
Abstract
Background:
Acetylcholinesterase (AChE) regulates the transmission of neural messages by hydrolyzing acetylcholine in synaptic spaces.
Objective:
The effects of many AChE inhibitors have been evaluated in the treatment of Alzheimer’s disease, but the present study examined a synthetic complex containing cobalt (SC) for the first time in the field of enzyme activity to evaluate enzyme inhibitory function.
Methods:
Ellman’s test was applied. AChE function was assessed in the presence of SC through docking and molecular dynamics analyses. The second structure of AChE was studied through circular dichroism (CD) spectroscopy.
Results:
Several enzymatic methods were utilized for the kinetics of AChE, which indicated the non-Michaelis and positive homotropic behavior of AChE in the absence of inhibitors (Hill coefficient = 1.33). However, the existence of inhibitors did not eliminate this homotropic state, and even AChE had a more sigmoidal shape than the galantamine at the presence of SC. Based on the CD spectroscopy results, AChE structure changed in the existence of inhibitors and substrates. Bioinformatics analysis revealed SC bonding to the channel of active site AChE. The number of hydrogen bonds was such that the flexibility of the enzyme protein structure due to inhibitor binding reduced AChE function.
Conclusion:
The results reflected that AChE exhibited a non-Michaelis and positive homotropic behavior, leading to a more inhibitory effect on the SC than the galantamine. The positive homotropic behavior of AChE was intensified due to the alteration in AChE protein structure by binding SC to hydrophobic region in the active site pathway and impressing Trp84.
INTRODUCTION
Acetylcholinesterase (AChE, EC 3.1.1.7), a serine protease family, is considered as one of the most important enzymes in the nervous system [1]. Upon the completion of neurotransmission, this enzyme results in hydrolyzing acetylcholine as a neurotransmitter in cholinergic synapses [2]. Based on the results of the previous studies, AChE consists of the two binding sites of catalytic and peripheral for acetylcholine as a substrate [3]. AChE is characterized by a deep narrow channel with 20 Å length, which is called gorge or active site channel [3, 4]. The catalytic site of the enzyme is located at the bottom of the gorge, which includes two parts. The first is an ester segment with three amino acids Ser, His, and Glu, which is the main feature of all serine hydrolases. The other is an oxyanion hole composed of amino acids Gly and Ala [5]. The second substrate binding site, at the head of the gorge, is known as the peripheral anionic site, which is responsible for binding ammonium acetylcholine part. Acetylcholine binds to the enzyme with positive nitrogen charge [6]. In addition, the middle part of the enzyme (channel) is covered by the aromatic acid side chains of Trp, Tyr, and Phe, and little amino acids. However, the enzyme channel is hydrophobic throughout. The amino acids are highly conserved in various animal species [4, 7]. During the early 20th century, some researchers have found that the type of the interactions between the catalytic and peripheral sites affects the affinity of the ligand and enzyme [8]. The peripheral site provides a transient binding site for substrate, and consequently is effective in the catalytic efficiency of the enzyme [9]. This interaction between substrate and aromatic amino acids in peripheral site has a significant effect on enzymatic inhibition. One of the amino acids Trp84 or 86 influences the catalytic activity of the enzyme by creating two conformations [5]. In fact, this amino acid can act as a peripheral anionic subunit [10] and accept substrate, or make the substrate access to the active site difficult [11, 12].
AChE inhibitors such as tacrine, rivastigmine, galantamine, and donepezil are mostly used to treat and slow the progression of Alzheimer’s disease (AD). The drugs counteract a decrease in acetylcholine levels in the synaptic space since acetylcholine deficiency leads to the death of cholinergic neurons [13]. Further, galantamine is a selective and rapidly-reversible AChE inhibitor, which interacts with the anionic site and aromatic gorge [3]. It is an alkaloid compound extracted from plant Galanthus woronowii [3]. This drug is administered to treat mild to moderate AD in spite of associating with side effects such as weakness, nausea, vomiting, and weight loss. Therefore, drugs with fewer side effects should be replaced [3, 14]. It is prescribed for the treatment of mild to moderate AD despite its side effects such as weakness, nausea, vomiting, and weight loss. Patients with AD dementia had 12 points higher Mini-Mental State Examination scores at 6 months. They will benefit from functional abilities and will maintain their cognitive abilities within the baseline [15]. Galantamine can improve cognitive function in up to 70% of patients [16].
The synthetic complex (SC, [CoIII(L1) (py)3]ClO4), introduced in the previous research, was used in the present study [17]. This complex contains a pyridine ring and cobalt (III) (Fig. 1).
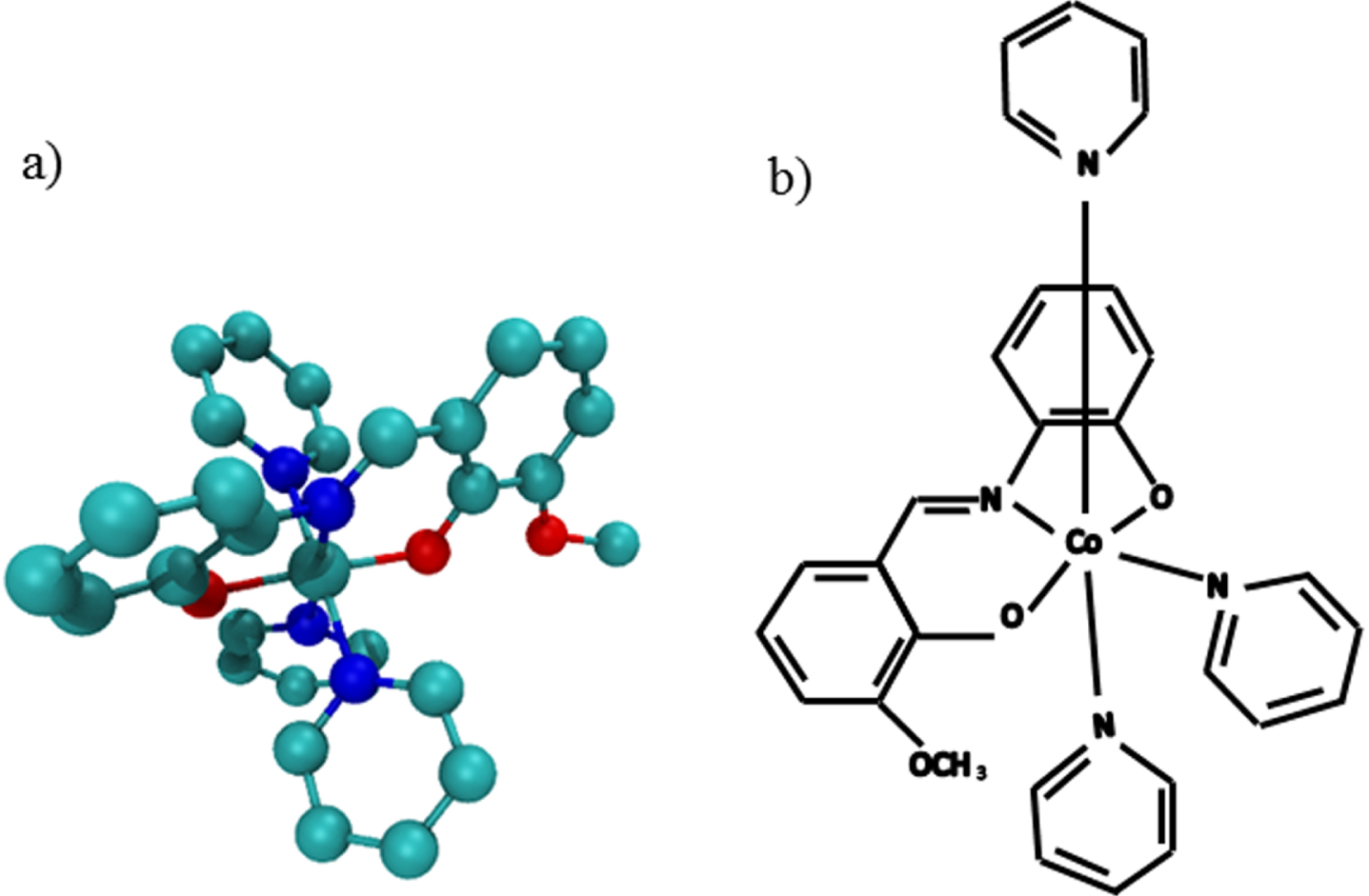
The synthetic complex [CoIII(L1)(py)3]ClO4, a) The two-dimensional image of synthetic complex. b) The 3D image of synthetic complex.
The biological importance of pyrimidine complexes has been emphasized. This SC is considered as a suitable inhibitor of gram-positive bacteria such as Staphylococcus aureus and Bacillus subtilis due to its easy passage through the bacterial membrane compared to the chloramphenicol and kanamycin antibiotics [17]. Given that the complex has been synthesized during the recent years, it needs further research about clinical importance in AD to exploit its properties. Thus, the present study evaluated AChE activity by the SC and compared the results with the galantamine.
METHODS
AChE from Electrophorus electricus (type V-S), acetylthiocholine iodide (ATCI), dithiobis nitrobenzoic acid (DTNB), and galantamine were purchased from Sigma Aldrich. The synthetic complex ([CoIII(L1)(py)3]ClO4) was synthesized in Semnan University.
Synthesis of Cobalt (III) complex
The complex was synthesized in good yields by reaction between the Schiff base ligand (3-methoxy-2-{(Z)[(2-hydroxyphenyl)imino]methyl}phenol (H2L1)) and Co(CH3COO)2.4H2O in methanol following similar procedure as described before [17]. A solution of Co (CH3COO)2.4H2O (0.125 g, 0.5 mmol) in methanol was added to a stirred solution of H2L1 (0.122 g, 0.5 mmol) in methanol. The pink solution turned brown immediately. pyridine (0.31 mL, 4 mmol) was added to the resulting solution, and air was bubbled through the reaction mixture for about 5 h. A solution of NaClO4 (0.061, 0.5 mmol) was then added to the resulting brown solution. The precipitate was washed and dissolved in methanol. Single crystals were obtained by slow evaporation of the methanol. The crystals were filtered off, washed with cold methanol, and dried under vacuum.
FT-IR: υmax cm–1 (KBr): 1,602 (s, C = N), 1,095 (s, ClO4). UV–Vis:λmax (nm) (ɛ, M–1 cm–1) (CH3CN): 525 (542), 408 (8,900), 323 (6,700), 247 (39,000). 1H-NMR (CDCl3, 500 MHz): δ= 3.95 (s, 3Ha), 6.66–8.08 (23H, Aromatic ring proton and Imine proton). Anal. Calcd. for C29H26CoN4O7Cl: C, 54.7; H, 4.1; N, 8.8. Found: C, 54.5; H, 4.3; N, 8.7 %.
Spectroscopic characterization of Cobalt (III) complex
The complex was synthesized and characterized by physico-chemical and spectroscopic methods. In the FT-IR spectra of the complex, strong sharp absorption band at 1,602 cm–1 can be assigned to the C = N(imine) stretching vibration. Compared to the free ligand, these bands were shifted to lower wave numbers upon coordination. The stretching vibration of ClO4– anion is observed at 1,095 cm–1. Charge transfer and intra-ligand transitions occur in the 411–192 nm region and the molar extinction coefficients of these bands are higher than those observed for the free ligand. In 1H-NMR spectra of complex singlets at 3.95 and 3.80 ppm are assigned to the CH3 protons of the Schiff base ligand. The crystal structure of the complex were determined by X-ray crystallography and completely discussed in [17]. The Cobalt (III) center in this complex has a slightly distorted octahedral environment.
Assessment of AChE enzyme activity and inhibition
An enzyme activity assay was performed through employing Ellman’s method [18, 19]. In this method, acetylthiocholine (substrate) is applied as a substrate and the thiol group of the product is measured by using DTNB. The catalytic activity of the enzyme enhances following an increase in the production of yellow 2-nitro-5-thiobenzoate (A412) anion by interacting thiocholine with reagent DTNB. One unit of AChE is defined as the amount of enzyme which converts one micromole of substrate into product per minute. Accordingly, the assay was carried out in a 96-well microliter plate in a total volume of 105μl including 0.1 mM phosphate buffer solution at pH = 8, 0.03 unit/ml AChE, and 5 mM DTNB solution, as well as the various concentrations of SC (1–4μM) and galantamine (1, 3, and 5μM) [20, 21] in each well. Before placing the plate in a microplate reader, the appropriate levels of ATCI (0.1–1 mM) were added and absorbance was kinetically followed at 412 nm and 37°C. Additionally, galantamine and buffer solution at pH = 8 were utilized instead of the SC as the positive and negative controls, respectively.
Evaluation of secondary structure by using circular dichroism (CD)
First, 0.2 mg/ml AChE and 0.4 mM substrate (ATCI) were prepared for studying secondary structure, and galantamine and SC were used at their maximum inhibitory concentrations. CD device was set in the wavelength range of 190–260 nm. Further, circular dichroism nerve network software (CDNN) was applied to calculate the percentage of α-helix and β-sheet in the enzyme structure in the presence and absence of substrate, galantamine, and SC. In this program, the number of amino acids in the enzyme was considered 2156 aa, and its molecular weight, concentration, and cuvette length were 237160 Da, 0.2 mg/ml, and 0.1 cm, respectively.
Examination of AChE-SC interaction through employing docking method
The interaction between the SC and AChE molecules was evaluated through employing molecular docking analysis. The 3D structure of the enzyme molecule was extracted from the protein data bank (https://rcsb.org/) (PDB code: 1EVE). In addition, the SDF file of galantamine (PubChem CID: 9651) was obtained from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). The molecular structure of the SC was drawn and 3D optimized by using HyperChem program version 8. Further, AutoDock tool version 1.5.6 was utilized for docking AChE molecule. The points of grids were 35Å × 35Å × 35Å (x, y, and z) with a grid spacing of 1Å through AutoDock Vina program. Regarding the docking process, the ‘number of a generation’ declined to 200 conformers for SC. A structure with the least energy content and best binding site was selected. Furthermore, VMD version 1.9.3 program was used to prepare the 3D schematic diagrams of the docking model to represent the different interaction types between AChE and SC.
Molecular assessment by using molecular dynamics (MD) simulation
MD simulations were performed for both AChE molecule and its complex with SC. Additionally, further analyses were carried out by using GROMACS simulation software package (version 5.1.4) on a CentOS Linux system by applying GROMOS96 43a1 force field [22–25]. The modeled enzyme molecule was solvated in a cubic box with 1 nm of solvent on all sides based on the simple point-charge (SPC216) water model. Further, genion tool was utilized and all systems were neutralized by adding Cl– and Na+ counter ions to replace initial SPC water molecule. The systems were primarily energy-minimized through a total of 5000 steps of calculations [26]. Then, the system was equilibrated for 100 ps isochoric-isothermal (NVT) and isothermal-isobaric (NPT) ensembles under the conditions of position restraints for heavy atoms. All simulations were run for 100 ns, and atomic coordinates were saved every 2 ps [27]. Finally, the results of the generated files were analyzed by using the standard software presented by GROMACS 5.1.4 [28].
Analysis of MD trajectory
The relative analysis of structural deviations in AChE and its complex was calculated in this study. The gmx rmsf, gmx rms, gmx hbond gromacs, and gmx gyrate tools were respectively used for root-mean-square fluctuation (RMSF) root-mean-square deviation (RMSD), and hydrogen bond analysis, as well as the radius of gyration (Rg) one.
Statistical measurements
All data were expressed as mean±SEM. Each of the treatment groups was compared with enzyme group through employing one-way ANOVA, repeated measure analysis, and Tukey’s post-hoc test. All analyses were carried out in Microsoft Excel 2016 (Microsoft Corporation, Redmond, WA, USA) and SPSS version 16 statistical software (SPSS Inc., Chicago, IL, USA). A statistically significant difference was set at p < 0.05. Each experiment was performed in triplicate.
RESULTS
AChE activity
AChE activity was measured according to Ellman’s method. In addition, enzyme studies started by drawing a saturation curve (Michaelis-Menten plot) of AChE by using acetylthiocholine as substrate (Fig. 2a). The Michaelis behavior of AChE kinetics should be ensured before calculating Michaelis constant (Km). Eadie-Hofstee plot is useful to check the kinetic behavior of enzymes graphically. As shown in Fig. 2b, AChE fails to represent Michaelis kinetic behavior due to non-linearity. The curvature to the right is known as a characteristic of the positive homotropic behavior of the enzyme [29]. This behavior of the enzyme reflects that thioacetylcoline improves the enzyme tendency to substrate at its high level and plays an activating role. It is a molecular mechanism to modulate the enzyme activity and substrate concentration, similar to the molecular allosteric mechanism in the hemoglobin molecule. Thus, the Hill plot and its equation were used to determine the amount of this behavior (Fig. 2c). The slope (nH = 1.33) confirmed non-Michaelis behavior with a positive homotropic effect. The S1/2 was obtained based on the Hill plot for 0.22 mM. S1/2 is considered as the level of acetylthiocholine at which the rate of AChE activity is half of the maximum velocity (Vmax). Indeed, it is equal to Km in enzymes with non-Michaelis behavior [29].

Kinetics of AChE activity, Kinetics of AChE activity in the absence of inhibitors, (a) Michaelis-Menten plot, (b) Eadie-Hafstee plot, (c) Hill plot, (d) Michaelis-Menten plot in the absence and presence of galantamine (Gal) (1, 3 and 5μM). Michaelis-Menten plot in the presence of galantamine at 1μM (e), 3μM (f), 5μM (g), Eadie-Hafstee plot in the presence of galantamine at 1μM (h), 3μM (i), 5μM (j). Hill plot in the presence of galantamine at 1μM (k), 3μM (l), 5μM (m), (n) Michaelis-Menten plot in the absence and presence of synthetic complex (SC) (1, 2, 3 and 4μM). Michaelis-Menten plot in the presence of SC with concentrations of 1μM (o), 2μM (p) and 3μM (q). Eadie-Hafstee plot in the presence of SC at 1μM (r), 3μM (s), 5μM (t). Hill plot in the presence of SC with concentrations of 1μM (u), 2μM (v) and 3μM (w).
AChE kinetics in the presence of inhibitors
Currently, galantamine is one of the important inhibitors of AChE, which is used as a drug for AD treatment [14]. In the present study, galantamine was considered as a positive control of AChE activity inhibition. The galantamine was evaluated at 1, 3, and 5μM concentrations in the AChE saturation curve [20, 21]. The plot exhibited the inhibition of the enzyme activity by all three levels of galantamine (Fig. 2d). A rise in galantamine concentration led to a reduction in the sigmoidal shape of AChE curve and conversion to the linear form (Fig. 2e–g).
As displayed in the Eadie-Hafstee plot in Fig. 2h–j, a non-Michaelian and positive homotropic behavior is observed for the enzyme in the presence of galantamine. The slopes of the Hill plots (Fig. 2k–m) were greater than the unit, which confirms the positive homotropic effect for the enzyme in the existence of galantamine.
Further, the inhibition of the SC was examined to specify the type of enzyme behavior. Based on the results, the SC at different concentrations inhibited AChE strongly (Fig. 2n) in a concentration-dependent manner. Furthermore, the sigmoidal shape of the AChE saturation curve enhanced by improving the SC level (Fig. 2o–q). This observation was confirmed by the type of curvature in the Eadie-Hafstee plot (Fig. 2r–t). The increasing SC concentration made the enzyme intensify its positive homotropic behavior. Similarly, a severe increase and decrease were respectively found in the Hill coefficient (nH) and S1/2 following a rise in the SC level (Fig. 2u–w). Figure 3 depicts the effects of the SC and galantamine with various concentrations on nH and S1/2.
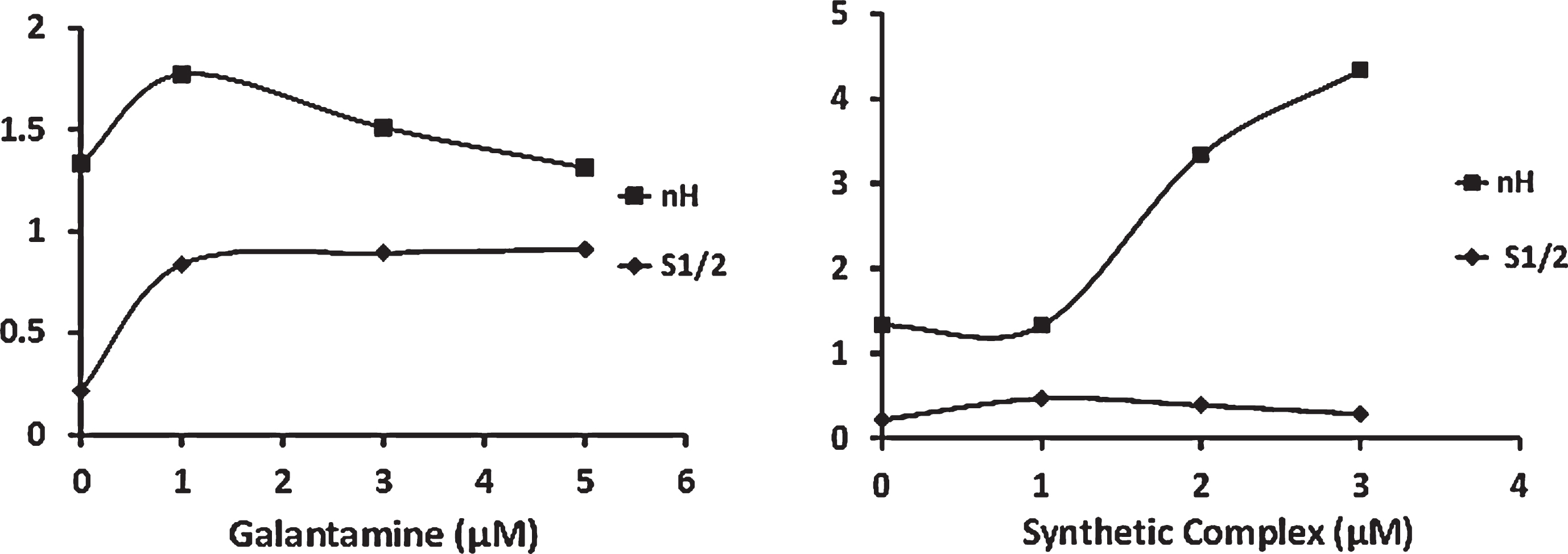
The effect of different concentrations of galantamine (Gal) and synthetic complex (SC) on Hill coefficient (nH) and S1/2 . The Hill coefficient and S1/2 diagrams show the ratio of the enzyme hemotropic behavior and cooperative effect in the presence of galantamine to the synthetic complex. Numerical information of this figure is in the diagram.
AChE enzymatic activity was measured to assess the simultaneous effect of galantamine and SC (Fig. 4). The results indicated a decrease in AChE activity by galantamine and SC (20 and 25% inhibition, respectively). Surprisingly, the activity promoted in the presence of both inhibitors compared the existence of each inhibitor alone. However, no significant difference was obtained in comparison to the activity without inhibitor (p > 0.05).

The inhibitory effect of two inhibitors galantamine (Gal) and synthetic complex (SC), we selected the maximum rate of enzyme activity at a concentration of 4μM SC and a concentration of 3μM gal. These concentrations have the greatest inhibitory effect. We used them according to Fig. 2d and 2n for comparison in the form of column graphs. There is no significant difference the enzyme activity in the presence of both Gal-SC inhibitors against the enzyme alone (p > 0.05), while the p-values are significant in both groups: Enz-Gal in opposition to the enzyme without inhibitor and also Enz-SC against enzyme alone (p < 0.05). The star indicates the difference between the two inhibitors and the enzyme alone. It also shows significance. The parts that do not have any star indicate that there is no significant difference.
Effect of inhibitors on AChE structure
Considering the significant effect of the two inhibitors on AChE catalytic activity, the present study evaluated a probable structural change on the enzyme molecule in the presence of the inhibitors through employing CD spectroscopy. It seems that galantamine led to more changes in the spectrum than the SC in Fig. 5a.
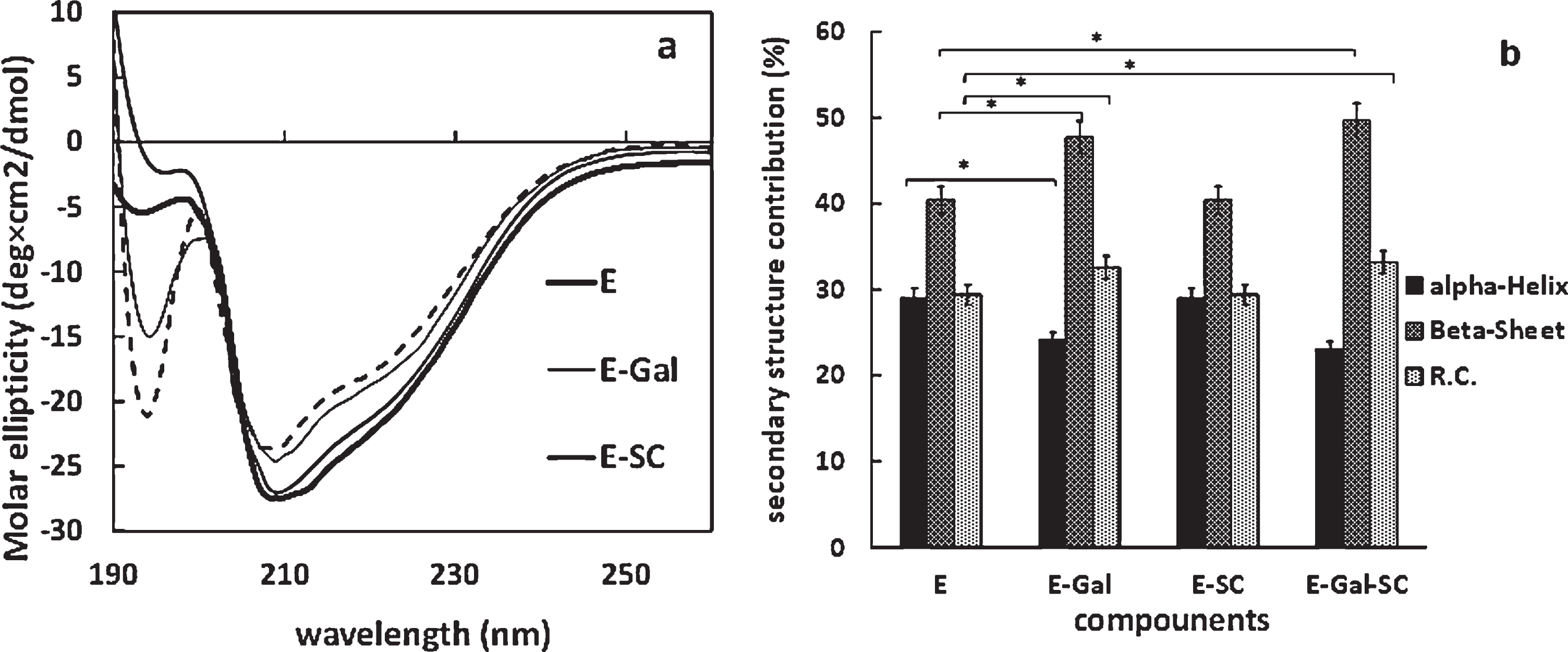
Effect of inhibitors on the structure of AChE (a) CD spectra of AChE in the presence and absence of substrate, galantamine (Gal), synthetic complex (SC) and their various complexes in the wavelength range 190 to 260 nm. (b) The percentages of alpha-helix and beta-sheet and random coil (R.C.) in the structure of AChE in the absence and presence of inhibitors galantamine (Gal) and synthetic complex (SC) (p < 0.05).
The change in secondary structures in the presence of each inhibitor and both is presented in Fig. 5b for further analysis. Following the use of galantamine, the α-helix and β-sheet content of the AChE molecule declined (23%) and improved (47%) significantly, respectively. Surprisingly, the SC had no significant effect on the structured content (p > 0.05). However, AChE molecule represented structural changes in the existence of both inhibitors, similar to that observed in the presence of galantamine alone (Fig. 5b).
Docking of AChE with complex
To survey the interaction between galantamine and AChE, AutoDock software was applied to determine the possible binding sites. Based on the results in Fig. 6a, galantamine bound to AChE and was surrounded by Tyr70, Asp72, Tyr121, Glu270, Trp279, Leu282, Phe284, Asp285, Ser286, Phe290, and Tyr334. This drug could penetrate AChE with a higher binding free energy than for the SC (–8.9 kcal/mol).

Schematic structure of 1EVE and SC. Docking enzyme simulation in the presence of inhibitors with VMD software (a) Galantamine -Enzyme (binding energy is –8.9 (kcal/mol)) (b) Synthetic complex-Enzyme (Binding energy is –7.6 (kcal/mol)). (c) Galantamine- Synthetic complex -Enzyme (Binding energy is –7.8 (kcal/mol)).
Figure 6b demonstrates the interaction between the SC and AChE. The results reflected that the complex was bound to the different binding sites of AChE compared to the galantamine. Additionally, the molecule was bound to the hydrophobic core of AChE. It was surrounded by the Trp84, Pro86, Leu127, Val129, Tyr442, and Glu445 which are involved in the hydrophobic forces. The SC was located in the long channel of the active site interacting with the mentioned residues (Fig. 6b). The aromatic structure of the SC was surrounded by the phenyl ring of Tyr442 and indole one of Trp84 in appropriate orientation for binding via π - π stacking interaction. Along with the hydrophobic interactions, several amino acids participated in the active site of the enzyme to form the strong interactions between the SC and AChE. The results suggested that the SC was bound to AChE and exhibited higher interaction affinity. This molecule could penetrate AChE with a suitable binding free energy (–7.6 kcal/mol). In order to examine the triplex complex between the two inhibitors and enzyme, AChE-SC complex was docked in the presence of galantamine (as experimental order). The results indicated the bonding of galantamine to a binding site different than the previous one. In this new binding site, ser79, Trp84, Val129, Val431, Tyr442, and Glu445 surrounded galantamine (Fig. 6c). The binding free energy of two inhibitors and AChE was –7.8 kcal /mol.
MD simulation analysis
MD simulation is considered as a more valid method for supporting experimental results in protein-ligand complex based on the structural aspects at the atomic level. The results can represent theoretical knowledge about the effects of the SC (ligand) binding AChE (protein) through structural features. The MD simulation on AChE and ligand-protein types revealed the structural behavior of the enzyme in a dynamic system. The simulations were carried out for both AChE and SC-AChE to recognize the structural understanding of the complex. Further, RMSD, RMSF, Rg, and intermolecular hydrogen bond were utilized to explore the fluctuations caused by the SC binding during simulation.
In the MD studies, AChE was inserted to galantamine. The mean RMSD was 0.24 nm, and 10 peaks were reported for RMSF in the range of residue 45, 130, 200, 70, 150, 195, 230, 250, 350, and 490. Furthermore, Rg reduced from 2.32 to 2.27 nm and two intermolecular hydrogen bond were detected [30].
RMSD is often used for calculating the mean distance between the atoms (commonly backbone ones) of two superimposed proteins and valuating the similarity between two globular protein structures [27]. The RMSD plot related to AChE and AChE-SC structures exhibited a convergence pattern during 100-ns simulations (Fig. 7a). Considering the structures, the mean RMSD was respectively determined 0.249±0.01648 and 0.205±0.0237 nm during 100 ns. A convergence was obtained for both AChE and complex structures at the end of 100-ns simulations, which confirmed stable trajectory. Despite the stability of both enzyme and SC-AChE, the RMSD of SC-AChE changed and was different.

Functional effects of AChE binding with SC (a) RMSD plot of the AChE and AChE-SC in 100 ns of simulation. Average of RMSD of the AChE (Blue) and AChE-SC (Red) (b) RMSF plot of the AChE and AChE-SC in 100 ns of simulation. Average of RMSF of the AChE is (Blue) and AChE-SC (Red). The maxima fluctuations of both systems, shown in the resid 74–86, 279–290 and 338–340. (c) Number of H-bond plot. (d) Radious of gyration plot of the AChE and AChE-SC in 100 ns of simulation.
RMSF analysis is another tool to assess the dynamic stability of the system, which reflects the flexibility of a certain residue [27]. The dynamic behavior of residues due to ligand binding was determined by using RMSF based on the C-alpha atoms. RMSF curve was plotted for AChE and its complex structures (Fig. 7b). During 100 ns, the mean RMSF was equal to 0.1072±0.055 and 0.0109±0.0515 nm according to AChE and SC-AChE structures, respectively. The fluctuation between AChE and AChE-SC was observed in resid 74–86 and 279–290 during the simulation. The mentioned sites located in the active sites underwent acylation processes.
An intermolecular hydrogen bond is critical in protein stability, as well as interacting with protein and solvent. Regarding this study, the number of the hydrogen bond was calculated for AChE and complex types. The results indicated the existence of a hydrogen bond in AChE-Gal, and its absence in the AChE-SC. As displayed in Fig. 7c, the number of the intramolecular hydrogen bonds between amino acids is less in AChE-SC than the AChE during MD simulation. However, no significant difference is found between the two materials in terms of the hydrogen bond plot.
The Rg of a protein is defined as the root-mean-square-distance from each of its atoms to their centroid (Fig. 7d). The AChE structure became unfolded during 60 ns, while it was refolded at the end of the simulation in the presence of SC. Rg is considered as an indicator unfolding (disclosing) protein structure.
DISCUSSION
AChE is functionally unique due to its special structure. The molecule involves the two important sites of active and peripheral anionic [10]. According to Dvir et al. [4], substrate (acetylthiocoline) is transiently attached to the peripheral anionic site in the first step. Therefore, the inhibition of the substrate is observed at its higher levels, while the enzyme activity accelerates in lower concentrations. The previous studies have suggested the existence of at least one other binding site other than the active one on AChE structure for the substrate, which exhibits a regulatory effect [31]. In addition, the rate of enzyme activity gradually increases by raising the substrate level. This issue is called positive homotropic effect [14]. The present study analyzed the kinetics and behavior of AChE through enzymology for the first time. The results reflected that AChE consisted of two binding sites for the substrate, which are consistent with those of the previous research [4]. Thus, it is expected that the enzyme does not have Michaelis behavior, to which the researchers did not pay attention. The enzymological study demonstrated the non-Michaelis behavior of the enzyme, and consequently the sigmoidal shape of curve. The Eadie-Hofstee plot and Hill plot equation revealed that AChE belonged to the allosteric enzymes and represented a non-Michaelis behavior. The Hill coefficient, as the most important measurement index, was obtained 1.33 in the absence of inhibitors. Despite the smallness of the coefficient, it indicated a low positive cooperative effect. In other words, the enzyme tendency to the substrate enhanced by promoting the substrate concentration [29]. The slope of the Hill plot related to AChE exhibited a non-Michaelis behavior. Accordingly, S1/2 (equivalent to km) was applied in this behavioral model. Indeed, S1/2 is the concentration of acetylthiocholine at which the enzyme level reaches half of Vmax [29]. Based on the results of the present study, the enzyme behavior cannot be determined through the Michaelis model and Lineweaver-Burk plot when the behavioral model of AChE is non-Michaelis, which is in line with those of the previous research [32].
Galantamine is one of the safest drugs for AD patients [16]. It is the only acetylcholinesterase inhibitor shows a significant reduction in the risk of dementia. Over time, galantamine reduces mortality more than other inhibitors, and because it performs better in terms of cognitive effects and ranks first [33]. In this study, we study evaluated AChE behavior in the existence of galantamine as a positive control. Galantamine is a traditional drug [3], which the different types of its inhibition such as mixed [34] or competitive [32] have been reported in the previous studies. However, the results of the present study represented the non-Michaelis effect of galantamine on AChE by considering the enzyme behavior. Further, the docking analysis with the enzyme according to Atanasova revealed the bonding of galantamine to another site of the active site [35], as well as its allosteric behavior in nicotinic acetylcholine receptors [36]. The CD and docking results proposed the change in AChE structure, and consequently an increase in S1/2 as one of the inhibitory mechanisms of galantamine. Given that S1/2 is equivalent to km, galantamine diminished the effect of substrate affinity for the enzyme by raising S1/2. Based on the kinetics of Eadie-Hofstee plot, the non-Michaelis behavior of AChE decreased and inhibited by enhancing galantamine concentration.
The SC is known as a cobalt (III) Schiff base complex with biological significance such as antibacterial activity [17]. The present study evaluated the inhibitory effect of the SC on AChE and its potential as a drug for the first time.
The results demonstrated the effect of the SC on the kinetic behavior of AChE which was non-Michaelis and a promotion in its level increased inhibitory intensity. Indeed, the inhibitory effect of the enzyme is associated with an improvement in Hill coefficient, which reduces S1/2 and AChE activity. This inhibitory power of the SC can be attributed to the presence of three pyridine rings with quaternary amine, a methoxy group in the benzene ring, and two phenyl rings [37–39]. Chigurupati et al. outlined high inhibitory activity and binding to the active site in the compounds containing methanone, phenyl, indole ring, and pyrazole or a simple aryl group such as benzene [39]. However, the SC interacts with the phenyl ring of Tyr442 and indole one of Trp84 at the peripheral anionic site and helps inhibit the enzyme. In fact, the amino acid Trp84 is involved in regulating the allosteric changes in AChE, as well as the correct orientation of the substrate [40]. The SC may represent a higher inhibitory intensity than the galantamine because of binding in this region, especially interacting with Trp84. Katrina referred to the existence of methoxy group, as well as the appropriate distance between functional groups as the factors influencing the allosteric inhibition of AChE [41], which the SC had these properties. However, Abdul Manap [42] introduced the binding of pyridine ring to Trp86 as one of the reasons for the ability of the inhibitors to bind to both sites. Based on the CD and docking results, at least one of the inhibitory mechanisms of the SC was through raising Hill coefficient. Thus, the enzyme structure changed without varying the percentage of alpha and beta.
While the different appearance of the enzyme in the presence of two inhibitors, the p-value between the enzyme and the Enz-Gal-SC does not show a significant difference. Therefore, due to its large load error, there is no claim to increase enzyme activity in the presence of two inhibitors. It is possible that the interaction effect of gal and SC caused the inhibitory effect of each other to be neutralized, resulting in no effect of enzyme inhibition. Perhaps the effect of these two inhibitors is in a position of the enzyme where an increased increase in AChE activity is observed.
We examine the second structure in Fig. 5b, which is a solid component of a protein structure. Although the secondary structure of the enzyme was relatively unaffected by the synthesis of the synthesis; We show the changes in the spatial structure of the enzyme in Fig. 7, so that the SC interacts at several points when exposed to the enzyme (Fig. 7b). According to Fig. 7c, the enzyme is unfolded and refolded after encountering SC, and this folding remains stable. The SC binds to the hydrophobic region, but the large changes in the secondary structure of the enzyme in the presence of the galantamine as shown in Fig. 5b are probably due to the hydrogen bonding that occurs at the enzyme activity site. In general, we expected that any change in enzyme behavior would occur following a change in its structure [43]. There is currently information that the effectors often attach to the enzyme but do not necessarily change its structure [44].
Furthermore, MD results indicated the interaction between the enzyme and SC molecules, leading to the enzyme stability while remaining the SC attached to AChE. RMSF curve reflected the involvement of amino acids Pro86, Leu127, Val129, Tyr442, and Glu445 in this interaction, along with Trp84. The RMSF results are consistent with the docking ones which suggested Trp84 as an important component of AChE. Some researchers have pointed out that the inhibitor interaction with this amino acid plays a critical role in changing the behavior of the enzyme [40]. However, no significant difference was obtained in the number of hydrogen bond to enzyme interactions in the absence or presence of the SC. This issue is ascribed to the existence of hydrophobic bonds in the interaction between the SC and enzyme, which was well observed in their docking. Additionally, the results of Rg variations in the SC-AChE demonstrated the greater stability of the enzyme protein structure after binding to the inhibitor, as well as preventing the enzyme separation and unfolding by the SC. Accordingly, the enzyme can be more affected by the inhibitor. Regarding AChE activity, Roca confirmed the mechanism of allosteric inhibition by binding inhibitors to the peripheral anionic site and proposed the cooperative effect of some inhibitors on the enzyme. They found a non-classical effect on cell recognition and protein adhesion process for the inhibitors interacting with the peripheral anionic site, along with affecting the allosteric activity of the enzyme. In fact, the peripheral anionic site exhibits a non-cholinergic enzymatic function which may be involved in producing and storing beta-amyloid plaques in AD patients [45].Finally, the ability of the SC to reduce beta amyloid should be evaluated in future since extensive studies have been conducted on drugs with this ability.
Conclusions
In the present study, the inhibitory effect of the SC ([CoIII(L1)(py)3]ClO4) on AChE was compared with that of AD drug (galantamine). In fact, this kinetic enzyme study was first performed using several enzymatic methods in terms of Michaelian and homotropic behavior extensively. The kinetic results confirmed those of the previous studies on AChE structure. AChE represented a positive homotropic behavior in the presence of the SC, as well as in the existence of the substrate alone. Due to the regulatory effect of the substrate on a position other than the active site, even in the presence of inhibitors, a rise in its level increased the tendency of AChE. Further, the inhibitors altered the enzyme protein structure, changed the activity, and inhibited AChE. However, the SC had more inhibitory effect than the galantamine, which was elucidated by the results of docking and MD analyses. The results reflected that the SC binds to the hydrophobic region in the active site pathway and affected the allosteric behavior of AChE by influencing Trp84. Therefore, the SC is involved in regulating orientation and directing the substrates to the active site. Finally, an enhancement in the SC concentration resulted in decreasing the activity of AChE as one of the target enzymes in treating AD and reducing its progression.