
Abstract
Although Alzheimer’s disease is most often studied in terms of memory impairments, olfactory dysfunction begins in the early stages. We tested olfactory learning, sensitivity, and response bias using signal detection methods in 12-month-old male and female 5xFAD mice and their wildtype controls in the operant olfactometer. Odor detection was not reduced in the 5xFAD mice, but learning was, which was worse in female 5xFAD mice than in males. Female mice were more conservative in their response strategy. Signal detection analysis allows us to discriminate between cognitive and sensory deficits of male and female mouse models of AD.
Keywords
Introduction
Although the most salient cognitive symptoms of Alzheimer’s disease (AD) are learning and memory deficits [1–3], one of the earliest symptoms is a decline in olfactory function [4–6], which has been shown to be a better predictor of AD onset than loss of verbal episodic memory [5]. This predictive property of olfactory dysfunction makes the olfactory pathway a worthy target for diagnosing AD and studying its progression. However, the distinction between healthy aging and AD, in terms of olfactory deficits, is complex [7–9]. Olfactory deficits have been observed in more than half of the population over 65 [10, 11] including those with mild cognitive impairment, AD, and other types of dementia, along with cognitively healthy individuals [7, 12–15]. Age-related decline in odor identification has been correlated with a faster rate of cognitive decline and reduced volume of the hippocampus, entorhinal, fusiform, and middle temporal cortices [16]. Thus, the presence of olfactory learning and identification deficits may be an important feature differentiating AD phenotypes from healthy aging.
Investigating olfactory deficits in mouse models of AD can help us understand the effects of AD related mutations in these mice [17]. Both the Tg2576 and the APP/PS1 mice have olfactory deficits accompanied by amyloid-β (Aβ) pathology in the olfactory pathways [18, 19]. Olfactory deficits have also been reported in tau transgenic mice [20] and in the 3xTg-AD mouse, which carries both Aβ and tau pathology related transgenes [21, 22]. On the other hand, the 5xFAD mice showed no deficits in odor detection or olfactory working memory in the operant olfactometer [22–24] and, despite significant Aβ pathology in the olfactory bulb, 5xFAD mice did not show age-related deficits in olfactory memory in a Pavlovian conditioned odor preference task [25]. While the conditioned odor preference task is valuable for testing long-term odor memory [26], it does not provide a measure of olfactory learning as is obtained from the operant olfactometer [22, 24]. Therefore, the aim of the present study was to examine olfactory sensitivity and learning in the operant olfactometer in 12-month-old 5xFAD mice. This timepoint was selected as the 5xFAD mice show abnormal whisking behavior [27, 28], impaired hearing [29], cognitive deficits [30–32], motor deficits [33, 34], and metabolic deficits at this age, leading to increased frailty scores and early death [35, 36].
Methods
Subjects
Male 5xFAD mice (B6SJLT-Tg (APPSwFlLon, PSEN1*M146L*L286V) 6799Vas/Mmjax) were purchased from Jackson labs (Bar Harbor, ME, USA; stock #034840) and bred with female B6SJLF1/J mice (Jackson labs; stock #100012), and the offspring were tested. The 5xFAD mouse carries three human APP mutations (Sweden, London, Florida) and two presenilin 1 mutations (M146L and L286V), causing accelerated Aβ42 accumulation [37]. Five male and 8 female WT, and 5 male and 9 female 5xFAD mice at 12 months of age were tested. Mice were naive to behavioral testing. Pups were weaned at 21 days of age and housed in same sex groups of 2–4 in transparent polyethylene cages (35×12×12 cm) with ad libitum food (Purina Rodent Chow #5001) and tap water. Housing cages contained pine chip bedding and a polyvinyl chloride tube (5 cm diameter, 8 cm long) for enrichment. The housing room was on a 12:12 h reversed light/dark cycle. Mice were genotyped for the APP and PS1 transgenes using polymerase chain reaction by Dr. Chris Sinal (Department of Pharmacology, Dalhousie University). All test procedures were approved by the Dalhousie Committee on Animal Care (Protocol #14-059).
Apparatus
Liquid dilution olfactometers (Knosys Olfactometers Inc.) previously described [22, 38] were used. Air was sent through a filter after which it was split into two pathways, one as clean air, and the other flowed through a manifold which controlled the air flow through saturation bottles and into a T-junction, where clean and odorized air flows converged. The odor sampling port contained an infrared beam to detect nose-pokes, a reinforcement tube delivering the reward, and a sensor that detected when the mice were licking the tube.
Odors
Ethyl acetate was used as the odorant as it has commonly been used in olfactory detection tasks in mice [22, 40]. Vapor concentrations in the odorant bottles used were 1, 0.1, 0.01, 0.001, 0.0001, and 0.00001 ppm. The vapor concentration presented to the mice in the odor sampling port was approximately 5% of the concentration in the odorant bottles [38]. All odors were diluted with heavy mineral oil.
Water restriction
Ten days prior to the start of testing mice were individually housed and placed on water restriction. Mice were weighed daily and given measured amounts of mash (powdered food pellets mixed with water) to maintain their weight at approximately 85% of free feeding weight.
Behavioral testing
All behavioral testing was done during the dark phase of the light/dark cycle. The mice were initially trained for 20 trials to lick the reinforcement tube to receive a water reward and were rewarded for simply licking the reinforcement tube. The inter-trial interval increased from 0.1 s to 12 s over the 20 trials. During the next stage of training an odor was introduced and the mice were required to keep their head in the odor sampling port while the final valve diverted the odor into the port. The amount of time the mice were required to keep their head in the port increased from 0.1 s to 1.1 s over 120 trials. This training was completed when the mice performed 20 trials with the final valve on for 1.1 s. During the testing phase the mice were presented with a stimulus odor when they inserted their head into the odor sampling port, either a rewarded stimulus (S+) consisting of air pumped through the ethyl acetate odorant bottle, or an unrewarded stimulus (S–) of air pumped through an odorant bottle containing mineral oil. When the mice were presented with the S+, they were rewarded for licking the reinforcement tube. Trials were initiated by the mice poking their nose into the odor sampling port, with a minimum inter trial interval of 4 s. The mice were presented first with the highest concentration of ethyl acetate (1 ppm) and given 5 blocks of 20 trials, each consisting of 10 S+ trials and 10 S–trials presented in a random order. They were then tested on the remaining concentrations in descending order. They received 100 trials over 5 blocks on each odor concentration except for the lowest concentration (0.00001 ppm) on which they received 200 trials over 10 blocks. The mice received 100 trials per day, with the lowest concentration tested over two days. These extra blocks on the lowest concentration provide a period of stable performance for the signal detection analysis.
Statistical analyses
R version 4.1.1 [41] was used for all analyses, using the “nlme” [42] and “MuMln” [43] packages. To determine if there were differences in odor sensitivity, the sensitivity index (d’) for each mouse on the last five blocks of the lowest odor concentration were calculated and compared with linear models. This measures the difference between the distributions of the signal and noise, with a larger d’ indicating the signal was more readily detected. The d’ value was calculated by subtracting the z score that corresponded to the false-alarm rate from the z score that corresponded to the hit rate [44]. To examine differences in response strategy, the response bias (c) for each mouse on the last 5 blocks of the lowest odor concentration were calculated and compared with linear models. This measures the general tendency to respond on any given trial and was calculated by taking the negative mean of the z score that corresponded to the hit rate and the z score that corresponded to the false-alarm rate [44]. The last 5 blocks were used as these signal detection measures assume stable performance. Overall performance and learning were investigated by comparing the accuracy of the mice with linear mixed effects models. Models were compared using Akaike Information Criterion corrected for small sample size (AICc) [45]. All possible valid models, including null models without any effects, were calculated and the model with the lowest AICc was reported. The use of AICc allows for different models to be compared, and the model which best fits the data to beselected.
Results
5xFAD and WT mice do not differ in olfactory sensitivity
To determine if there were differences in olfactory sensitivity, the sensitivity index (d’) on the last five blocks of the lowest odor concentration were compared. The null model (AICc = 80.7, weight = 0.459) best explained the data, indicating no differences in the ability of the mice to detect this odor (Fig. 1).
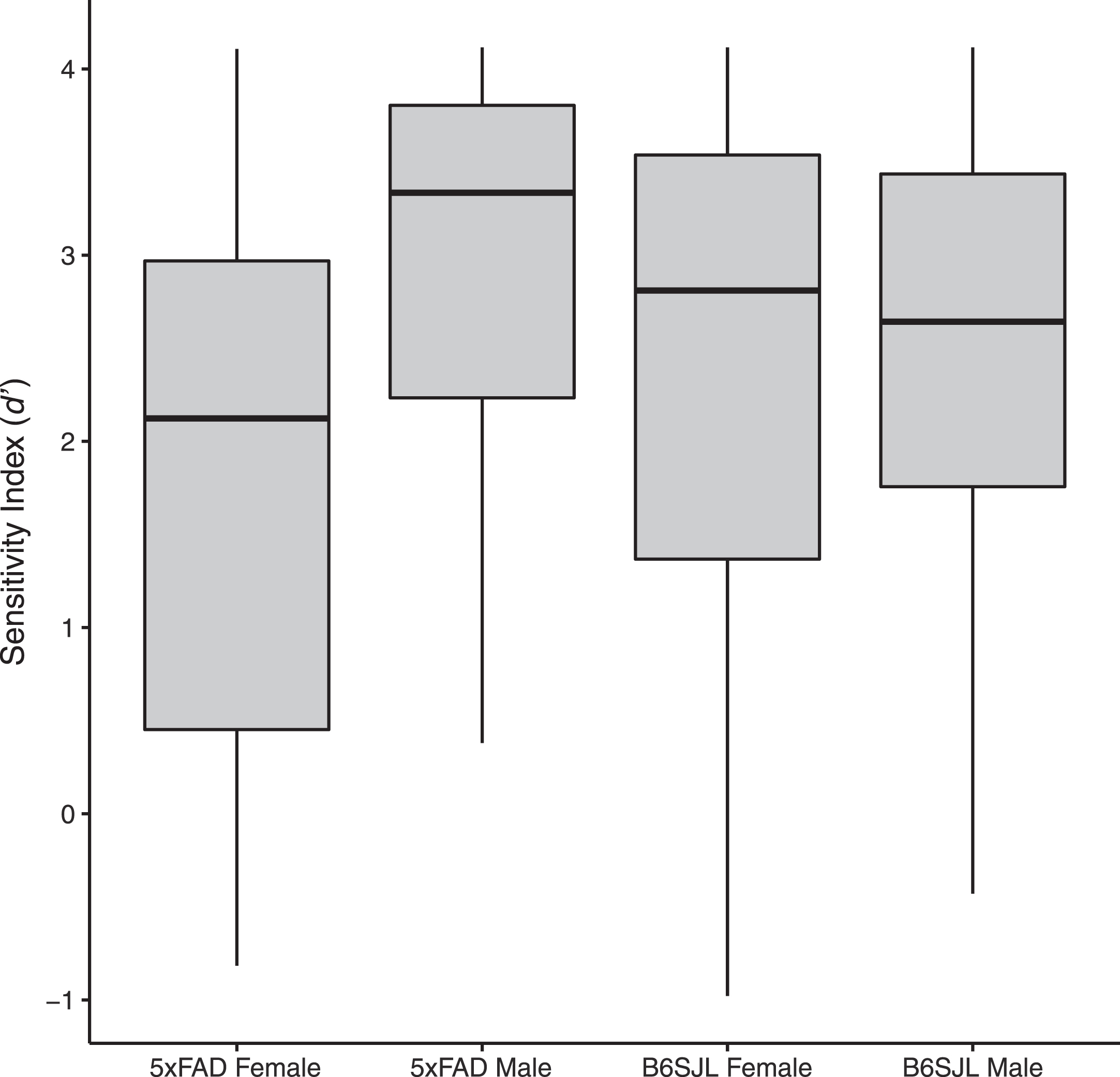
Boxplots, showing the median, 25th, and 75th percentiles, with whiskers extending to the furthest points within 1.5 interquartile ranges, and outliers shown as dots, for the sensitivity index (d’) on the last five block of trials for male and female 5xFAD and WT (B6SJL) mice on the lowest odour concentration (0.00001 ppm ethyl acetate). There were no significant differences between groups.
Female mice are more conservative in their responses than males
To determine if there were differences in response strategies used by the mice, their response bias (c) on the last five blocks of the lowest odor concentration were compared. The model with a main effect of sex (AICc = 55.7, weight = 0.685) best explained the data, with the female mice (c = –0.285±790) having a more conservative bias than the male mice (c = –0.468±0.407; Fig. 2). This model differed significantly from the null model (AICc = 60.3, weight = 0.069, p = 0.011).

Boxplots, showing the median, 25th, and 75th percentiles, with whiskers extending to the furthest points within 1.5 interquartile ranges, and outliers shown as dots, for response bias (c) on the last five block of trials for male and female 5xFAD and WT (B6SJL) mice on the lowest odor concentration (0.00001 ppm ethyl acetate). The female mice had a more conservative response bias than the male mice (p = 0.011) and there was no effect of genotype.
5xFAD mice show olfactory learning deficits
To determine if there were differences in learning rate, the accuracy levels across all blocks were compared. The model with main effects of genotype, sex, odor concentration, and block, as well as genotype by sex, genotype by block, sex by block, sex by odor concentration, and block by odor concentration interactions (AICc = –613.2, weight = 0.141) best explained the data and differed significantly from the null model (AICc = –358.0, weight < 0.001, p < 0.0001). The overall trend is that the WT mice had higher accuracy (0.83±0.20) than the 5xFAD mice (0.81±0.21), and male mice had higher accuracy (0.85±0.18) than female mice (0.79±0.22). When splitting the data by genotype, there was a main effect of sex in the 5xFAD mice, with the males having higher accuracy (0.88±0.16) than the females (0.76±0.22), but no such difference in the WT mice. Additionally, the main effect of block was significant for the WT (p = 0.03) but not for the 5xFAD mice (p = 0.45), suggesting a lower effect of learning, or relearning, the task across blocks in the 5xFAD mice. Accuracy increased as the mice progressed through blocks (Fig. 3), and this increase was most pronounced on the first odor concentration (1ppm) with accuracy on the first block (0.52±0.09) being much lower than the final block (0.83±0.15), and less pronounced on the final odor concentration (0.00001 ppm) going from 0.77 (±0.21) on the first block, to 0.87 (±0.20) on the second block, to 0.88 (±0.16) on the final block. The males (0.56±0.14) had lower accuracy than the females (0.76±0.19) on the first odor concentration (1 ppm), but males had higher accuracy on all other concentrations. There is a trend where on the first blocks of each odor concentration the difference in accuracy between the males (0.75±0.19) and females (0.66±0.21) was greater than the difference between the males (0.89±0.15) and females (0.86±0.19) on the fifth blocks. This larger difference between performance on the first and fifth blocks suggests the females were slower than the males to relearn the task with each new odor concentration.

Mean (±95% confidence interval) accuracy of female and male 5xFAD and WT (B6SJL) mice on each block of 20 trials for each of the six odor concentrations. Overall, the wildtype mice had higher mean accuracy (0.83±0.20) than the transgenic mice (0.81±0.21), and male mice had higher mean accuracy (0.85±0.18) than female mice (0.79±0.22). Wildtype mice had a greater effect of block than transgenic mice, and accuracy was greatest overall (0.91±0.13) on the third odor concentration (0.01 ppm ethyl acetate).
Discussion
We studied odor detection and olfactory learning in 12-month-old 5xFAD mice. Our results indicated that: the 5xFAD mice showed deficits in olfactory learning at 12 months of age and this deficit is greater in female mice than male mice; the 5xFAD mice did not have deficits in olfactory sensitivity; and the 5xFAD mice did not use a different strategy than WT mice to learn the odor detection task, but the female mice of both genotypes were more conservative with their responses than males. The operant olfactometer does not rely on auditory or visuospatial cues, nor on motor performance, which makes it an ideal task to measure learning and memory in the 5xFAD mice, which show age related sensory [31], motor [34], and cognitive [32] deficits. While deficits in odor detection and olfactory learning in humans with AD may be confounded by loss in olfactory sensitivity [46], our results show that the 5xFAD mice do not have decreased olfactory sensitivity at 12 months of age. This agrees with our previous findings in 5xFAD mice at 6 months of age [22, 25], hence the operant olfactometer can be reliably used to study learning deficits in the 5xFAD mice. We had fewer male mice than female mice in this study. This is often an issue with studies of aged mice due to differences in mortality rates in AD mouse models [36].
Our results also showed an impairment in olfactory learning in the 5xFAD mice at 12 months of age, which agrees with [47] who showed olfactory learning deficits are present as early as 4 months of age in this model. Performance of the mice was highest on the third odor concentration tested (0.01 ppm). We propose this is because when tested at this concentration the mice had learned the task well from the prior testing, and where being presented with an odor concentration which was easily detected.
However, O’Leary and Brown [32] did not show any deficits in olfactory memory using the conditioned odor preference task. This may be due to the nature of the tests, as the conditioned odor preference task relies on Pavlovian conditioning [48], and the olfactometer uses operant conditioning [38]. Indeed, intact Pavlovian learning in the fear conditioning task has been shown in the 5xFAD mice [49], while operant conditioning is disturbed [24]. Similarly, an intact eye blink reflex has been shown in AD patients [50], yet patients show deficits in operant conditioning [51]. The female 5xFAD mice appeared to have a greater impairment in learning compared to the males, which is in line with findings of greater pathology in female 5xFAD mice compared to males [52, 53].
We also showed that female mice were more conservative in their responses. Sex differences in mouse decision making have been reported [54–56], and we have previously reported sex differences in AD mouse models [22, 58]. This further demonstrates the importance of examining sex differences when working with mouse modelsof AD.
Our results show the value of analyzing olfactory learning and memory using the olfactometer, as it is not confounded by other behavioral or sensory dysfunctions seen in the 5xFAD mice. Thus, it can be reliably used to study disease mechanisms and test novel therapeutics. The value of a signal detection analysis is that it examines the cognitive strategies used by the mice, and allows the for the discrimination between cognitive and sensory deficits in mouse models of AD.