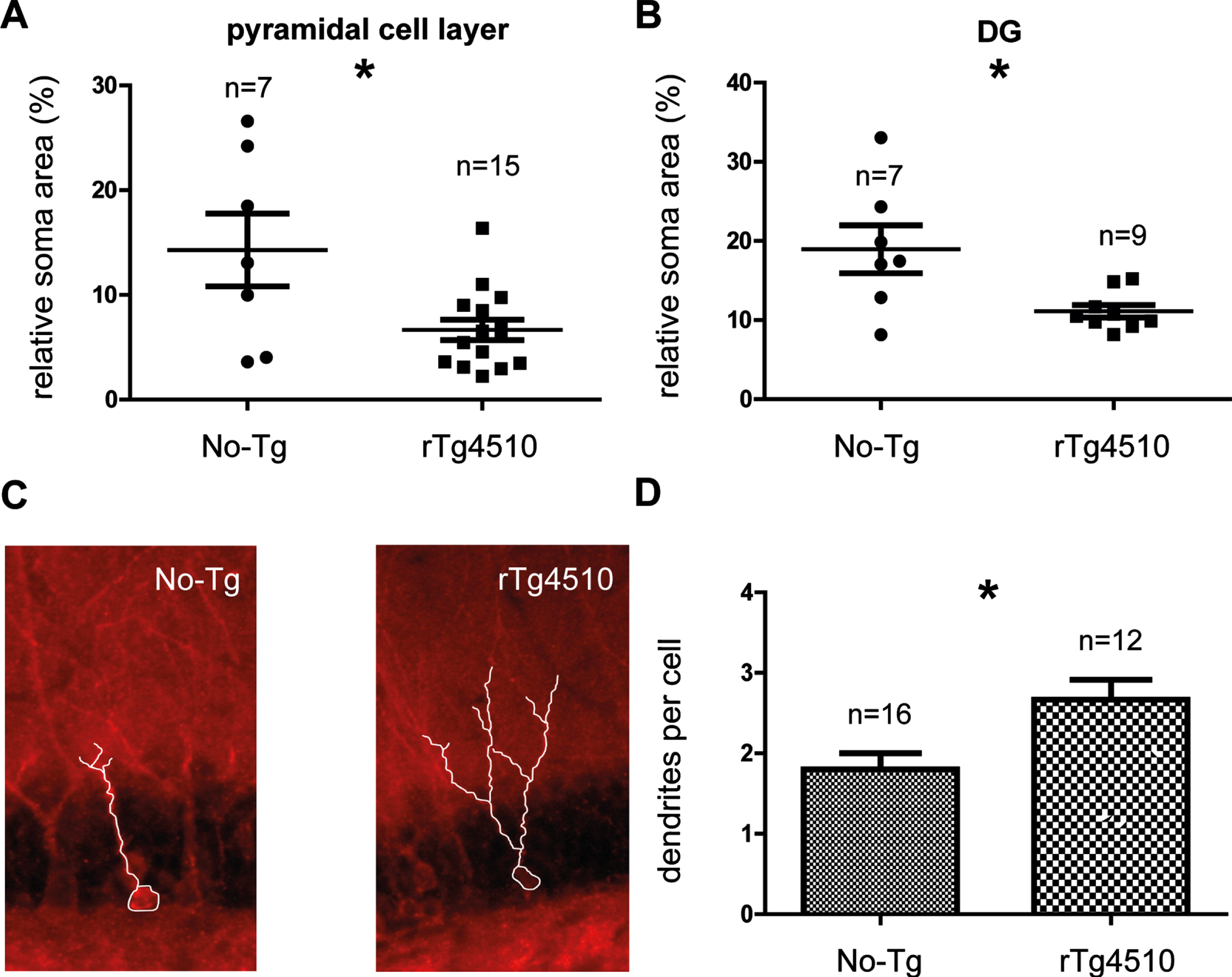
Hippocampal morphology in the young rTg4510 mice. The somatic area of the pyramidal cell layer (A) and DG layer (B) were significantly decreased in the young rTg4510 mice. Examples of cells in the DG are shown (C). The dendrites per cell in the DG layer were significantly increased in the rTg4510 mice (D). All graphs show experimental mean and standard error of the mean, * denotes a significant difference between groups p < 0.05.
Although the area of individual soma was de-creased in the PN30-35 rTg4510 mice, the dendrites per cell in the DG were significantly increased in the PN30-35 rTg4510 mice.
Changes in hippocampal network activity in young rTg4510 mice
Our previous findings have suggested that very early accumulation of intracellular pTau in hippocampal neurons could have an impact on their network activity [9, 10]. To further evaluate this possibility, we studied the intrinsic spontaneously ongoing oscillatory activity in the hippocampus of the PN30-35-days-old rTg4510 mice in vivo (Fig. 5A). In line with our previous report [9, 31], population activity from No-Tg animals showed rhythmic theta activity in the hippocampal area (Fig. 5B). Filtered traces further confirmed the presence of rhythmic hippocampal network activity at theta band-frequency (3–10 Hz) and gamma-band frequencies (> 25 Hz) (Fig. 5B, lower traces). Similarly, hippocampal network activity recorded from the young rTg4510 mice displayed rhythmic activity at theta-band frequency (3–10 Hz) and gamma band-frequencies (> 25 Hz) (Fig. 5C, see lower traces). The power spectrum analysis revealed a stable power theta band activity with a mean frequency of 8.41±0.52 Hz in the No-Tg group (Fig. 5D) and 7.13±0.16 Hz in the young rTg4510 mice (Fig. 5D). Statistical comparison between groups revealed a significant decrease in hippocampal theta frequency in the young rTg4510 mice when compared to the family-related No-Tg mice (unpaired 2-tailed t test, t = 2.670, df = 15, p = 0.0175; Mann–Whitney U = 9, p = 0.0097, No-Tg: n = 7; rTg4510: n = 10, Fig. 5D). Regarding the relative power of the oscillatory activity, no significant differences were observed in the relative power at delta band (1–3 Hz) between groups (No-Tg = 25.51±2.38% and rTg4510 = 21.11±3.52%; unpaired 2-tailed t test, t = 1.07, df = 16, p = 0.3004; Mann–Whitney U = 28, p = 0.3154, No-Tg: n = 10; rTg4510: n = 8, Fig. 5E). However, a significant reduction was observed in the relative theta power in young rTg4510 animals when compared to the No-Tg group (No-Tg: 43.39±1.31% and rTg4510:38.65±1.21%; unpaired 2-tailed t test, t = 2.594, df = 16, p = 0.0196; Mann–Whitney U = 13, p = 0.0155, No-Tg: n = 10; rTg4510: n = 8, Fig. 5F). Moreover, statistical analysis of young rTg4510 exhibited a significant increase in slow and fast gamma power (slow gamma (25–45 Hz): No-Tg = 4.51±0.58% and rTg4510 = 7.37±1.35%; unpaired 2-tailed t test, t = 2.077, df = 16, p = 0.0542; Mann–Whitney U = 21, p = 0.1011, No-Tg: n = 10; rTg4510: n = 8, Fig. 5G, and fast gamma (45–250 Hz): No-Tg = 0.83±0.12% and rTg4510 = 2.35±0.54%; unpaired 2-tailed t test, t = 3.036, df = 16, p = 0.0079; Mann–Whitney U = 5, p = 0.0009, No-Tg: n = 10; rTg4510: n = 8, Fig. 5H).

Oscillatory activity is altered in the hippocampus of very young rTg4510 mice. In vivo recording site within the hippocampus (A). Representative traces of spontaneous field activity recorded in the hippocampus of No-Tg (B, upper trace) and rTg4510 mice (C, upper trace). Same recordings are presented after being filtered for the theta band frequency (middle traces) and gamma band frequency (lower traces). The power spectrum analysis revealed significant decrease in hippocampal theta frequency in the young rTg4510 mice when compared to the family-related No-Tg mice (D). The power spectrum analysis showed no significant changes in the relative power of the delta band (1–3 Hz) between No-Tg and rTg4510 groups (E). Power spectrum analysis of the theta band showed a significant reduction in the relative power (F), and a significant increase in the relative power of the slow gamma band (G) and fast gamma band (H) of the rTg4510 animals. All graphs show experimental mean and standard error of the mean, NS denotes no significant difference between groups p > 0.05 and * denotes a significant difference between groups p < 0.05.
Summarizing, the electrophysiological characterization demonstrated changes in the population rhythmic activity, at the theta and gamma frequency bands, in the CA1/subiculum of PN30-35 rTg4510 mice.
DISCUSSION
In this study, we assessed the impact of early pTau accumulation on neuronal function in the hippocampus. First, we showed that in rTg4510 mice pTau (sites S396/T231/S235) accumulates at an early age (PN30-35) in different neurons from the hippocampus including principal cells and PV + interneurons (Fig. 1). In line with our findings, hyperphosphorylated tau at sites T231/S235 (phospho-epitope AT180), was observed in the CA1 pyramidal neuron layer of rTg4510 mice at one and two months of age [32]. Besides, hyperphosphorylated tau at sites S400, S404, and S412 was reported in rTg4510 mice at 6 weeks of age [33]. Importantly, pTau accumulation was observed prior to changes in hippocampal density (Fig. 1), indicating that tau hyperphosphorylation occurs before cell loss. Supporting these findings, neuronal loss in rTg4510 mice was reported after five months of age [34]. Conversely, hyperphosphorylated tau at sites S199/S202/T205 (phospho-epitope AT8) was not present in rTg4510 mice at 6 weeks of age [33]. Altogether, data in young rTg4510 mice showed that hyperphosphorylation of tau protein at the carboxyl terminus (tau’s MD region) is among the earliest events, and it occurs before the classical fibrillar structure (NFTs) appears. Similarly, this sequence of hyperphosphorylation events was observed in patients with AD and related disorders [2, 3]. Of direct relevance, these disorders are characterized by synaptic dysfunction in the early phases of pathogenesis [35, 36]. In the same regard, several studies reported age-dependent synaptic alterations in rTg4510 mice [37–40]. Mechanistically, increases of phosphorylation in sites near tau’s MD reduced tau binding ability, therefore perturbing the synaptic and behavior response [10, 41–43]. Nonetheless, we showed that this early neuronal pTau expression and accumulation does not impair hippocampal and motor behavior in young rTg4510 mice (Figs. 2 3). On the contrary, young rTg4510 mice showed a significant improvement in cognitive performance in the object recognition task (Fig. 2). In support of our findings, spatial memory deficits in rTg4510 mice were first detected at 4 months of age [44]. Accordingly, it was demonstrated that the P301L mice, an rTg4510 homologous model, did not suffer either cognitive or motor deficits at the early age of 10 weeks [20]. Further, P301L mice showed increased LTP in the DG correlated with improved cognitive performance [20]. This improved cognitive performance in early stages of the P301L mice model was related to the absence of marked pTau accumulation and the absence of morphological changes [20]. In contrast, our data revealed steady pTau accumulation associated with the enhanced cognitive performance and absence of motor deficits in young rTg4510 mice. These findings are in line with the recently reported steady accumulation of pTau related to the absence of cognitive impairment in the 5-week-old 3xTg-AD model [9].
These new findings in the young rTg4510 model, along with the well-characterized cognitive and motor deficits starting at 4 months of age [44–46], support the non-deleterious role for pTau during the early stage of disease.
In normal neuronal process tau is involved in the formation of long cytoplasmic extensions that will become the axon and the dendrites [47, 48]. Tau facilitates tubulin assembly, stabilizes polymerized microtubules, and nucleates microtubules [49]. Since the tau binding site on tubulin overlaps with that for other proteins like the molecular motor kinesin, tau also influences processes like axonal transport [50]. In sum, tau protein participates in maintaining the neuron’s shape and the neural transmission [48]. Additionally, tau can bind to proteins such as spectrin, actin, phosphatases like PP1 and PP2A, kinases like CDK5, GSK3 and Fyn tyrosine kinase, and synaptic proteins like postsynaptic density protein-95 (PSD-95) [10, 51–53]. Tau also participates in the most studied cellular model of memory, LTP and LTD [10, 54]. Of relevance, tau phosphorylation is the predominant way in which tau function can be regulated [10, 55]. For example, tau phosphorylation at Thr231 and Ser262 affects microtubule binding [53]. In the same regard, tau phosphorylation at sites labeled by PHF-1 and AT180 affects the interaction between tau and Fyn and between tau and PSD-95 [10, 43]. Thus, the phosphorylation of tau protein also influences the distribution of this protein [56]. For instance, tau phosphorylation in its carboxy-terminal domain is mainly found in the distal axonal region and dendrites [56]. Additionally, hyperphosphorylated tau has been described in extracts of both normal and AD tissue [2, 58]. Altogether, current data along with the data reported in this study support the non-deleterious role for hyperphosphorylated tau during the early stage of disease.
Regarding the dominant tau hypothesis, pTau is critical for cognitive decline in tauopathies [1, 41–43]. Tau becomes abnormally phosphorylated, dissociates from microtubules, aggregates into NFTs, and translocates from the axon to the dendritic spines, causing synaptic and network dysfunction [8, 43]. Pathologically, the progressive increase and accumulation of pTau have been associated with neuronal atrophy, proposed to be a result of synaptic degradation and neuronal loss [34, 59]. Mechanistically, it was demonstrated that pTau accumulates in the somatodendritic compartment of neurons and has a dendritic function in the postsynaptic targeting of the Src kinase Fyn [43, 60]. Phosphorylation of the NMDAR by Fyn promotes its interaction with the scaffolding PSD-95, leading to excitotoxic downstream signaling and synaptic alterations [43].
Aiming to address the potential morphological correlates of the increased pTau in the hippocampus of the young rTg4510 mice, we evaluated structural changes in hippocampal subfields (Fig. 4). Despite the reduction in area of individual soma in the hippocampus, no dendritic pathology was detected (Fig. 4). Indeed, an increased number of dendrites per cell in granule neurons was found (Fig. 4). Contrary to the dominant tau hypothesis, an increased number of dendrites and significant improvement in cognitive performance support the non-deleterious role for hyperphosphorylated tau during the early stage of disease in the young rTg4510 mice.
Considering that dendrites are complex structures consisting of a dense network of cytoskeletal, transmembrane and scaffolding molecules, and surface receptors [61], this last observation suggests an interesting role for pTau in dendritic morphology. Giving support to this hypothesis, it was reported that the PACSIN1-family of cytoplasmic adapter protein is a tau binding partner in regulating microtubule dynamics and forming axonal plasticity [62]. Interestingly, PCASIN1 blockade results in impaired axonal elongation and a higher number of primary axonal branches in mouse dorsal root ganglia neurons, which is induced by reduced tau microtubule binding ability and accumulation in the central domain of growth cones [62]. As previously mentioned, is widely known that tau’s affinity for microtubules is actively regulated by phosphorylation [41, 42]. Thus, an early increase in phosphorylation levels of dendritic tau disrupts the interaction between tau and dendritic proteins [10, 41–43]. Similarly, we believe that the higher number of dendritic branches in young rTg4510 mice could serve as a compensatory mechanism exerted by pTau during early stages of disease progression. This observation contributes to the hypothesis that pTau protein during early stages of disease progression could be involved in neuroprotection and neural function improvement, rather than being exclusively involved in neurodegeneration [9–12, 20].
In sum, the current data demonstrated that pTau increases are not correlated with cognitive or motor impairment, or with major morphological alterations. In contrast, pTau increases were correlated with a rise in the number of dendrites in DG cells and improved cognition. Therefore, tau mutation and early pTau accumulation per se are not critical for cognitive and motor decline.
Non-fibrillar pTau and neuronal activity in the young rTg4510 mice
Despite the absence of cognitive and locomotor alterations in early AD development, we have previously reported changes in the hippocampal network (oscillatory activity) of 5-week-old J20 and 3xTgAD mice [9, 63]. The two transgenic mouse models are characterized by accumulation of pTau at MD sites during early stages of disease [9, 64].
Aiming to evaluate ongoing activity in the young anesthetized rTg4510 mice, we assessed their hippocampal oscillatory activity (Fig. 5). During slow-wave sleep (SWS) and under anesthesia, the membrane potential spontaneously transitions between the hyperpolarized silent down state and the depolarized-up state, from which action potential arises [8, 65]. Therefore, anesthesia serves as a model for SWS that allows the evaluation of spontaneous ongoing activity of the neurons within an intact network [8, 66].
Here we showed that hippocampal ongoing activity recorded in the CA1/subiculum from PN30-35 rTg4510 mice was altered before any cognitive and locomotor deficits (Fig. 5). Changes include slower frequency oscillations in the theta band, reduced power in the lower frequency bands (theta), and increased power in the gamma band (Fig. 5). It is worth mentioning that electrophysiological studies in rTg4510 mice have mostly been performed in mice over 7 months old [67]. Regardless, at 7 to 8 months of age, when rTg4510 mice exhibit advanced pTau deposition (NFTs) and progressive neurodegeneration, theta band recorded in the hippocampal CA1 was also decreased [68]. Hypothesizing that pTau aggregates (NFTs) are the seminal event for alterations in network oscillations, the inoculation of fibrillar aggregates of human tau fragments containing the four-microtubule-binding repeat domains in 3-month-old P301L mice reduced the power of the hippocampal CA1 theta and delta bands [7]. In agreement, by expressing P301L human tau in adult rats, theta activity showed significantly lower amplitude values when directly compared to the control, while gamma activity showed higher amplitude values [6], which is what we found here.
Overall, slower frequency oscillations and reduced power in the theta band, along with pTau accumulation in principal cells and PV + interneurons suggested that early stages of tauopathy were mainly characterized by a reduction in hippocampal excitability. In this regard, our previous work demonstrated that increases in pTau in principal cells and PV+interneurons reduced hippocampal excitability in 5-week-old 3xTgAD mice [9, 19]. Thus, we demonstrated that increases in pTau at MD sites prevent seizure activity in 5-week-old 3xTgAD mice [9, 19]. In addition, we reported that increases in sites T231, S235, S396, and S404 reduced the susceptibility to excitotoxicity by perturbing the PSD-95–tau–Fyn synaptic complex, therefore promoting the synaptic response to LTD [10, 11]. A patch-clamp of hippocampal CA1 neurons from rTg4510 mice showed a more depolarized threshold for action potential generation and a reduction in firing, further supporting the role of pTau in reducing neuronal excitability [32]. Besides, reduced neuronal excitability results from the relocation of the axon initial segment down the axon in a pTau-dependent manner [32]. We therefore propose that a reduction in hippocampal excitability caused by an increase in pTau contributes to the remodeling in hippocampal activity.
Aside from the control of neuron excitability, an alternative hypothesis suggests that regardless of whether the neurons bear NFTs, non-fibrillar pTau presence is sufficient to alter neuronal activity [8, 9]. In this regard, it was demonstrated that non-fibrillar pTau reduces the activity of single neocortical pyramidal cells and the neocortical network in 3- to 5-month-old rTg4510 mice [8]. Thus, it is possible that intracellular pTau affecting only a fraction of the neurons escalates to a disruption of the entire neuronal network [8]. Supporting this hypothesis, we showed that non-fibrillar pTau accumulates in pyramidal neurons and in some PV+interneurons from one-month-old 3xTg-AD mice [9]. Additionally, intracellular pTau affecting a fraction of the neurons correlated with changes in oscillatory activity [9]. Because the harmonious activity between pyramidal neurons and PV + interneurons is mainly responsible for the generation and maintenance of oscillatory rhythms [69], we proposed that early non-fibrillar pTau accumulation in young rTg4510 mice is greatly responsible for the electrophysiological alterations describe here. Thus, these early alterations preceded cognitive and motor deficits, defining the prodromal stage of tauopathies. Additionally, we believe that the changes recorded in the young rTg4510 mice share a similar mechanism to that of young 3xTgAD mice and young J20 mice, in which pTau affects a fraction of pyramidal cells and PV + interneurons [9, 31].
Overall, the findings in this report showed that: 1) Tau mutation and early non-fibrillar pTau accumulation per se are not critical for cognitive and motor decline, 2) the presence of early non-fibrillar pTau accumulation correlated with improved cognition and improved dendritic morphology in DG cells, suggesting a positive role for pTau, at least during very early stages of tauopathy development, 3) intracellular pTau affecting only a fraction of the neurons might escalate to a disruption of the entire neuronal network, and 4) the recorded electrophysiological changes become a potential biomarker for detecting the prodromal stage of tauopathies.
Footnotes
ACKNOWLEDGMENTS
We thank Jessica Gonzalez Norris for proofreading. We thank the following facilities from INB: Proteogenomic Facility Unit (Adriana Gallardo), Microscopy Facility Unit (Nydia Hernández-Rios), Behavioral Analysis Core Facility (Deisy Gasca), and Vivarium Facility (Alejandra Castilla and Martín García). We also thank Benito Ordaz-Sánchez and Azucena Aguilar-Vázquez for technical support.
This research was supported by Consejo Nacional de Ciencia y Tecnología CONACYT (grant numbers 269021, 319863 and A1-S-7540) and by Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México UNAM, México (grant number AG200521). S M-R was awarded a Cátedra position by CONACYT, México.