
Abstract
Background:
Alzheimer’s disease (AD) characterized by neurofibrillary tangles caused by hyperphosphorylated tau is the most common cause of dementia. Zeaxanthin (Zea), derived from fruits and vegetables, may reduce the risk of AD. Endoplasmic reticulum stress (ERS) might cause memory impairment in AD.
Objective:
Here, we studied protective role of Zea on the relationship among ERS, activity of glycogen synthase kinase 3β (GSK-3β, tau phosphorylated kinase), and p-Tau (Ser 396 and Thr 231).
Methods:
The results were obtained in non-RA and RA group by using different treatment, such as 9-cis-retinoic acid (RA), TM (ERS inducer), Zea, 4-PBA (ERS inhibitor), and SB216763 (GSK-3β inhibitor). The methods included flow cytometry and MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] for the detections of cell cycle and cell viability and western blot as a third measure of proteins in relation to ERS and tau phosphorylation. We have collected and analyzed all the data that suggested application of drugs for the treatment in non-RA and RA group.
Results:
Zea displays its protection on TM-induced cell injury, upregulation of GRP78 expression, and change of GSK-3β activity and tau phosphorylation when 4-PBA and SB216763 interfere with the process.
Conclusion:
These studies indicated that Zea is in vicious circle in ERS, GSK-3β, and tau phosphorylation, and further reflect its potential value in AD.
INTRODUCTION
Alzheimer’s disease (AD) is related to a particular neuropathology and results in cognitive decline \nobreak connected with age and with a particular onset [1]. This neurodegenerative disease is classically characterized by two hallmark pathologies: amyloid-β plaque deposition and hyperphosphorylated tau-caused neurofibrillary tangles [2]. Trajectory of deaths from AD was likely exacerbated in 2020 because of the COVID-19 pandemic and total payments in 2021 for hospice services, health care, and long-term care for people who age 65 and older with dementia are \nobreak evaluated to reach $355 billion [3]. On the one hand, highly promising drugs targeting amyloid- β recently failed to show clinical benefits in phase III trials, and on the other hand, most anti-tau agents in clinical trials involved in immunotherapies are only in the early stages of clinical research when some drugs reached phase II [4].
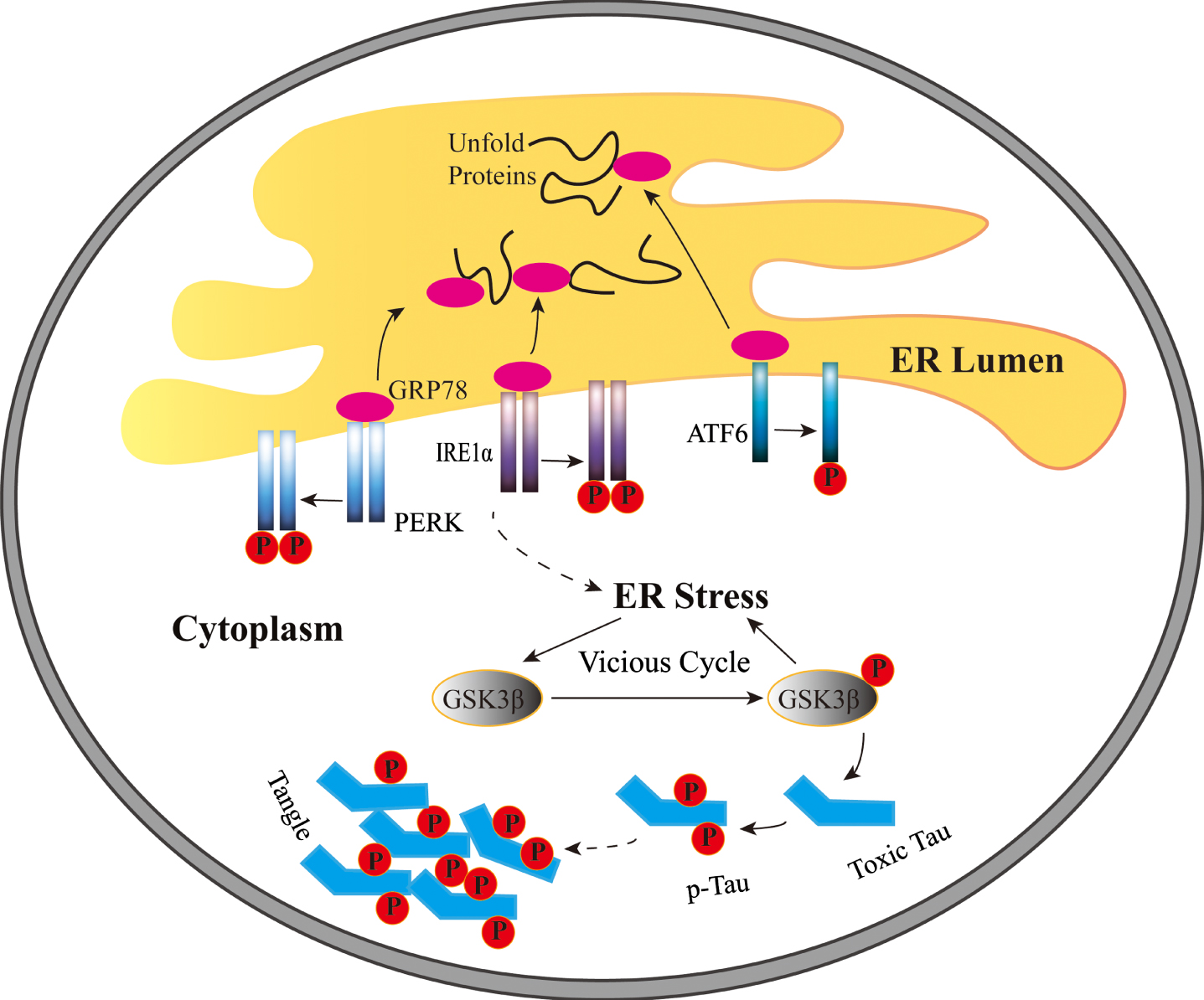
A schematic diagram indicating the current working hypothesis relative to functions of endoplasmic reticulum stress and activity of GSK-3β on SH-SY5Y cells in the non-RA group and the RA group. TM-induced ER stress stimulates intracellular GRP78 to separate from three susceptor and further induce unfolded protein response. In addition, ER stress extenuates Tyr216 phosphorylation of GSK-3β in the non-RA group and accelerates it in the group with RA treatment. Furthermore, tau phosphorylation could be regulated by pGSK-3β-Tyr216. pGSK-3β-Tyr216 could adjust ER stress and phosphorylation of tau when GSK-3β activity is inhibited. It indicates that the relationship between ER stress and GSK-3β exists in a vicious cycle in cells.
Endoplasmic reticulum stress (ERS) plays an essential role in neurodegenerative diseases including amyotrophic lateral sclerosis [5], Huntington’s disease [6], AD [7], and Parkinson’s disease [8], which is characterized by the aggregation and accumulation of misfolded proteins. In addition, ER, as a crucial organelle related to proteostasis, promotes cells toward apoptosis and autophagy [9]. ER homeostasis related to lipid synthesis and storage and release of Ca2 + is preserved through the various signaling pathways, which monitor cell survival, proliferation, and growth [9 –11]. In addition, the imbalance between proteins newly synthesized and folding capacity of ER causes ERS [12]. In mammals, unfolded protein response (UPR) is stimulated by three major ERS sensors, including inositol-requiring transmembrane kinase 1α (IRE1α), protein kinase-like ER kinase (PERK), and activating transcription factor 6 (ATF6) (as shown in Fig. 1) [13]. Glucose-regulated protein 78 (GRP78) is a chaperone heat shock protein (HSP) and the master of the UPR in the ER of normal cells [14].
Tau, as a group of microtubule-associated proteins in neurons which accumulated in neurofibrillary tangles in AD brain, displays an important role in modulating microtubule dynamics, neurite outgrowth, and axonal transport, and all these functions of tau are modulated by site-specific phosphorylation [15]. In AD, human tau is phosphorylated by glycogen synthase kinase 3β (GSK-3β) [16]. Serine 9 phosphorylation of GSK-3β (pGSK-3β-Ser9) triggers GSK-3β inhibition, by contrast, tyrosine 216 phosphorylation of GSK-3β (pGSK-3β-Tyr216) stimulates GSK-3β activation [17]. GSK-3β may play a vital role in neurofibrillary tangles and senile plaques formation, and AD-involved interaction of GSK-3β and tau may be potential therapeutic targets to AD [18].
Zeaxanthin (Zea) is one of carotenoids originated from dark green leafy vegetables and orange and yellow fruits and forms the macular pigment in the human eyes (see Fig. 2A for its chemical structure) [19]. It was hypothesized that lutein and Zea protect against cognition diseases and visual disorders [19]. High serum levels of lutein and Zea were found to be related to a lower risk of AD mortality in adults [20]. In recent years, clinical science underlying carotenoid-based nutritional interventions, including interventions of Zea, meso-Zea, and lutein, was more relevant to ocular disease [21, 22], age-related macular degeneration [23, 24], and cognitive function [25]. Therefore, Zea is a promising nutrient supplementation to future clinical research, and studying functions of Zea in AD is beneficial to find clinical drug of AD.
However, interaction between ERS and GSK-3β in AD was rarely mentioned. After tunicamycin (TM) treatment, GSK-3β was activated; whereas the hippocampus was injected with SB216763 to inhibit GSK-3β activity reduced TM-induced UPR and spatial memory impairment [26]. Small-interfering reduced the expression of GSK-3β by 80%, which can resist cell apoptosis induced by ER stress, and suggested that ER stress-induced apoptosis was partly mediated through the GSK-3β pathway [27]. Our previous study has proved that Zea ameliorated cell damage induced by TM by decreasing caspase-3 activity, protecting the integrity of the mitochondrial membrane and reducing the apoptosis rate [28]. In this study, we aimed to clarify the mechanism among ERS, GSK-3β, and tau phosphorylation, find potential protection of Zea to this mechanism, and further to find a possible treatment approach to AD.
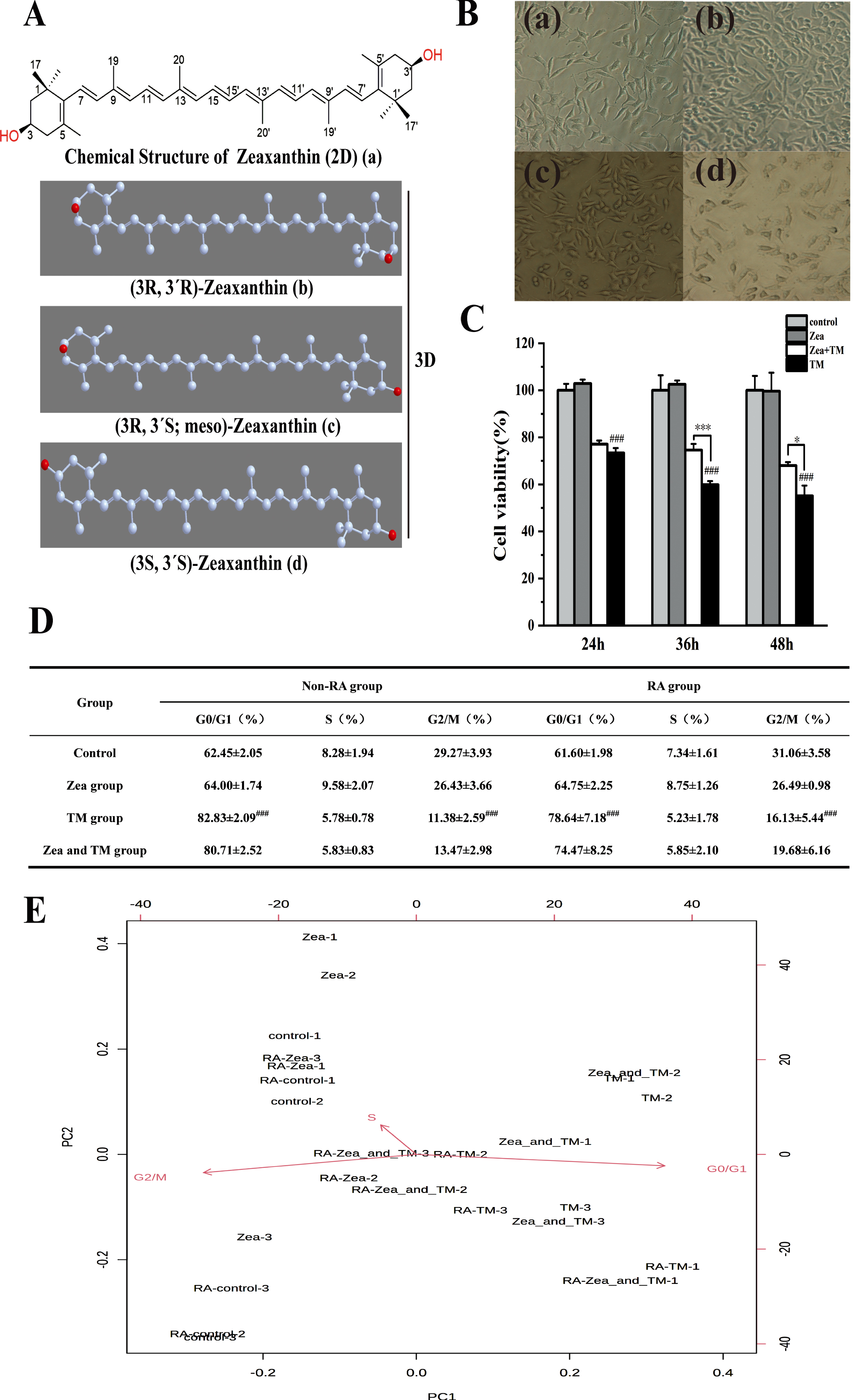
Effect of Zeaxanthin on TM-induced cell injury relative to endoplasmic reticulum stress. A) 2D and 3D chemical structure of Zeaxanthin. (a) Chemical planar structural formulas of Zeaxanthin (2D). (b–d) Stereochemical structural formula of Zeaxanthin (3D). B) Different time of RA treatment to SH-SY5Y cells. (a) control group, (b) RA treatment for 1d, (c) RA treatment for 3d, (d) RA treatment for 6d. C) Cells viability of SH-SY5Y cells after treated with TM (5μg/mL) or Zea (5μmol/L) or TM and Zea cotreatment in different time (24 h, 36 h, 48 h), while Zeaxanthin is used to pretreatment for 6 h (N = 3). Statistical differences (mean±SD) in panel A. # # # p < 0.005 versus control; * p < 0.05, *** p < 0.005 versus TM group. D) Table of cell cycle detection in non-RA and RA group. Values represent mean±SEM; n = 3 in each group. # # # p < 0.005 represents statistically significant differences. E) Biplot figure of programmable counter array to cell cycle in group of non-RA treatment and RA treatment, whereas SH-SY5Y cells of the RA group were treated with 10μmol/L Retinoic acid. Zea group used 5μmol/L Zeaxanthin. TM group used 5μg/mL Tunicamycin. Zea and TM group used Zeaxanthin to pretreat SH-SY5Y cells for 6 h, further used TM and Zea to cotreat cells for 36 h.
MATERIALS AND METHODS
Materials and reagents
The human SH-SY5Y cells used in this study was provided by the Chinese Academy of Medical Sciences in Beijing, China. RPMI 1640 medium (Invitrogen, Gibco, CA, USA) was crushed into powder at –4°C for 2 h and used for culture of cells. Zea was acquired from Shanghai YuanYe Biotechnology in Shanghai, China. TM was purchased from Huayueyang Biotechnology in Beijing, China. 4-Phenylbutyric acid (STBF7437V) was bought from Aldrich. 9-cis-Retinoic acid (R4643) was purchased from Sigma. Cell cycle detection kit (FXP021) was pursed from Beijing 4A Biotech Co., Ltd. Anti-GRP78 was obtained from Cell Signaling Technology in USA. Anti-GSK3 (phospho Y216 + Y279) (Ab68476) was bought from Abcam. Anti-GSK-3β (bsm-33293M), anti-phospho-Tau (Thr231, bs-2368R), anti-phospho-Tau (Ser396, bs-3446R), and anti-total tau (bsm-33332M) were pursed from Beijing Biosynthesis Biotechnology, Co., Ltd. Anti-GAPDH (2118s) was obtained from Cell Signaling Technology. SB216763 (a GSK-3β inhibitor) was purchased from MedChemExpress in the USA.
Cell cultures
SH-SY5Y cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine, 100μg/mL streptomycin, and 100 U/ml penicillin, and then incubated in a CO2 incubator under the conditions of 37°C and 5% CO2. Cells with density of 105 cells /mL were cultured containing 3 mL medium each of growth media. After overnight, SH-SY5Y cells were differentiated with 9-cis-Retinoic acid (RA) in the media, observing for neurite out-growth in RA group. Cells were examined for neurite outgrowth at 1 day, 3 days, and 6 days, with media being changed after every 48 h [29].
Drugs exposure
SH-SY5Y cells in non-RA group and cells after RA treatment for 6 days were added into culture dishes and then reached at least 80% confluence before further treatment. Cells were divided into four groups: control group, TM group (treated with TM at 5μg/mL), and Zea group (treated with Zea at 5μM) and TM + Zea group (cotreatment with 5μg/mL TM and 5μM Zea). Cells were incubated with 4-PBA (an ERS inhibitor) at 5 mM for inhibiting ERS or SB216763 (a GSK-3β inhibitor) at 20μM. Then we established additional inhibitor groups: TM + 4-PBA group, TM + Zea + 4-PBA group, TM+SB216763 group, and TM + Zea + SB216763 group.
MTT assay
SH-SY5Y cells were added into 96-well plate about 1×105 cells of each well and incubated for almost 24 h. Next, cells were treated with drugs for different time and then detect cell viability. Each well added 20μL of a 5 mg/mL MTT solution to 200μL medium and the plate was further incubated for 4 h at 37°C, and then, the plate was centrifuged at 1000 g in 4°C for 10 min. 150μL of DMSO was further added to each well. For dissolving the dye, 96-well plate was placed on a shaker for 10 min. the absorbance was determined spectrophotometrically at 570 nm by using a reference wavelength of 630 nm.
Cell cycle analysis
SH-SY5Y cells was cultured in RPMI 1640 medium in an incubator at 37°C with 5% CO2 at 3×104 cells/mL and placed in a constant temperature incubator at 37°C overnight. After adding drugs according to the experiment, the culture was continued for 36 h. After the cells were digested with trypsin, the digestion was further terminated in \nobreak complete medium, and the cells were gently resuspended in the culture medium. After following the instructions of the cell cycle test kit, we detected the cell cycle in a flow cytometer, collected the data in CELL QUEST PRO software, and analyzed the cell cycle in Modifit LT software.
Sample preparation and western blot analysis
SH-SY5Y cells and cells treated by RA were collected and lysed by using a 100μL RIPA lysis buffer with protease inhibitors. The lysates were centrifuged at 12,000 g in 4°C for 10 min, and the supernatant was collected. Then, the remaining protein sample was mixed with 5×SDS PAGE loading buffer in a ratio of 4:1, placed into the boiling water bath for 5–10 min, and cooled to room temperature. Protein concentrations were measured using BCA assay kits. The 20μg of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE, 8–15%) gel and then transferred to polyvinylidene fluoride membranes (Millipore, USA). Non-specific binding proteins were blocked with 5% (wt/vol) skim milk for 1 h at room temperature. The membranes were incubated with primary antibodies against anti-GRP78, anti-pGSK-3β-Tyr216, anti-GSK-3β, anti-p-Tau (Thr-231), anti-p-Tau (Ser-396), anti-t-Tau and anti-GAPDH overnight at 4°C, and then incubated with the corresponding horseradish peroxidase-linked secondary antibodies for 2 h at room temperature. The optical density was detected with a Quant RT ECL Gel Imaging System. Quantification of the optical density was performed by using Image J. Tau-related protein biomarkers were analyzed in the present study and normalized against GAPDH.
Statistical analysis
Values for all measurements are presented as means±standard deviation. One-way ANOVA was performed with Newman–Keuls Multiple Comparison posthoc test. p < 0.05 was considered statistically significant.
RESULTS
Visual appearance of SH-SY5Y cells treated by RA
The SH-SY5Y cell line was often used as a surrogate for neurons in vitro studies. Quantitative analysis of over 9400 proteins across a 7-day treatment with retinoic acid by using proteomics technologies based on state-of-the-art mass spectrometry could represent a rich resource for investigating temporal protein abundance changes in SH-SY5Y cells differentiated with retinoic acid [30]. Sequential exposure of SH-SY5Y cells to retinoic acid could yield homogeneous populations of cells with neuronal morphology [31]. Treatment with RA may augment survival signaling and attenuate basal apoptotic pathways in SH-SY5Y cells, which increased cell viability. SH-SY5Y cells became elongated from fusiform or teardrop shape after RA treatment for 0–6 days, and the length of cell synapses gradually increased (Fig. 2B). Photomicrographs were obtained with a 20×objective lens on a Nikon ECLIPSE camera (TE2000-M) adapted to the microscope.
Effect of Zea on cell injury induced by TM in non-RA group and RA group
As shown in Fig. 2C, SH-SY5Y cells were treated with TM and Ze for 24 h, 36 h, and 48 h after 6 h of Zea pretreatment. Compared with the blank group, the cell survival rate decreased significantly at different times when treated with TM alone (p < 0.001). Compared with TM treatment alone, effect of co-treatment at different times reversed the TM-caused damage of SH-SY5Y cells. It existed significant changes between the TM + Zea group and TM group. Compared with the TM injury group, the survival rate of SH-SY5Y cells increased significantly at 36 h (p < 0.01). Therefore, the co-treatment time of TM and Zea was selected as 36 h for subsequentexperiments.
Compared with control group, the cell number in the G0/G1 phase of the TM injury group increased significantly in non-RA group and RA group (p < 0.005), which were respectively 82.83% and 78.64% (Fig. 2D), and the G2/M phase cells in the TM treatment group were significantly reduced in the non-RA group (p < 0.005). Compared with the TM model group, the number of cells in the G0/G1 phase decreased in TM + Zea group. It indicated that ERS could cause the cell to block the G1 phase in both the undifferentiated and differentiated group, and Zea could reverse this block. As presented in Fig. 2E, biplot figure of programmable counter array to cell cycle in non-RA and RA group indicated that TM group and TM + Zea group were more relevant to G0/G1 period. This result was consistent with the result in Fig. 2D.
Function of Zea on the expression of ERS-related protein GRP78
GRP78 as a central regulator of ERS plays a key roles in cell survival and apoptosis through modulated the activation of transmembrane ERS sensors: PERK, IRE1, and ATF6 [32]. Figure 2A-C showed that the levels of GRP78 expression in non-RA and RA group increased significantly after 5μg/mL TM treatment (p < 0.005) in comparison with the control group. Compared with the TM group, after treatment with 5μM Zea, the expression level of GRP78 decreased significantly in the non-RA treatment group (p < 0.005) (Fig. 3A, B), and the expression level of GRP78 in the RA group decreased significantly (p < 0.05) (Fig. 3A, C). Previous study indicated that TM could induce endoplasmic reticulum stress and activation of ER stress-associated proteins GRP78 [33]. After adding the ERS inhibitor, compared with the TM group, the TM + 4-PBA group showed a significant decrease in protein levels in both groups (p < 0.05), and further adding Zea, the GRP78 levels of the two models occurred reversed, and there was a very significant difference in the undifferentiated group (p < 0.01) (Fig. 3A-C). The results show that Zea can alleviate the increase of GRP78 expression in the ERS cell model and Zea had a certain effect on alleviating ERS.
Effect of Zea on regulation to GSK-3β activity and phosphorylation of tau expression in non-RA group with inhibition of ERS or activity of GSK-3β
GSK-3β appeared to be activated by its phosphorylation at tyrosine 216 [34]. As shown in Fig. 3D-F, compared with control group, TM treatment in non-RA group reduced the activity of GSK-3β, while Zea significantly alleviated the decrease in p-GSK-3β/GAPDH caused by TM (p < 0.01), relieving significantly the decrease of the level of p-GSK-3β/t-GSK-3β caused by TM (p < 0.05) (Fig. 3F). Compared with TM group, the relative expression of p-GSK-3β/t-GSK-3β decreased in TM + 4-PBA group, and Zea alleviated this decrease. It suggested that ERS can reduce activity of GSK-3β in the TM-induced ERS model. After ERS is inhibited, the effect of TM on GSK-3β activity decreased, and Zea can alleviate it.
Two major tau kinases (GSK-3β and cyclin-dependent kinase 5) were involved in Pb-induced tau hyperphosphorylation [35]. As shown in Fig. 3H-J, the p-Tau (Ser396)/GAPDH in the TM group was significantly different to control group (p < 0.005). The expression of p-Tau (Ser396)/GAPDH was unregulated in the co-treatment of Zea and TM compared with TM group. Compared with TM group, there was a significant difference in p-Tau (Thr231)/GAPDH in the TM and Zea co-treatment group (p < 0.05). p-Tau (Ser396)/t-Tau and p-Tau (Thr231)/t-Tau showed extremely significant differences in TM group compared with control group (p < 0.005). p-Tau (Ser396)/t-Tau and p-Tau (Thr231)/t-Tau were upregulated compared with the TM group. Compared with TM group, the relative expression levels of p-Tau (Ser396) and p-Tau (Thr231) in TM + 4-PBA group decreased. Zea additional treatment can upregulate the expression of tau phosphorylation in comparison with TM + 4-PBA group. Studies have shown that ERS can inhibit Tau phosphorylation in non-RA group, and the results were consistent with the relative expression trend of pGSK-3β-Tyr 216 protein.
Inhibiting the activity of GSK-3β can decrease the expression of p-GSK-3β and p-Tau in non-RA group, and Zea can reduce the decrease of expression
As shown in Fig. 4 A-C, compared with the TM group, the expression of GSK-3β phosphorylation decreased after additional inhibitor treatment, and TM+Zea+SB216763 could increase the expression of p-GSK-3β. The experiment showed that the GSK-3β inhibitor SB216763 can effectively inhibit the Tyr216 phosphorylation of GSK-3β, thereby inhibiting the activity of GSK-3β, which was consistent to two previous study finding that SB216763 could inhibit the activity of GSK-3β [36, 37].
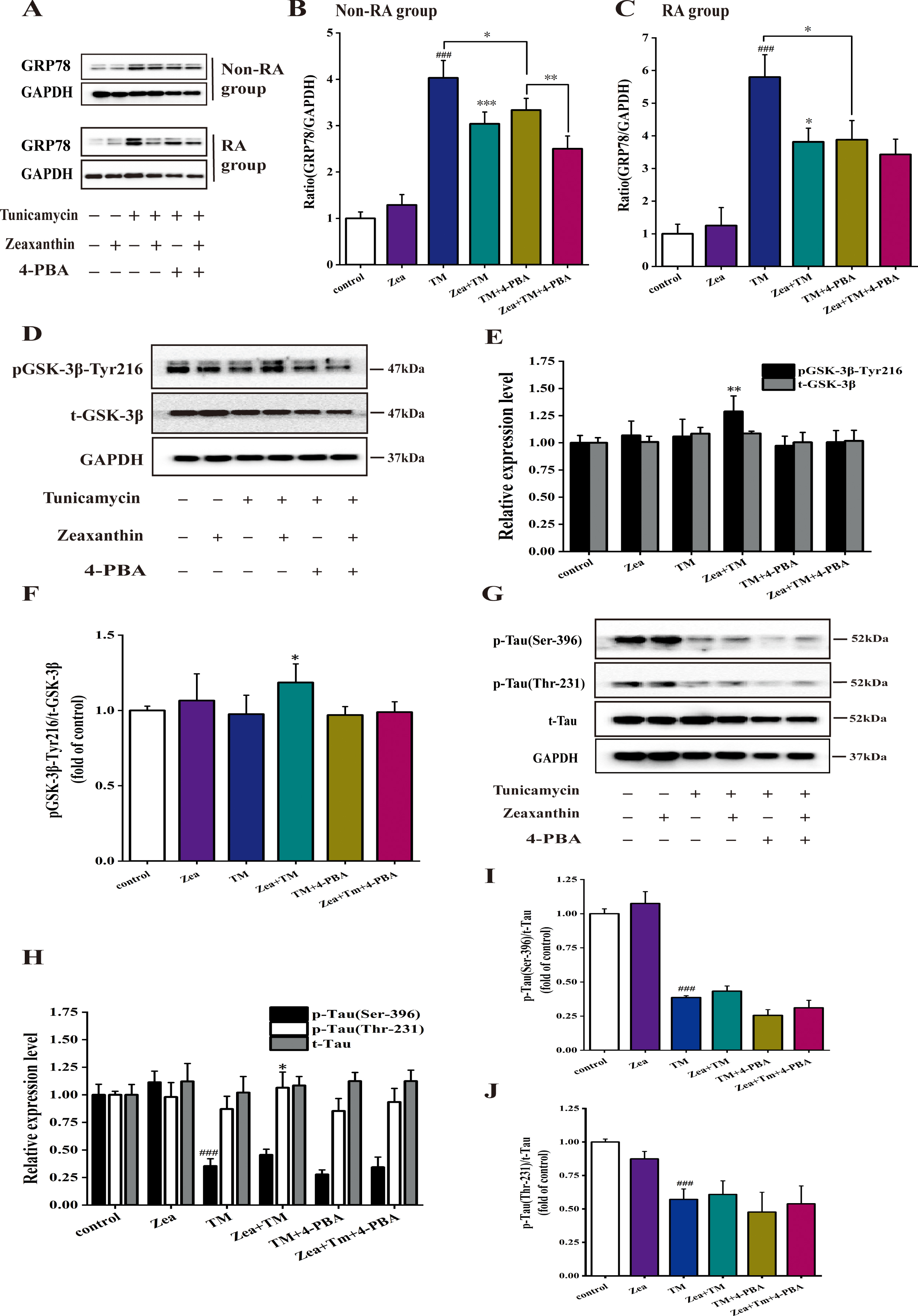
Zeaxanthin regulated increase of GRP78 expression and the decrease of pGSK-3β-Tyr216, p-Tau (Ser-396), and p-Tau (Thr-231) protein induced by TM. A) GRP78 and GAPDH expression was detected by western blot. This experiment was divided into six groups, containing control group, Zea group, TM group, Zea and TM group, TM and 4-PBA (an ERS inhibitor, 5 mM) group, Zea, TM, and 4-PBA group. B) Histograms of the relative amount of GRP78 based on the western blotting in non-RA group. C) Histograms of the relative amount of GRP78 based on the western blotting in RA group. # # # p < 0.005 versus control; * p < 0.05, ** p < 0.01, *** p < 0.005 versus TM group. Zeaxanthin alleviates the decrease expression of pGSK-3β-Tyr216 and p-Tau (Ser-396 and Thr-231) protein caused by ERS in non-RA group of SH-SY5Y cells. D-J) SH-SY5Y cells were incubated with TM (5μg/mL), Zea (5μM), and 4-PBA (5 mM), then the cell extracts were prepared for analyses of Tyr216 phosphorylation of GSK-3β (D–F), total GSK-3β (D, E), total tau (H, I), phosphorylated tau at Thr231, Ser396 (H-J). # # # p < 0.005 versus control group; * p < 0.05, ** p < 0.01 versus TM group at the same time point (mean±SD).

Zeaxanthin regulated the decrease of pGSK-3β-Tyr216 and p-Tau (Ser-396) protein expression and the increase of expression of p-Tau (Thr-231) when GSK-3β activity is inhibited in non-RA group of SH-SY5Y cells. A–G) SH-SY5Y cells were incubated with TM (5μg/mL), Zea (5μM), and SB216763 (20μM), then the cell extracts were prepared for analyses of Tyr216 phosphorylation of GSK-3β (A–C), total GSK-3β (A, B), total tau (E, G), phosphorylated tau at Thr231,Ser396 (D–G). # # # p < 0.005 versus control group; * p < 0.05, ** p < 0.01 versus TM group at the same time point (mean±SD).
Western blot results showed that compared with control group, TM treatment can significantly reduce the level of Ser396 and Thr231 phosphorylation of Tau (p < 0.005) as shown in Fig. 4D-G. Among the six groups related to GSK-3β inhibition treatment, compared with TM group, TM + Zea group reduced Ser396 site phosphorylation of tau and increase the expression of p-Tau (Thr231). After adding GSK-3β inhibitor, tau phosphorylation levels were reduced. Compared with the TM+SB216763 group, the TM + Zea + SB216763 group reduced the level of tau phosphorylation.
Effect of Zea on regulation to GSK-3β activity and phosphorylation of tau expression in RA group with inhibition of ERS or activity of GSK-3β
As shown in Fig. 5A-C, TM can significantly increase the expression of p-GSK-3β (p < 0.005) in RA group. Compared with the TM group, p-GSK-3β expression of TM+Zea group decreased significantly (p < 0.05) and GSK-3β activity of TM + SB216763 group was significantly reduced (p < 0.005). Zea can reduce the activity of GSK-3β to group treated with GSK-3β inhibitor. It indicated that ERS can promote an increase of GSK-3β activity in RA group.
In Fig. 5D-G, compared with the control group, the expression of p-Tau (Ser396) and p-Tau (Thr231) in TM group increased significantly (p < 0.005). Zea and TM group showed a downregulation level of tau phosphorylation compared with TM group. Tau phosphorylation caused by additional treatment of 4-PBA could be downregulated compared with TM group. This trend of the results was consistent with result of the GSK-3β in RA group. Interestingly, Zea has an inhibitory effect on the expression of tau phosphorylated protein in TM + SB216763 group, and this inhibitory effect has a significant difference in the expression of p-Tau (Ser396) (p < 0.05) and very significant difference in the expression of p-Tau (Thr231) (p < 0.005). In summary, ERS can promote tau phosphorylation in RA group, inhibiting ERS can weaken the pro-phosphorylation effect, and Zea can alleviate the effect in TM+ERS inhibitor group.
Inhibiting the activity of GSK-3β can decrease the expression of p-GSK-3β and in RA group, and Zea can reduce this decrease of expression
Resveratrol can significantly inhibit ERS induced by 2-DG and SB216763 blocked the function of resveratrol, indicating that resveratrol can prevent protects cardiomyocytes from ERS by GSK-3β pathway [38]. TM group significantly increased the phosphorylation of GSK-3β compared with control group (p < 0.005) in Fig. 6A-C. Compared with the TM group, the Zea + TM group can significantly reduce the activity of GSK-3β (p < 0.05), and the TM + SB216763 group has a very significant decrease in GSK-3β activity (p < 0.005). The additional treatment of Zea can further reduce the phosphorylation of GSK-3β of TM + SB216763 treatment.
As shown in Fig. 6D-G, TM can significantly increase the phosphorylation of tau in Ser396 site (p < 0.005), and significantly increase the phosphorylation of Tau in Thr231 site (p < 0.01) compared to control group. Zea treatment can alleviate the phosphorylation of tau caused by TM, and the phosphorylation of Tau in Ser396 site was significantly different (p < 0.01). Additional SB216763 treatment can reduce the expression level of p-Tau compared to TM group, and Zea+TM+SB216763 group can be found to alleviate the decrease of tau phosphorylation in TM+SB216763 group.
Inhibiting the activity of GSK-3β can decrease the expression of GRP78 in non-RA and RA group, and Zea can reduce the decrease of expression
Aging-induced GRP78 expression was associated with GSK-3β-Tyr216-mediated c-Jun N-terminal kinase phosphorylation, which was downregulated upon Capsosiphon fulvescens glycoproteins administration [39]. Figure 6H-J shows that whether in non-RA group or in RA group, GSK-3β inhibitor can downregulate the increase in GRP78 expression in cells caused by TM, its downregulation effect is significantly different in non-RA group (p < 0.005), the downregulation effect was significantly different in RA group (p < 0.05). Compared with the TM+SB216763 group, the Zea+TM+SB216763 group can reduce the expression of GRP78.
DISCUSSION
Many neurodegenerative diseases including AD, Parkinson’s disease, amyotrophic lateral sclerosis, and prions diseases were found to be involved in ER and UPR dysfunctions by the accumulation and aggregation of misfolded proteins [40]. Due to survival or death, the pharmacological manipulation of ER-UPR may become the focus of potential therapies in tauopathies [41]. The immunoreactivity of phosphorylated tau co-localized with that of p-PERK in the hippocampus of aged P301L mutant tau transgenic mice and researchers hypothesized that ERS could induce phosphorylation of tau and vice versa in inducing AD-like pathogenesis [42]. Our study did not verify the changes between ERS and tau phosphorylation in neurons and brains of animals and other mammals, nor did we study further neurofibrillary tangles relevant to AD.
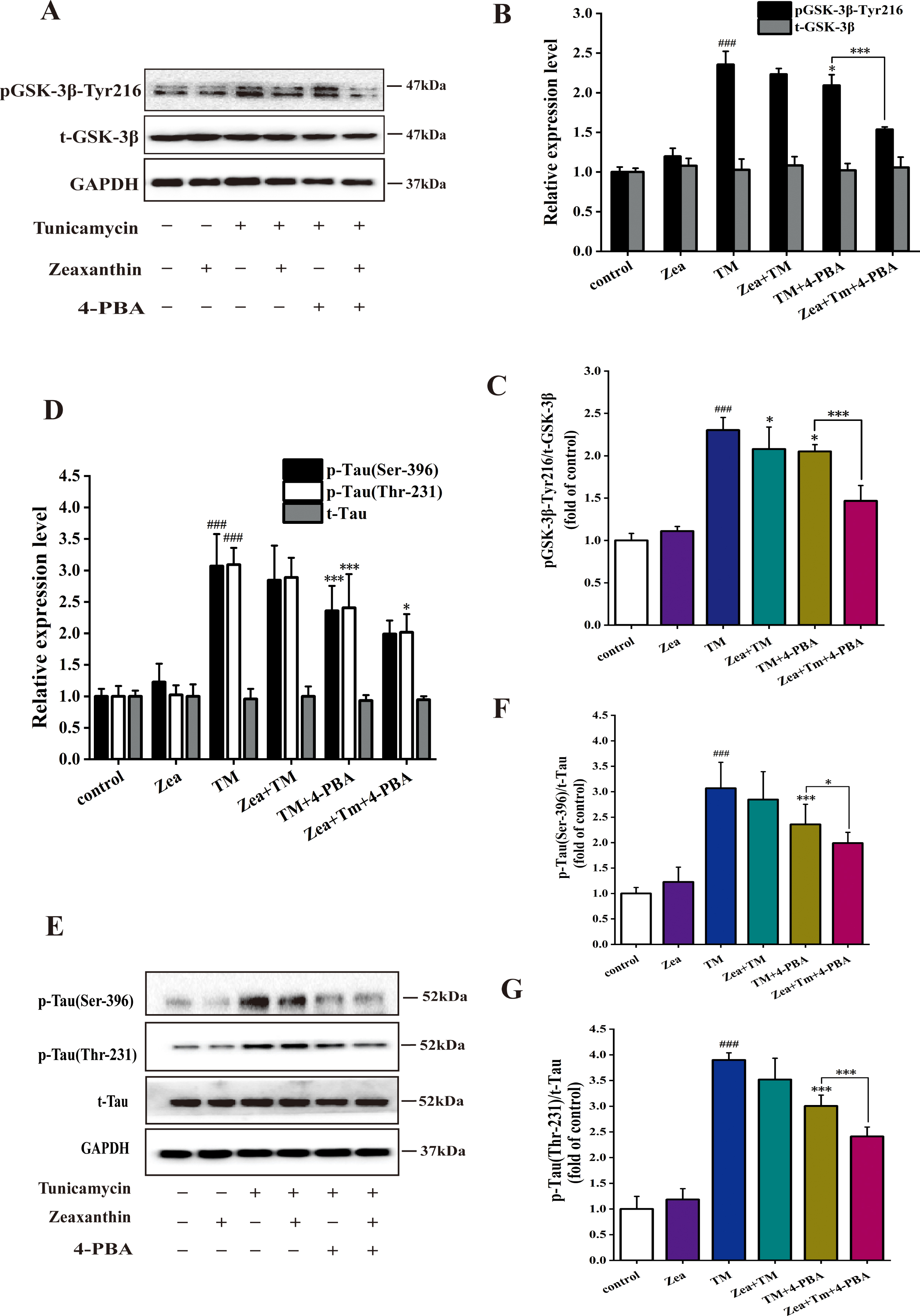
Zeaxanthin downregulated the increase of expression of pGSK-3β-Tyr216 and p-Tau (Ser-396 and Thr-231) protein caused by ERS in RA group of SH-SY5Y cells. A-G) SH-SY5Y cells were incubated with TM (5μg/mL), Zea (5μM), and 4-PBA (5 mM), then the cell extracts were prepared for analyses of Tyr216 phosphorylation of GSK-3β (A–C), total GSK-3β (A, B), total tau (D, E), phosphorylated tau at Thr231,Ser396 (D–G). # # # p < 0.005 versus control group; * p < 0.05, ** p < 0.01 versus TM group at the same time point (mean±SD).
Nowadays, more and more research correlated to AD were associated with beneficial ingredients including omega-3 fatty acids (ω3-FAs) [43], carotenoids [43, 44], and vitamin E [45]. The effect of ω3-FAs with testing of spinal fluid and memory tests showed that an interaction between ω3-FAs and B vitamin status in relation to cognitive decline and brain atrophy [46]. Study shows that carotenoids may be powerful anti-amyloidogenic molecules in preventing AD, especially since most of them are considered nutraceutical compounds and can cross the blood-brain barrier [47]. The properties of vitamin E to anti-inflammatory and hypocholesterolemia causes its importance for brain health, and the levels of it in AD patients when lower than in non-demented controls indicating vitamin E might be a good candidate to against AD [48]. Significant beneficial effects of vitamins C and E in women with APOE E4 and men without APOE E4 may reduce the risk of cognitive decline and may minimize the risk for AD [49].
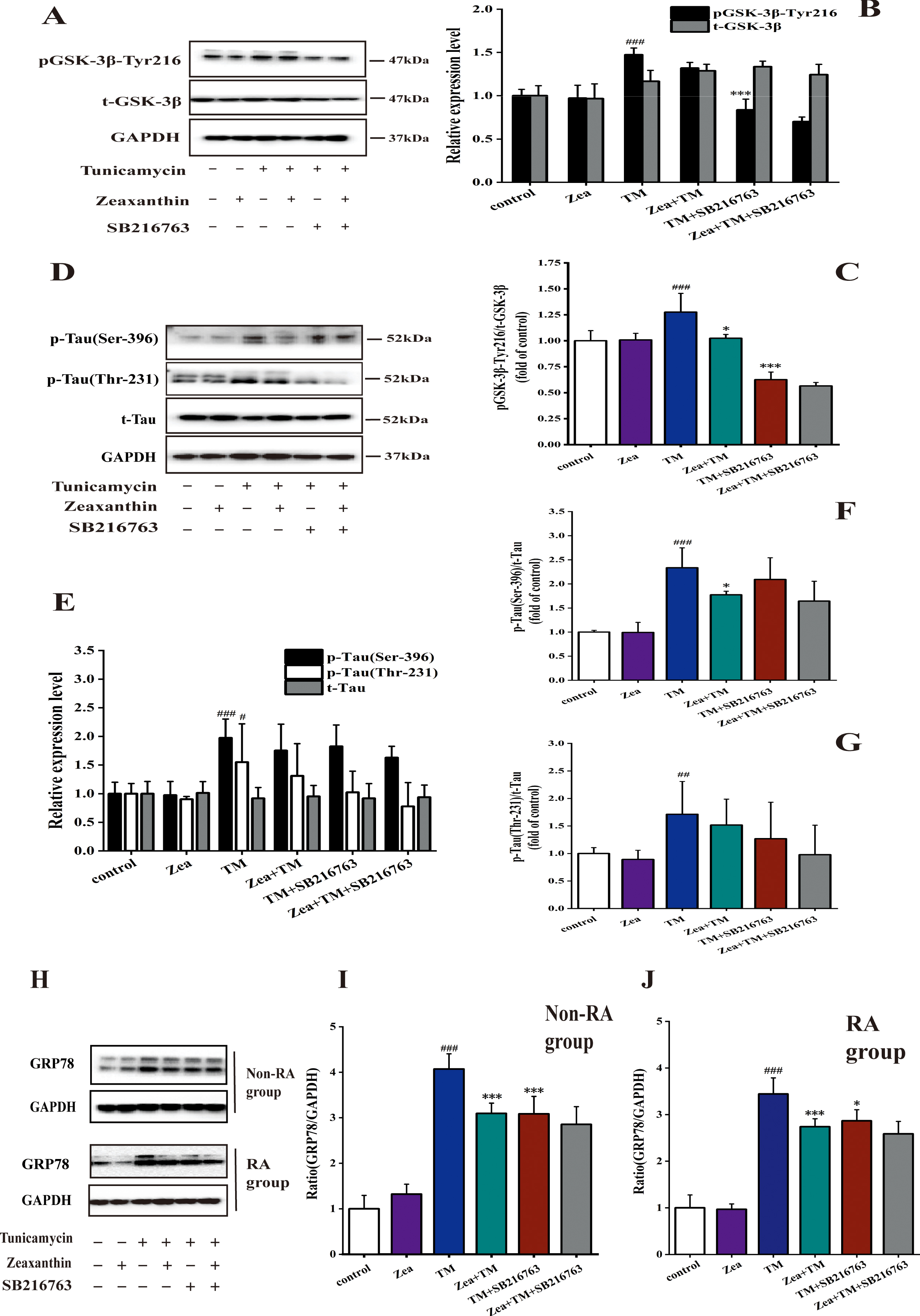
Zeaxanthin downregulated the decrease of expression of pGSK-3β-Tyr216, p-Tau (Ser-396 and Thr-231) protein, and GRP78 caused by GSK-3β activity inhibition in RA group of SH-SY5Y cells. A–G) SH-SY5Y cells were incubated with TM (5μg/mL), Zea (5μM), and SB216763 (a GSK-3β activity inhibitor, 20μM), then the cell extracts were prepared for analyses of Tyr216 phosphorylation of GSK-3β (A–C), total GSK-3β (A, B), total tau (D, E), phosphorylated tau at Thr231,Ser396 (D–G). # # p < 0.01, # # # p < 0.005 versus control group; * p < 0.05, *** p < 0.005 versus TM group at the same time point (mean±SD). H) GRP78 and GAPDH expression was detected by western blot. This experiment was divided into six groups, containing control group, Zea group, TM group, Zea and TM group, TM and SB216763 (20μM) group, and Zea, TM, and SB216763 group. I) Histograms of the relative amount of GRP78 based on the western blotting in non-RA group. J) Histograms of the relative amount of GRP78 based on the western blotting in RA group. # # # p < 0.005 versus control; * p < 0.05, *** p < 0.005 versus TM group.
In this work, the effects of Zea and TM on SH-SY5Y cells were investigated. We found that Zea extenuated TM-induced cell injury, including upregulation of cell viability and cell numbers of G0/G1 phase. TM, a ER stress activator, induced apoptosis [50], made G1 checkpoint arrest [51], and decreased cell viability [52]. Zea shows its protective role on human limbal and conjunctival epithelial cells against UV-induced cell death and oxidative stress [53]. Supplementation with the macular carotenoids (meso-Zea, Zea, lutein) benefited patients with AD [54]. The more favorable feature is represented by the protective role of Zea in reducing ERS-caused expression of GRP78 in non-RA and RA groups. The result in non-RA group about GRP78 was mentioned in our past result [28].
Increasing evidence suggests that hyperphosphorylated tau protein plays a pivotal role in the occurrence and development of AD via activation of GSK-3β signaling pathway [55 –57]. Our results indicated that Zea attenuated the decrease of GSK-3β activation and tau phosphorylation induced by ERS, and GSK-3β activity inhibition can lower ERS-caused change of GSK-3β activation, tau phosphorylation, and expression of GRP78 in non-RA group. These suggest that Zea could influence activation of GSK-3β in non-RA group to modulate ERS and tau-involved change.
Moreover, the upregulation of GSK-3β mediated by the loss of miR-219-5p may be a possible mechanism that leads to tau phosphorylation and progression of AD [58]. In the RA group of our study, Zea alleviated the increase of GSK-3β activation and tau phosphorylation induced by ERS, and GSK-3β activity inhibition also can reduce ERS-caused change of GSK-3β activation, tau phosphorylation, and GRP78 expression, which indicated that ERS could lead to tau phosphorylation by regulating theGSK-3β pathway. Meanwhile, our results were similar to the study that increased GRP78 was found in AD model mice, and it induced tau hyperphosphorylation via activating GSK-3β in HEK293 cells stably expressing the longest human tau (HEK293/tau) or N2a cells [59].
Conclusion
In summary, there exists a vicious circle among ERS, tau phosphorylation, and GSK-3β activation in non-RA and RA groups (schematic diagram as shown in Fig. 1). Therefore, ERS contributed by TM may modulate tau phosphorylation which involves GSK-3β activation and inhibition, and then may cause AD-related neurofibrillary tangles and AD pathology.
Footnotes
ACKNOWLEDGMENTS
The authors are grateful to Beijing Key Laboratory of Bioactive Substances and Functional Food and the Institute of Functional Factors and Brain Science.
This work was supported by the National Natural Science Foundation of China (31471587), and the Academic Research Projects of Beijing Union University (ZK40201902, JZ10202001, XP202008).