
Abstract
Background:
Previous research suggests physical activity attenuates grey and white matter loss; however, there appears to be individual variability in this effect. Understanding factors that can influence the relationship between physical activity and brain volume may enable prediction of individual response.
Objective:
The current study examined the relationship between objectively-measured physical activity and brain volume; and whether this relationship is moderated by age, sex, or a priori candidate genetic factors, brain-derived neurotrophic factor (BDNF) Val66Met, or apolipoprotein (APOE) ɛ4 allele carriage.
Methods:
Data from 10,083 men and women (50 years and over) of the UK Biobank were used to examine the study objectives. All participants underwent a magnetic resonance imaging scan to quantify grey and white matter volumes, physical activity monitoring via actigraphy, and genotyping.
Results:
Physical activity was associated with total grey matter volume, total white matter volume, and right hippocampal volume. Only males had an association between higher physical activity levels and greater cortical grey matter volume, total grey matter volume, and right hippocampal volume. Age moderated the relationship between physical activity and white matter volume.
Conclusion:
Our results indicate that in males, but not females, an association exists between objectively-measured physical activity and grey matter volume. Age may also play a role in impacting the relationship between physical activity and brain volume. Future research should evaluate longitudinal brain volumetrics to better understand the nature of age and sex-effects on the physical activity and brain volume relationship.
INTRODUCTION
Atrophy of grey and white matter occurs as a function of aging and is accelerated in neurodegenerative conditions, such as dementia [1]. There is no effective pharmaceutical treatment for dementia or cognitive decline that alters the course of disease progression, nor that has been demonstrated to attenuate grey matter loss associated with either aging or disease. Recent updates from the Lancet Commission on Dementia Prevention [2] posits nine modifiable risk factors as contributing to dementia risk, one of which is physical inactivity. Indeed, higher levels of habitual physical activity and exercise interventions have both been shown to either attenuate age-related grey matter volume loss [3–5], or in the case of some brain regions (i.e., the hippocampus; brain region involved in learning and memory) contribute to increases in grey matter volume [6–9]. Nevertheless, there appears to be some distinct heterogeneity in study findings across this field [10–12], and individual variability in response likely influences the relationship between physical activity and brain volume [13].
Understanding moderators of the relationship between physical activity and brain volume is vital, as it may enable the identification of individuals, or groups of individuals, that gain the greatest neural benefits from physical activity [13, 14]. A moderating variable is one that changes the effect of an exposure on an outcome, i.e., a variable that influences the relationship between physical activity and brain volume. Moderation is confirmed when the relationship between two variables is different across various levels of the moderating variable. In the case of physical activity and brain volume, a number of potential moderators have been posited and investigated, including non-modifiable between-person factors such as age, sex, and genetics.
Age may influence the relationship between physical activity and brain volume, whereby an effect may only be discernible once reaching, or within, a particular age range. For example, age may influence the effects of physical activity on important mediating molecules, such as brain-derived neurotrophic factor (BDNF) [15]. Sex has also emerged as a possible moderator of the relationship between physical activity and brain health. Females tend to be disproportionately affected by Alzheimer’s disease, with females observed to have higher rates of incidence, greater accumulation of associated pathologies, and greater decreases in brain volume [16]. Based on sex differences in physiological response to physical activity [17, 18], it is possible the beneficial effects of physical activity on the brain may differ between males and females. Literature examining the relationship between physical activity and cognition indicates females typically gain more benefits from higher levels of activity [19]. Nevertheless, the examination of sex-effects in the relationships between physical activity and brain volume have not been as consistent, with some studies identifying only males [20] and others only females [21, 22] gain benefit from higher physical activity levels.
A substantial amount of literature exists to suggest that salient genetic factors play a role in moderating the relationship between physical activity and brain health. One such potential moderating factor is a single nucleotide polymorphism of the BDNF gene, termed Val66Met, which influences the secretion of BDNF: a potent neuromodulator, associated with better brain health and the levels of which are decreased in Alzheimer’s disease. Given the independent link between BDNF Val66Met carriage and smaller brain volume [23, 24], and the potent effect of exercise in increasing circulating BDNF levels [25], there is a strong theoretical basis to suggest a moderating effect of this genetic factor. Indeed, one report suggests Val/Val homozygotes, but not Met carriers, have a relationship between higher levels of physical activity and brain volume [26]. The apolipoprotein (APOE) ɛ4 allele is the greatest known genetic risk factor for sporadic Alzheimer’s disease and is also associated with swifter cognitive decline and smaller brain volume in cognitively normal older adults. To our knowledge only one study (n = 97) has examined the moderating effects of APOE ɛ4 carriage on the relationship between physical activity and brain volume, finding that only ɛ4 carriers had reduced hippocampal atrophy in relation to higher physical activity levels [4].
Previous studies of moderators of the physical activity and brain volume relationship have examined non-modifiable moderators in isolation, and/or in relatively small samples. The current study will build on prior work by utilizing data from a large cohort study (UK Biobank) to examine the moderating effects of a number of non-modifiable factors that may influence the relationship between physical activity and brain volume. We will address two specific aims: 1) Examine the relationship between objectively-measured physical activity and brain volume in the UK Biobank; and 2) Examine whether the relationship between objectively-measured physical activity and brain volume is moderated by age, sex, BDNF Val66Met, or APOE ɛ4 allele carriage.
METHODS
Participants
The UK Biobank (https://ukbiobank.ac.uk/) is a large prospective cohort study that commenced in 2006 and has collected health data on over 500,000 participants [27]. Individuals aged 40–69 years were recruited to attend one of 22 data collection sites between 2006 and 2010 for a baseline assessment. Data pertinent to the current study was collected in 2013–2015 (accelerometer data) and 2014 onward (magnetic resonance imaging; MRI, scans).
Standard protocol approvals, registrations, and consents
Ethical approval was provided by the NHS Na-tional Research Ethics Committee (11/NW/0382). The current analyses were conducted as a part of the UK Biobank application 45567. All participants provided informed consent prior to participation.
Exclusion and inclusion criteria
The dataset received from UK Biobank contained data from 502,527 participants (Supplementary Figure 1). As our research focus is understanding the relationship between physical activity and brain volume in older adults, we only included individuals aged 50 years and older at baseline (n = 384,648). In addition, we excluded participants with a diagnosis of Alzheimer’s disease, dementia (of any kind), or mild cognitive disorder (n = 1917). Finally, only participants with complete actigraphy and volumetric MRI data as well as data required for covariates (outlined in further detail below) were used in the analyses presented here (n = 10,083).
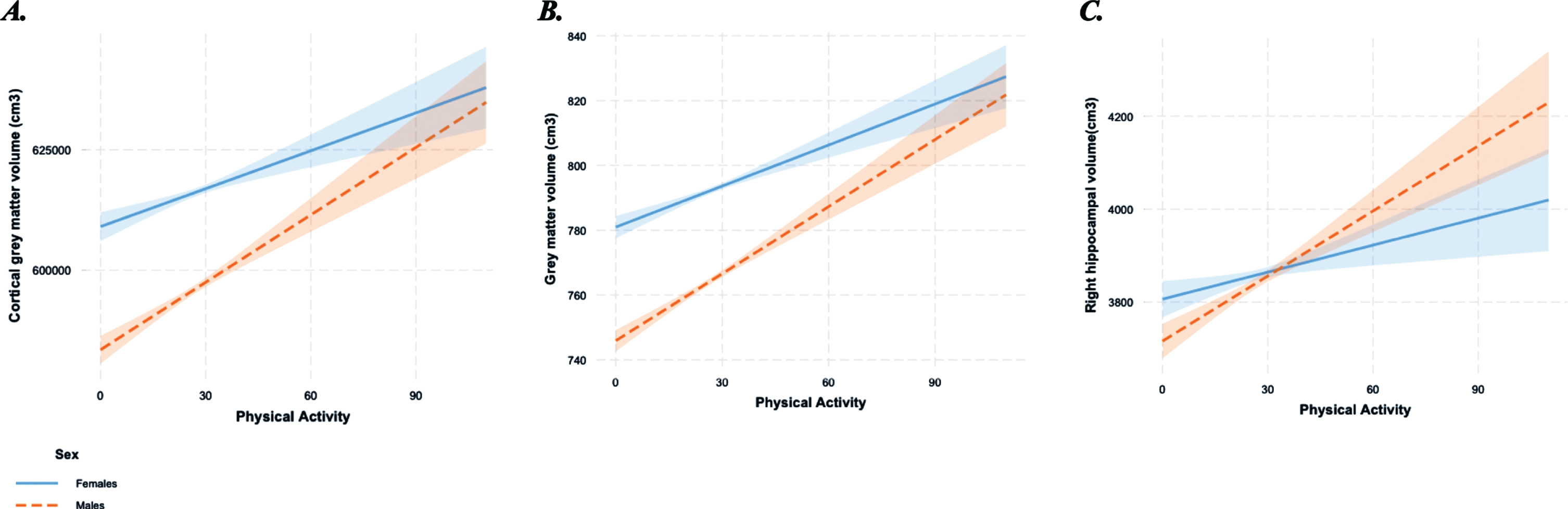
Interaction between physical activity (adjusted average daily accelerations) and sex on A) Cortical grey matter volume (normalized for head size scaling factor); B) Grey matter volume (normalized for head size scaling factor); C) Right hippocampal volume (normalized for head size scaling factor). Dotted line represents males and solid line represents females.
Habitual physical activity measurement (actigraphy)
Between February 2013 and December 2015, participants of the UK Biobank were invited at random to undergo objective habitual physical activity assessment through wearing an accelerometer for 7 days. From June 2013, participants that agreed to participate were sent Axivity AX3 wrist-worn triaxial accelerometers (Axivity Ltd., United Kingdom). The actigraphy units were programmed to commence data collection at 10 am two working days after postage and continue to collect acceleration data during wear-time on the dominant wrist over the following 7 days at 100 Hz with a dynamic range of ±8 g. Within the current study, we have used average daily acceleration, adjusted for wear time (in days), as our measure of habitual physical activity. This measure combines physical activity occurrence and intensity, and it is the recommended variable to use for physical activity analysis in the UK Biobank.
Structural magnetic resonance imaging
Volumetric MRI data were acquired using a Siemens Skyra 3T scanner (Siemens Healthcare, Erlangen, Germany), with a standard 32-channel RF receive head coil as per procedures outlined by the UK Biobank [28]. Briefly, the 3D MPRAGE T1-weighted images underwent acquired pre-processing and analysis using FSL packages (version 5, FMRIB Software Library, Oxford, England). Imaging-derived phenotypes, specifically total white and grey matter volumes, and grey matter regions of interest were generated by the image-processing pipeline developed by, and run on behalf of, the UK Biobank [29]. All brain volume variables used within the current study have been normalized for head size using the following equation: Normalized volume = raw volume –b *([head scaling factor] –[mean head scaling factor]); where ‘b’ is the slope of the regression of the region of interest on head scaling factor.
Genotyping
Genotyping for the UK Biobank was conducted by Affymetrix on purpose-designed arrays; ∼50,000 samples were analyzed using the BiLEVE Axiom array and ∼450,000 on the Affymetric UK Biobank Axiom array. Details of quality control practices have been described previously [30]. BDNFVal66Met polymorphism was used to classify participants as either Met carriers or Val/Val homozygotes. In addition, participants were identified as either APOE ɛ4 allele carriers or non-carriers.
Demographic and medical data
A number of variables were identified as potential covariates in the relationship between physical activity and brain volume, including education, depression, socioeconomic status, overall health rating, and vascular conditions and diabetes.
Participants reported their highest educational attainment (seven categories provided) and socioeconomic status was estimated using the Townsend deprivation index [31]. Participants were asked whether their doctor had diagnosed them with diabetes, hypertension, myocardial infarction, angina, stroke, or hypertension. Overall health was self-reported and scored on a 4-point scale from ‘poor’ to ‘excellent’. For any of the above variables, participants that stated ‘prefer not to answer’ or with missing data, were excluded from the analyses.
Statistical analysis
Statistical analyses were conducted in R (R Core Team; 2013) and Statistical Package for the Social Sciences (SPSS, IBM, Version 24). p-values from independent variables of interest and interactions were corrected for multiple comparisons using false discovery rate [32]. Descriptive statistics were calculated for demographic and outcome variables, and differences across sex, APOE ɛ4 allele, and BDNF Val66Met polymorphism carriage were examined using independent samples t-tests (continuous variables) and chi-square analyses (categorical variables).
Linear models were used to examine the relationship between habitual objectively-measured physical activity (average daily accelerations; independent variable) and brain volume (dependent variable) in all participants, adjusting for potential confounders: age, sex, body mass index, APOE ɛ4 allele, BDNF Val66Met, education, depression, Townsend deprivation index, overall health rating, and doctor diagnosed vascular conditions and diabetes (‘A’ models). The interaction terms physical activity*age, physical activity*sex, physical activity*APOE, physical activity*BDNF were then added to ‘A’ models to examine the moderating effect of non-modifiable factors on the relationship between physical activity and brain volume (‘B’ models).
RESULTS
Descriptive data
Examination of health and demographic differences between sexes (Table 1) revealed males were older (t = 11.0, p < 0.001), had a higher body mass index (t = 11.1, p < 0.001), lower levels of objectively-measured habitual physical activity (t = 6.6, p < 0.001), poorer self-reported health ratings (χ2 = 15.4, p < 0.01), higher prevalence of vascular conditions (χ2 = 143.5, p < 0.001) and diabetes (χ2 = 25.0, p < 0.001), compared with female participants. In addition, cross-sectional evaluations of normalized brain volume differences revealed males had smaller volumes of total grey matter (t = 34.5, p < 0.001), cortical grey matter (t = 28.6, p < 0.001), and right hippocampal volume (t = 2.1, p < 0.05; analysis not corrected for age or other covariates).
Descriptive statistics of UK Biobank participants at baseline (2006–2010) aged over 50 with accelerometer and MRI data, stratified by sex
*p < 0.05, **p < 0.01; ***p < 0.001; Chi-square analyses for categorical variables and independent sample t-tests were conducted for continuous variables. APOE, apolipoprotein E; BDNF Val66Met, brain-derived neurotrophic factor Valine66Methionine single nucleotide polymorphism.
Health and demographic details across APOE ɛ4 and BDNF Val66Met are detailed in Supplementary Table 1. The genotypic distribution of all polymorphisms of interest did not deviate from Hardy-Weinberg Equilibrium (rs6265, p = 0.20; rs429358, p = 0.43; rs7412, p = 0.95). Health and demographic differences between participants included in the current study, and those from the larger UK Biobank, are presented in Supplementary Table 2.
The relationship between habitual physical activity levels (average acceleration adjusted for time worn) and cortical grey matter volume (normalized for head size scaling factor) from magnetic resonance imaging
*p < 0.05; **p < 0.01; ***p < 0.001. 95% CI, 95% confidence interval; APOE ɛ4, apolipoprotein ɛ4 allele carriage; B, unstandardized beta value from linear model; BDNF Val66Met, brain-derived neurotrophic factor Valine66Methionine single nucleotide polymorphism; BMI, body mass index; LL, lower limit; UL, upper limit; PA, objectively-quantified physical activity (average accelerations adjusted for wear time).
Physical activity and brain volume
Physical activity was associated with larger total grey matter volume (B = 0.14, p = 0.001, q = 0.003), white matter volume (B = 0.21, p < 0.001, q < 0.001), and right hippocampal volume (B = 1.45, p = 0.008, q = 0.013; Tables 3–5).
The relationship between habitual physical activity levels (average acceleration adjusted for time worn) and white matter volume (normalized for head size scaling factor) from magnetic resonance imaging
*p < 0.05; **p < 0.01; ***p < 0.001. 95% CI, 95% confidence interval; APOE ɛ4, apolipoprotein ɛ4 allele carriage; B, unstandardized beta value from linear model; BDNF Val66Met, brain-derived neurotrophic factor Valine66Methionine single nucleotide polymorphism; BMI, body mass index; LL, lower limit; UL, upper limit; PA, objectively-quantified physical activity (average accelerations adjusted for wear time).
The relationship between habitual physical activity levels (average acceleration adjusted for time worn) and grey matter volume (normalized for head size scaling factor) from magnetic resonance imaging
*p < 0.05; **p < 0.01; ***p < 0.001. 95% CI, 95% confidence interval; APOE ɛ4, apolipoprotein ɛ4 allele carriage; B, unstandardized beta value from linear model; BDNF Val66Met, brain-derived neurotrophic factor Valine66Methionine single nucleotide polymorphism; BMI, body mass index; LL, lower limit; UL, upper limit; PA, objectively-quantified physical activity (adjusted average accelerations).
The relationship between habitual physical activity levels (average acceleration adjusted for time worn) and right hippocampal volume (normalized for head size scaling factor) from magnetic resonance imaging
*p < 0.05; **p < 0.01; ***p < 0.001. 95% CI, 95% confidence interval; APOE ɛ4, apolipoprotein ɛ4 allele carriage; B, unstandardized beta value from linear model; BDNF Val66Met, brain-derived neurotrophic factor Valine66Methionine single nucleotide polymorphism; BMI, body mass index; LL, lower limit; UL, upper limit; PA, objectively-quantified physical activity (adjusted average accelerations).
Interaction effects
An effect of the physical activity*sex interaction on cortical grey matter (B = 0.22, p = 0.003, q = 0.004), total grey matter (B = 0.30, p < 0.001, q = 0.001) and right hippocampal volume (B = 3.60, p = 0.001, q = 0.002; Tables 2, 5) was observed. Post-hoc analyses, following stratification of the cohort by sex, revealed that only males had an association between physical activity levels and greater cortical grey matter volume (B = 0.13, p = 0.01), total grey matter volume (B = 0.23, p < 0.001), and right hippocampal volume (B = 3.05, p < 0.001; Fig. 1A-C).
A physical activity*age interaction was observed on white matter volume (B = –0.03, p = 0.002, q = 0.01; Table 3). For clearer interpretation of the physical activity*age interaction, we stratified the cohort by median age (< 60 years, and ≥60 years) and re-ran the linear models. In those aged less than 60 years, there was an observed association between physical activity and greater white matter volume (B = 0.30, p < 0.001). This association did not exist in those aged 60 years and over (B = 0.14, p > 0.05).
DISCUSSION
The aim of the current study was to examine non-modifiable factors as moderators of the relationship between habitual objectively-measured physical activity and brain volume in a large population-based cohort of older adults (aged 50 years and over). In our total dataset of 10,083 older men and women, we found objectively-measured physical activity levels were associated with larger total grey matter volume, white matter, and right hippocampal volume. Through examination of potential moderating variables, we observed that sex moderated the relationship between physical activity and cortical grey matter, total grey matter, and right hippocampal volume; whereby only males had an association between objectively-measured physical activity and these brain volume measurements. In addition, we observed that the relationship between physical activity and white matter volume existed in the lower age range. We did not observe any moderating effects of APOE ɛ4 allele carriage or the BDNF Val66Met polymorphism on the relationship between physical activity and brain volume.
We report higher physical activity levels are linked with larger total grey matter, white matter, and right hippocampal volumes in our large sample. Our findings build on earlier work using UK Biobank data that report objectively measured physical activity to be associated with total grey matter volume and both left and right hippocampal volumes in 5,272 adults [33]. Our contrasting findings regarding left hippocampal volume could be attributed to differences in our samples, e.g., we implemented a minimum age cut-off of 50 years. Nevertheless, the literature regarding effects of physical activity and exercise on both right and left hippocampal volume is complex and inconsistent. Some studies report relationships between physical activity and both left and right hippocampal volumes [34], some studies report effects on only the left [12] and others only the right hippocampus [11]. The mechanisms underlying these differential effects remain to be elucidated; however, it is possible that physical activity-induced BDNF may act differently on the left and right hippocampi in different samples. Future research should also consider examining the functions of the left and right hippocampus (verbal and visual-spatial memory respectively), as this may provide insight into the functional outcomes of the physical activity-hippocampus relationship [35]. Indeed, previous work in this field suggests that in natural environments, physical activity has been associated with increases in cognitive demands on spatial orientation and memory, which may underpin the links between physical activity and brain health, and more specifically right hippocampal volume [36].
Our study revealed the relationship between physical activity and total grey matter and right hippocampal volumes was confined to males only. The current literature regarding sex differences in the physical activity-brain volume relationship is relatively small. Our findings are consistent with longitudinal work from Barha and colleagues [20] who identified higher levels of self-reported walking at baseline was associated with larger hippocampal volume 10 years later in males only, and larger dorsolateral prefrontal cortex in females only. It is possible that different brain regions are affected differently by physical activity in males versus females: this is reflected by functional data suggesting females are more likely to gain benefits from physical activity in terms of executive function (i.e., frontal lobe mediated brain function) [37]. In addition, BDNF expression is reduced in post-menopausal women, and although this effect can be partially attenuated with structured exercise, females may not achieve the same neurotrophic response as males [38]. Nevertheless, in a smaller cross-sectional analysis, higher levels of objectively-measured walking were associated with larger hippocampal volume in females only [21], with a follow-up study indicating this effect was confined to the subiculum [39]. It is important to note that only 26 males were included in this study, limiting the statistical power required to detect an effect. Another potential explanation for our findings could be due to the observed poorer health outcomes for men in our cohort. Indeed, males had a higher body mass index and reported poorer health ratings, both factors we know are associated with poorer brain health [40, 41]. It is possible that physical activity may play a role in mitigating declines in brain volume that are associated with poorer brain outcomes. In older adults an obesity paradox has been demonstrated [42]; therefore, it is also possible that greater body mass (as seen in males of this cohort) combined with physical activity may positively influence grey matter volume. Finally, the males in our cohort reported attaining higher levels of education, and thus in combination with greater physical activity, could lead to increased cognitive and brain reserve [43]. Although we detected sex as a moderator of the relationship between physical activity and brain volume in our large sample, this work requires further confirmation utilizing longitudinal analyses and exercise interventions to further understand the nature of these sex differences.
We identified an effect of age on the relationship between physical activity and white matter volume. Following stratification of the cohort by median age, it was identified that those aged less than 60 had an association between physical activity and white matter volume, yet those aged 60 years and over did not. Lifespan changes in white matter volume seem to follow a different trajectory compared with grey matter volume changes. Grey matter volume has been reported to peak in adolescence and then slowly decline with age, whereas white matter volume increases through early adulthood, peaks at around the age of 40 and declines more abruptly and faster than grey matter volume after 50 [44]. Therefore, one possible explanation to our results could be that physical activity might be more effective earlier in adulthood to mitigate or delay that quick decline. Later in life, once white matter reductions are more extensive, the ability of physical activity to induce white matter changes might wane. In this line, mice studies have shown that physical activity promotes the proliferation of oligodendrocyte precursor cells (OPCs) [45], fostering myelination and increasing hippocampal volume [46]. Unfortunately, OPCs reserves decline with age [47], which might limit the ability of physical activity to activate this mechanism in older adults.
Our study did not detect a moderating effect of the BDNF Val66Met polymorphism nor the APOE ɛ4 allele on the physical activity and brain volume relationship. This is in contrast to previous work by our group: we identified a link between physical activity and hippocampal volume in BDNF Val/Val homozygotes, but not Met carriers [26]. In addition, other studies have identified moderating effects of the BDNF Val66Met polymorphism in the relationship between physical activity and episodic memory (hippocampal-dependent function) [48, 49]. In regards to the potential moderating effects of the APOE ɛ4 allele, a previous cross-sectional evaluation of cardiorespiratory fitness and brain volume identified no effect of genotype [50]. Yet, over an 18 month follow up, Smith and colleagues identified higher rates of hippocampal atrophy in ɛ4 carriers that were low active, yet stable hippocampal volume was observed in individuals that were high active [4]. In the current analysis, our large sample ensured we were sufficiently powered to detect moderating effects of genotype; thus, our divergent findings could be attributed to demographic differences of the studied cohorts, or other methodological differences across the studies. Indeed, it is possible that certain populations (i.e., clinical populations) may experience greater declines in brain health associated with carriage of ‘detrimental’ genotypes, and that studying moderation of genes, in conjunction with certain characteristics could detect gene effects (i.e., undertaking a double moderation analysis). Furthermore, it is likely that a myriad of genetic factors influence the relationship between physical activity and the brain. Thus, investigations of combinations of genetic factors, such as in polygenic risk scores, as moderators of the physical activity-brain volume relationship warrant investigation, particularly in large samples.
Our study had some limitations. The causal direction of our findings must be interpreted with caution. More specifically, it is possible that in males only, brain volume may play a role in influencing participation in physical activity. Future longitudinal and interventional research may aid in elucidating the nature of the moderating effect of sex on the relationship between physical activity and brain volume. Our study investigated sex as a moderator of the relationship between physical activity and brain volume; however, our research is unable to identify gender differences (referring to social, environmental, and behavioral factors), which may also play a role in the differential effects of physical activity on brain health. Finally, accelerometer data and MRI brain imaging were not conducted at the same timepoint, and thus variation in the time difference between these measures may have influenced our findings. Nevertheless, for privacy issues, and to avoid self-identification, dates of these assessments were not available in order to calculate time between assessments.
Our findings using a large dataset provides further evidence that habitual physical activity levels are associated with larger brain volume. In addition, we found that only men received the benefit in terms of larger total grey matter and right hippocampal volumes. Future research is needed to both understand whether there is variability in response to exercise interventions between the sexes and the potential moderating effect of other genetic factors, in order to design individualized intervention programs for the purpose of reducing dementia risk.
Footnotes
ACKNOWLEDGMENTS
The data used to generate this work were obtained from the UK Biobank (application 45567). We are grateful to all UK Biobank study participants, who generously donated their time and the UK Biobank team for their work collecting and processing the data. The authors acknowledge Kelsey Sewell (Murdoch University) for assistance with figure preparation.