
Abstract
Background:
The olfactory system is affected early in Alzheimer’s disease and olfactory loss can already be observed in patients with mild cognitive impairment (MCI). Olfactory training is effective for improving olfactory and cognitive function by stimulating the olfactory pathway, but its effect on patients with MCI remains unclear.
Objective:
The aim of this randomized, prospective, controlled, blinded study was to assess whether a 4-month period of olfactory training (frequent short-term sniffing various odors) may have an effect on olfactory function, cognitive function, and morphology of medial temporal lobe (MTL) subregions and olfactory bulb in MCI patients.
Methods:
A total of thirty-seven MCI patients were randomly assigned to the training group or a placebo group, which were performed twice a day for 4 months. Olfactory assessments, cognitive tests and magnetic resonance imaging were performed at the baseline and follow-up period.
Results:
After the training, there was an increase in odor discrimination, and increased cortical thickness of bilateral hippocampus (CA23DG and CA1) and mean MTL. Additionally, the change of olfactory score was positively associated with change of volume of olfactory bulb and hippocampus; the change of global cognition was positively associated with change of cortical thickness of hippocampus, entorhinal cortex and mean MTL; the change of cortical thickness of entorhinal cortex was positively associated with change of executive function.
Conclusion:
Olfactory training was associated with an increase in cortical thickness of the hippocampus but not olfactory bulb volume in patients with MCI. Olfactory training may serve as an early intervention of preventing hippocampal atrophy.
INTRODUCTION
Impairment of olfaction is a characteristic and early symptom in individuals at risk for developing dementia [1], in particular those with mild cognitive impairment (MCI) [2–4]. Odor identification deficits may be a predictor of vulnerable MCI individuals who will convert to Alzheimer’s disease (AD) [2, 5], and the score of odor identification have been found to be associated with cortical thickness and volume in entorhinal, inferior temporal, and middle temporal cortices and the hippocampus [6]. Reduced olfactory function relates to hippocampal atrophy [7–9], an indicator of cognitive decline and established predictor for an imminent conversion of MCI to AD dementia [10].
argethispage2pt In light of the current literature, there is striking evidence that olfactory processing is closely linked to emotions and cognition in humans. This association has already been discussed in healthy individuals [7], patients with depression [11, 12], and patients with cognitive impairment [2], and consequently raises the question whether modifications in one of these functions might be accompanied by changes in other systems.
While research findings consistently demonstrate that a structured training with odors improves the olfactory ability of subjects with and without olfactory dysfunction [11, 13], data on possible therapeutic effects based on the interaction between olfaction, emotion, and cognition are still rare. We know from patients with major depression, that olfactory function shows improvement through the course of successful antidepressant therapy [14, 15]. Vice versa, in a study where Parkinson’s disease patients improved their sense of smell by structured short-term exposure to odorants (“olfactory training”) over 12 weeks, we observed a pronounced improvement of smell functions linked to higher cognitive functions which might point towards an observable cognitive effect [16]. Further, a recent study in healthy elderly provided evidence that continuous olfactory training has a positive effect on verbal fluency and subjective well-being [17]. In otorhinolaryngology, such a structured training with odors has been an established therapy in post-infectious and post-traumatic smell loss for many years now and is based on the fact that the olfactory system has the ability to change and recover [18]. Together with the improvement in smell function, an increase in olfactory bulb (OB) volume after the training could be observed [19], which illustrates the neuroplasticity of this structure and supports the finding of many studies that larger OB volume is associated with better olfactory function [20, 21].
In light of the above findings, the objective of the current prospective, controlled, blinded study was to evaluate the change in olfactory, cognitive, and depression scores in MCI patients over a 4-month period of olfactory training consisting of frequent short-term exposure to various odors. In addition to the detailed neuropsychological and olfactory examination, changes in medial temporal lobe (MTL) sub-regions and OB morphology, likely associated with early cognitive changes or loss of smell, have been investigated with magnetic resonance imaging (MRI). We hypothesized that a continuous olfactory training task would have a positive effect on olfactory and cognitive performance in MCI and would be associated with an increase in OB volume and MTL subregions cortical thickness.
MATERIALS AND METHODS
Patients
A total of 37 subjects were recruited at the neuropsychiatric Memory Clinic at Technische Universität Dresden, Germany. Eligible subjects were between 45–85 years of age had been diagnosed with MCI according to the most frequently used diagnostic criteria [22] and had no significant further health impairments (e.g., chronic hypothyroidism, renal insufficiency) that can be associated with disorders of the olfactory function. Further, any acute or chronic inflammation of the nose and sinuses, relevant previous or concomitant ENT diseases and a history of any other neurological or psychiatric illness had been defined as exclusion criteria.
Detailed information about the study was given to all participants and written consent was obtained prior to study inclusion. All aspects of the study were performed in accordance with the Declaration of Helsinki. The study protocol was approved by the local ethics board of the Technische Universität Dresden (EK 136032015).
Training with odorants
All patients performed olfactory training over a period of 4 months. At the baseline visit, participants randomly received glass jars either with high (verum) or subliminal (placebo) odor concentrations.
Patients had to expose themselves twice daily to four odors (phenyl ethyl alcohol [PEA]: rose, eucalyptol: eucalyptus, citronellal: lemon, and eugenol: cloves). These four odorants were chosen to be representative of four odor categories claimed by Henning in his work on the “odor prism” (Geruchsprisma), where he tried to identify primary odors. These categories are flowery: blumig (e.g., rose), foul: faulig, fruity: fruchtig (e.g., lemon), aromatic: würzig (e.g., cloves), burnt: brenzlig, and resinous: harzig (e.g., eucalyptus). Patients received four brown glass jars (total volume 50 mL) with one of the four odors in each (1 mL each, soaked in cotton pads to prevent spilling). All jars were labelled with the odor name. Patients were asked to sniff the odors in the morning and in the evening for approximately 20 s each. To focus their attention on the training, they were asked to keep a diary in which they rated their overall olfactory abilities each Sunday (data not analyzed). Further, patients received a phone call by one of the experimenters 8 weeks after the training started 1) to ask about the patients’ olfactory function and 2) to maintain compliance with the training procedure.
The placebo group did the same procedures as the verum group with odorless containers, but their odor containers and labels were the same as the ones used as for the verum group. Therefore, the treatment expectancies of the two groups were controlled, and the only difference between the two groups was the presence of high odor concentrations in cotton pads in the containers.
Olfactory testing
Olfactory testing was performed before and after the training period of 4 months using the “Sniffin’ Sticks” test kit (Burghart Messtechnik, Wedel, Germany) which involves tests for odor threshold, odor discrimination, and odor identification. “Sniffin’ Sticks” are a validated psychophysical test of olfaction and have been described extensively in the literature [23]. Olfactory function was assessed at the level of odor thresholds for phenylethylalcohol (PEA), a relatively selective stimulant of the human olfactory system, and at a supra-threshold level for odor discrimination and odor identification. The sum of the scores from the three subtests resulted in the TDI score (Threshold, Discrimination, Identification) with a maximum of 48 points. A TDI score of 30.5 points or more suggests normosmia, a score between 16.5 and 30 points indicates reduced olfactory function in terms of hyposmia, and a score of less than 16.5 points implies functional anosmia.
Neuropsychological assessment
Cognitive assessments before and after training included dementia screening (Mini-Mental State Examination, MMSE) [24], verbal and visual learning and memory skills (word list learning, word list delayed recall, and visual memory delayed recall items of the Consortium to Establish a Register for Alzheimer’s Disease-Neuropsychological (CERAD-NP) test battery [25]), verbal fluency (animals, S-words), the digit span forward test from the Wechsler Memory Scale-Revised (WMS-R) [26], and the quantitative test of Trail Making Test (TMT) part A and B [27], the Boston Naming Test (BNT) [28], and the Nuremberg age inventory (NAI) Labyrinth-Test, and colored words test (“Farb-Wort-Test”, FWT) [29]. The Beck Depression Inventory-II (BDI-II) was administered to assess self-reported symptoms of depression before and after training.
Neuroimaging
A 3-Tesla MRI scanner (Siemens Sonata, Erlangen, Germany) with a 12-channel head coil was used for image acquisition. The protocol included a whole brain anatomical sequence without interslice gap (5-mm-thick standard T1-weighted 3D sequence) for every participant to rule out any organic brain disorders. Further, oblique coronal T2-weighted fast-spin echo scans (repetition time: 5,200 ms; echo time: 105 ms; slice thickness: 3 mm; spacing: 0 mm; 19 slices; inplane voxel size: 0.39×0.39 mm; field of view: 200 mm) were obtained.
Olfactory bulb volume
T2-weighted fast spin-echo images covering the anterior and middle segments of the base of the skull were offline processed and left and right OBs limits were drawn manually on each coronal slice using the AMIRA 3D visualization and modeling system (Visage Imaging, Carlsbad, USA). OB volumes were calculated by planimetric manual contouring (surface in mm2) and all surfaces were added and multiplied by 3 (3-mm slice thickness) to obtain a volume in cubic millimeters. The field of view was 256×256 mm2. The sudden change of diameter at the beginning of the olfactory tract was used as the distal demarcation of the OB, as suggested by Yousem and colleagues [30, 31].
Cortical thickness
To investigate cortical thickness within the MTL, the MRI data was analyzed with a cortical unfolding technique [32–34]. The method allows cortical thickness measurements within subregions of the hippocampus and its adjacent structures, thus, the cornu ammonis fields 1–3 including the dentate gyrus, the subiculum, entorhinal cortex, perirhinal cortex, parahippocampal cortex, and the fusiform gyrus. A region of interest (ROI) analysis of cortical thickness for the following subregions was conducted: CA1 (cornu ammonis), CA23DG (dentate gyrus), SUB (subiculum), ERC (entorhinal cortex), PRC (perirhinal cortex), PHC (parahippocampal cortex), and FUS (fusiform gyrus). Hippocampal thickness was defined as an average measure of CA23DG, CA1, and SUB (Fig. 1). Mean MTL thickness was defined as an average measure of all subregions. These ROIs are flattened into two-dimensional space based on metric multidimensional scaling after the manual masking of cerebrospinal fluid (CSF) and white matter. Before their mathematical projection to the flattened cortical maps, regional boundaries are specified using histological and MRI atlases [35, 36]. Mean thickness in each subregion was computed by averaging the thickness across all 2-D voxels within each ROI. Further, total hippocampal thickness and total MTL thickness in each subject was calculated by performing a similarly weighted average across all the subregions.
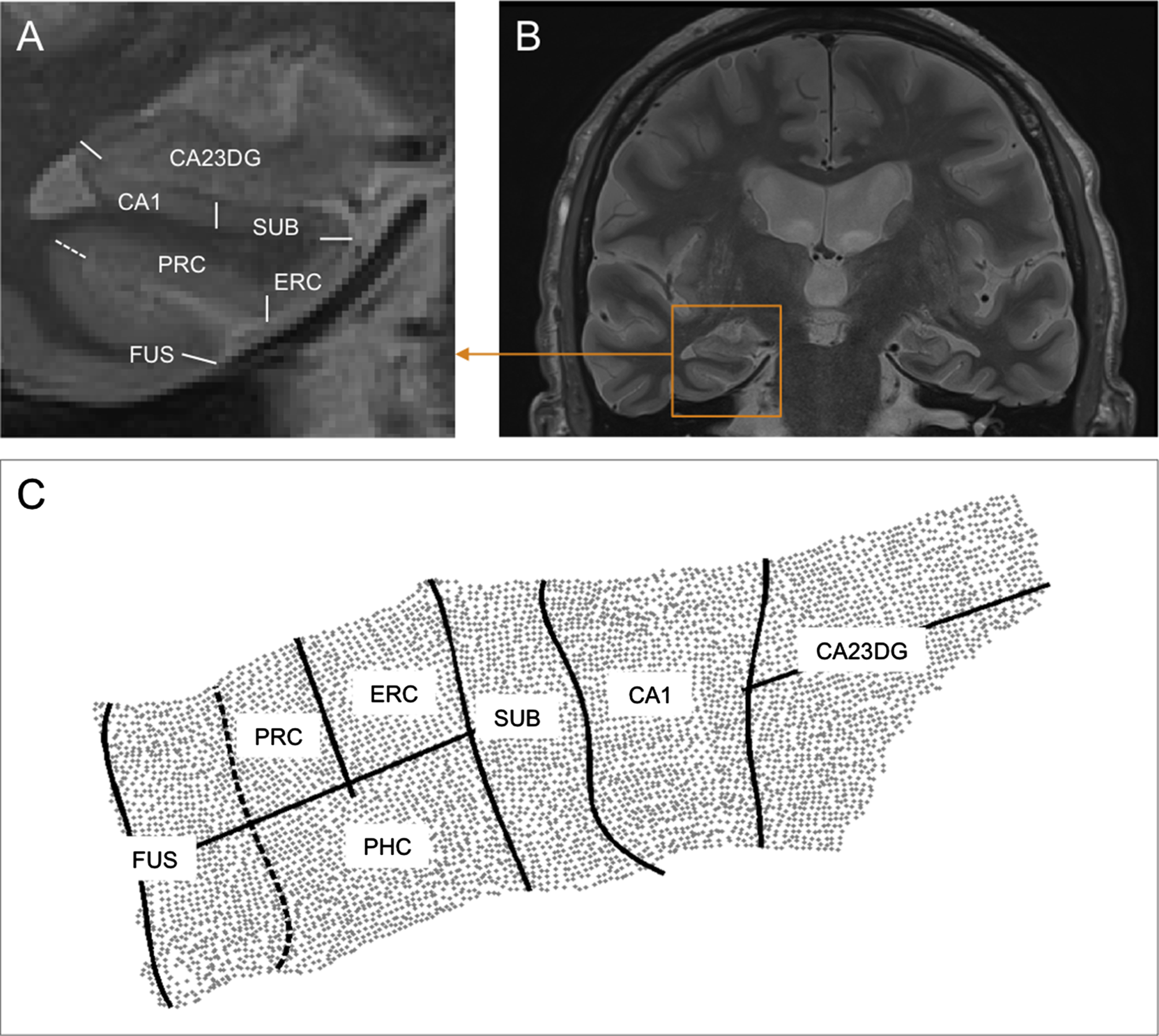
Cortical unfolding of MTL subregions. After oblique coronal scanning perpendicular to the long axis of the hippocampus (B), hippocampal and adjacent medial temporal lobe region boundaries are delineated (A). After segmenting white matter, gray matter. and CSF, the entire gray matter volume is mathematically flattened into 2-dimensional space (C). CA23DG, cornu ammonis fields 2,3, and dentate gyrus; CA1, cornu ammonis field 1; SUB, subiculum; ERC, entorhinal cortex; PRC, perirhinal cortex; PHC, parahippocampal cortex (not visible in A); FUS, fusiform gyrus. Dotted line represents the collateral sulcus.
Statistical analyses
The statistical analysis was performed with SPSS (version 25; IBM Corp., Armonk, NY, USA). If not mentioned otherwise, all data are displayed as means±standard deviation (SD) or numbers. Two-sample t-tests were used to compare the baseline characteristics between verum group and placebo group. The effect of training on changes of olfactory scores, cognitive scores and neuroimaging indicators were analyzed by performing repeated-measures ANCOVA, with “group” (verum group versus placebo group) and “time” (baseline versus follow-up) as factors, and control variables “age” and “sex”. Paired-t tests were used to explore the change of olfactory scores, cognitive scores, and neuroimaging indicators in verum group and placebo group, respectively. Partial correlation analysis was used to explore the relationship between change of olfactory scores, cognitive scores and neuroimaging indicators, and control variables included age and sex.
RESULTS
Study population
Thirty-seven patients (17 men, 20 women; mean±SD age: 70.9±6.2 years [range: 56–82 years]) participated in the study; 19 were randomly assigned to the verum group (“high-training group”) and 18 to the placebo group (“low-training group”). Baseline characteristics are shown in Table 1. In terms of these characteristics there were no significant differences between groups (p > 0.05).
Baseline descriptive statistics of patient groups who trained with high (verum) or subliminal (placebo) odor concentrations
TDI, olfactory sum score (threshold, discrimination, identification); MMSE, Mini-Mental State Examination; CERAD-NP, Consortium to Establish a Register for Alzheimer’s Disease-Neuropsychological; WMSR, Wechsler memory test range of numbers; NAI, Nuremberg age inventory; TMT, Trail-Making-Test.
Olfactory function
In the verum group, but not in the placebo group, there was an effect of “time” on odor discrimination score (F = 6.24, p = 0.018) suggesting improvement after training. There was no significant effect of “time*group” for odor threshold, odor discrimination, odor identification and TDI scores (p > 0.05) (Table 2) suggesting that both groups changed in a similar way.
Change of olfactory and cognitive scores in the verum group (high odor concentrations) and the placebo group (subliminal odor concentrations)
TDI, olfactory sum score (threshold, discrimination, identification); MMSE, Mini-mental State exam; CERAD-NP, Consortium to Establish a Register for Alzheimer’s Disease-Neuropsychological; WMSR, Wechsler memory test range of numbers; NAI, Nuremberg age inventory; TMT, Trail-Making-Test. The “time*group” represents the interactive effect of time and group on various scores.
Cognitive tests
There was a significant effect of “time*group” on WMS-R digit span backward (F = 5.66, p = 0.024), but there was no significant difference in the post-hoc comparison between groups. There was no effect of “time” on any cognitive score (Table 2).
Olfactory bulb volume and hippocampal volume
There was no significant effect of “time*group” or “time” on left, right and mean OB volume, as well as on left and right hippocampal volume (p > 0.05) (Table 3).
Change of olfactory bulb volume, hippocampal volume (in mm3) and cortical thickness (in mm3) of MTL subregions in in the verum group (high odor concentrations) and the placebo group (subliminal odor concentrations)
OB, olfactory bulb; DG, dentate gyrus; SUB, subiculum; ERC, entorhinal cortex; PRC, perirhinal cortex; PHC, parahippocampal cortex; FUS, fusiform gyrus; MTL, medial temporal lobe. The “time*group” represent the interactive effect of time and group on various indicators.
Cortical thickness of MTL subregions
There is a significant effect of “time” on mean MTL thickness, left MTL thickness, right MTL thickness, and mean hippocampal thickness (p < 0.05), with the bilateral subregions CA23DG and CA1 contributing to this finding (Table 3). Specifically, in the verum group, mean hippocampal thickness increased in the left hemisphere (t = 2.96, p = 0.01; contributing subregions CA23DG: t = 3.78, p = 0.02, CA1: t = 3.36, p = 0.04), and in the right hemisphere (t = 4.32, p = 0.001; contributing subregions CA23DG: t = 2.53, p = 0.023, CA1: t = 6.49, p < 0.001). In the placebo group, mean hippocampal thickness increased in the left hemisphere (t = 3.01, p = 0.01; contributing subregions CA23DG: t = 3.20, p = 0.08, CA1: t = 3.36, p = 0.04), and in the right hemisphere (t = 3.43, p = 0.005; contributing subregion CA1: t = 3.21, p = 0.008). Additionally, there was a significant effect of “time*group” on thickness of left PRC (F = 5.86, p = 0.022) and left PHC (F = 7.23, p = 0.012) (Table 3), but there was no significant change between baseline and follow-up in both group in the post-hoc comparison (p > 0.05).
Correlations
The partial correlation analyses suggested that the change of odor identification was associated with change of mean OB volume (r = 0.45, p = 0.035) and change of mean hippocampal volume (r = 0.66, p = 0.001); the change of TDI was associated with change of mean hippocampal volume (r = 0.49, p = 0.021); the change of MMST was associated with change of bilateral mean MTL thickness (r = 0.41, p = 0.046), mean hippocampal thickness (r = 0.57, p = 0.006), with cortical thickness changes in the subregions CA1 (r = 0.66, p = 0.001), and ERC (r = 0.48, p = 0.024); the change of TMT B was associated with change of mean ERC (r = –0.48, p = 0.025) (Fig. 2). No significant association was found between other changes of variables (p > 0.05).
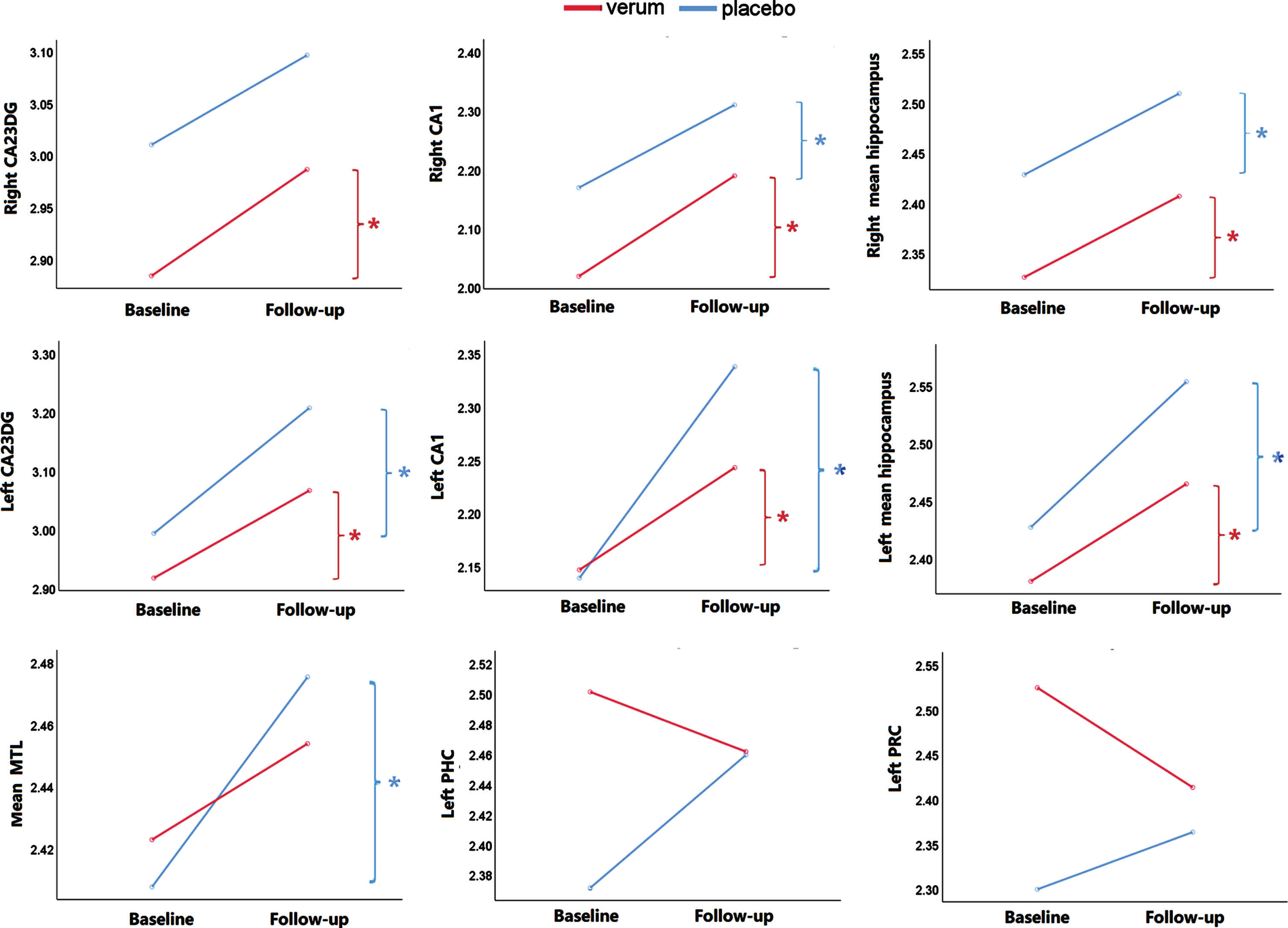
Significant change of cortical thickness in the verum group (high odor concentrations) and the placebo group (subliminal odor concentration). In the verum group, the cortical thickness of left CA23DG, left CA1, left mean hippocampal, right CA23DG, right CA1 (t = 6.49, p < 0.001), right hippocampal improved significantly. In the placebo group, the thickness of left CA23DG, left CA1, left mean hippocampal, right CA1, right mean hippocampal and mean MTL improved significantly. There was a significant effect of “time*group” on thickness of left PRC and left PHC. DG, dentate gyrus; PRC, perirhinal cortex; PHC, parahippocampal cortex; HC, hippocampus; MTL, medial temporal lobe. *p < 0.05.
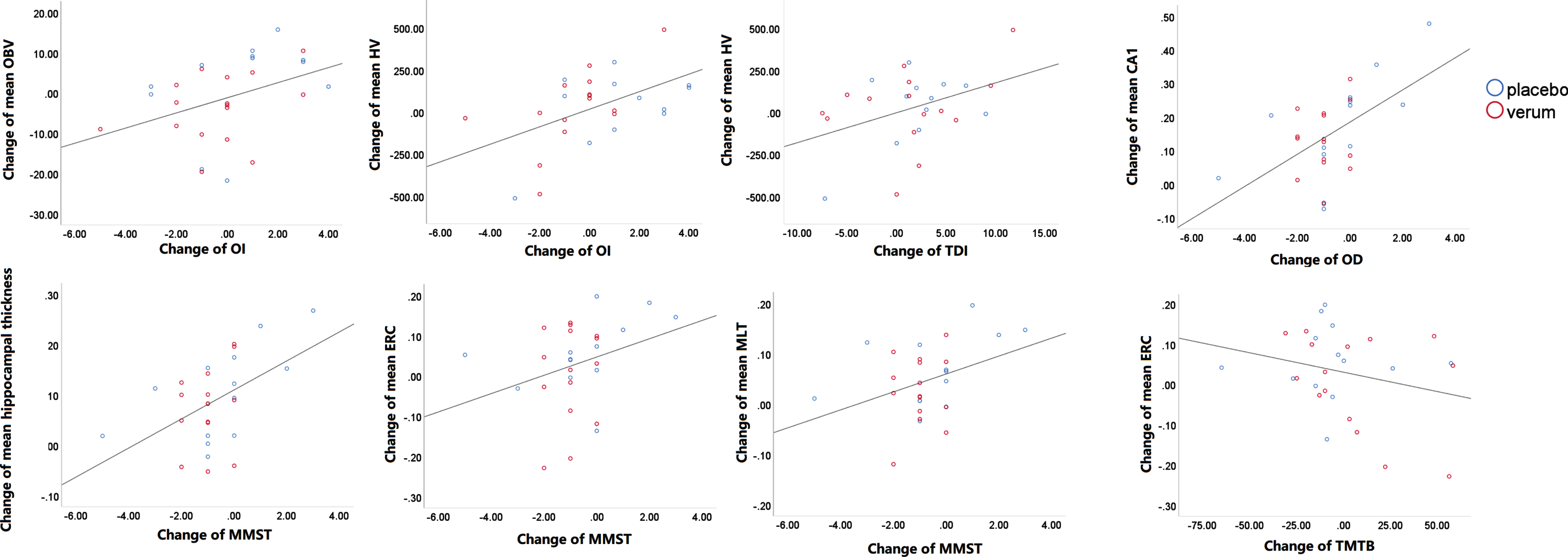
Associations between change of olfaction, cognition, and olfactory structure. Controlling for the age and sex, the partial correlation analyze suggested that the change of odor identification was associated with change of mean OB volume (r = 0.452, p = 0.035) and change of mean hippocampal volume (r = 0.661, p = 0.001); the change of TDI was associated with change of mean hippocampal volume (r = 0.489, p = 0.021); the change of MMSE was associated with change of mean CA1 (r = 0.655, p = 0.001), change of mean hippocampal thickness (r = 0.567, p = 0.006), change of mean ERC thickness (r = 0.478, p = 0.024) and change of mean MTL (r = 0.411, p = 0.046); the change of TMT B was associated with change of mean ERC thickness (r = –0.477, p = 0.025). OBV, olfactory bulb volume; ERC, entorhinal cortex; HV, hippocampal volume; MTL, medial temporal lobe; OI, Odor identification; OD, Odor discrimination; MMSE, Mini-Mental State Examination; TMT, Trail-Making-Test.
DISCUSSION
The present study explored the effect of training with odors on olfactory and cognitive function and on structures associated with smell and cognition in patients with MCI. The following results emerged: First, cortical thickness of the hippocampal subregions CA23DG and CA1 increased significantly in the verum group (high odor concentrations) and even in the placebo group (subliminal odor concentrations), but their OB volume and hippocampal volume remained stable. Second, there was a general training effect on increased odor discrimination in patients with MCI; significant improvement was reached in the verum group only. Third, a change of olfactory score was associated with a change of mean OB volume and mean hippocampal volume; the change of MMSE was associated with changes of mean MTL thickness and mean hippocampal thickness, with CA1 and ERC contributing to this finding.
The hippocampus plays a critical role in spatial navigation and formation of declarative memory, and it is one of the most important regions of the secondary olfactory cortex [37]. In the entorhinal-hippocampal circuit, the hippocampal CA1 receives information from entorhinal cortex via direct paths (ERC layer 3⟶CA1) and indirect path (ERC layer 2⟶DG⟶CA23⟶CA1), which is essential for memory formation and retrieval [38]. Among these subregions, the CA1 field is especially vulnerable to amyloid and tau pathology, and CA1 pyramidal cells are especially susceptible to neuronal loss and dendritic alterations in AD patients [39]. Moreover, hippocampal atrophy is a typical characteristic of AD [40], and stimulating hippocampus may contribute to the prevention of developing AD [41]. Animal studies suggested that training with odor caused up-regulation of microRNAs [42], mitochondrial small RNAs [43], endogenous siRNAs and noncoding RNA-derived small RNAs [44], but its effect on human hippocampus has not been reported yet. The present results suggested that not only training with odors, but also pure sniffing training can significantly increase cortical thickness of hippocampal subregions (especially CA1 and CA23DG), and the increased cortical thickness of hippocampus was associated with global cognitive function and odor discrimination. For one thing, the repeated odor-exposure induced synapse formation and spine sprouting, which increases cell genesis of glial or neuronal cells, and finally results in an increase of gray matter density or thickness [45, 46]. For another, within the dentate gyrus of the hippocampus as well as olfactory bulb we have the special situation that morphological changes due to neuronal plasticity is not only restricted to synapses and neurites but also to entire neurons through adult neurogenesis. Adult neurogenesis is activity-depended regulated and supports processing and storage of new memories. In rodents, odor enrichment increases the number of new born neurons in the olfactory bulb [47] and hippocampus [47]. Although adult neurogenesis decreases with age it is a lifelong and functional relevant process in the adult human brain, which is disrupted in patients with AD and MCI [49, 50]. In contrast to rodents, human adult neurogenesis is much more prominent in the hippocampus and negligible low in the olfactory bulb [51]. This might explain why olfactory training may have a selective effect on hippocampal thickness, which is likely linked to stimulation of adult hippocampal neurogenesis. It should also be considered that physical exercise has been suggested to increase the size of the hippocampus by regulating neurogenesis [52] and synaptic integrity [53], which might help to explain the effect of sniff training on the hippocampus. Furthermore, the repeated sniffing during the 4 months of training may also increase awareness of various odors the in daily life, and lead to similar effect of training with odors. Interestingly, recent studies suggested that nasal respiration is associated with memory processes, indicating that the effect of nasal respiration on neuronal activity is not only restricted to the olfactory system but also impacts other regions, and the activities of hippocampus and prefrontal cortex provide a potential neuronal mechanism behind respiratory modulation of memory [54].
Although the change of hippocampal volume is also positively associated with change of olfactory function, there was no significant effect of training on hippocampal volume, probably because the surface-based analysis is more sensitive to reflect the subtle change of grey matter compared with the voxel-based analysis. Consistently, Aïn et al. also found that olfactory training exhibited no effect on the voxel-based analysis, but significant change was observed in their analyses of cortical thickness in healthy young individuals [55]. Overall, sniff training is a promising early invention for AD patients, as well as other diseases related to hippocampal atrophy. Further studies with longer follow-up and larger sample sizes are needed to confirm the effect of sniff training and its underlying mechanism need to be clarified.
The present study suggested that the change of cortical thickness of the entorhinal cortex was positively associated with changes of global cognition and executive function, which is consistent with previous studies suggesting that the entorhinal cortex plays essential role in cognitive processing [56]. However, both verum group and placebo group did not exhibit significant changes of the cortical thickness of the entorhinal cortex, possibly because the tau pathology starts from the entorhinal cortex and propagates to the hippocampus in the course of AD disease [57]. A more severe damage of the entorhinal cortex renders it less responsive to the effects of training.
Improvement of OB volume by olfactory training has been reported in patients with idiopathic olfactory loss and healthy participants [58]. However, the present study suggested that both the verum group and placebo group exhibited stable OB volume after training, which is not consistent with our hypothesis. One explanation might be that the OB volume is reduced and exacerbate with disease development in subjects with MCI [59]. The training procedures, however, exhibited a maintaining effect on the OB volume, and their effect was more obvious on the high-order rather than the peripheral olfactory system. Furthermore, as mentioned above in contrast to the hippocampus there is very limited adult neurogenesis in the OB of aging humans [51]. Another explanation might be the analysis technique. Because the OB is very small and is not included in the template of voxel-based and surface-based analyses, the calculation of OB volume is performed by manually drawing, which is different from the analyses of MTL structure and may contribute to the differing results from the two analyses. Additionally, the change of the OB volume was associated with change of odor identification, which is consistent with previous studies that OB is also involved in high-order olfactory processing.
The present study suggested that training with odors significantly improved odor discrimination in MCI patients, which is consistent with previous studies in patients with olfactory loss. The increased odor discrimination might be considered indirect evidence of the improvement of cognitive function because cognitive processing is strongly involved in odor discrimination [60]. However, there was no other significant effect of training on olfactory and cognitive scores. Considering the positive association between changes of olfactory scores, cognitive scores, and hippocampal cortical thickness, it is possible that the improvement of hippocampal cortical thickness did not reach the significant level of improving the cognitive score. Based on the finding that morphological changes are restricted to the hippocampal formation, we suggest that adult neurogenesis is a key process involved in olfactory training-induced cortical thickness increase. This is in line with rodent and human studies showing that odor enrichment increases adult neurogenesis in OB and hippocampus, whereas neurogenesis in aging humans is mainly restricted to the hippocampus. Additionally, several limitations of the present study may also contribute to the negative effect. First, because MCI patients are characterized by amnestic deficits, some of the patients may have forgotten the daily training and did not receive the complete effect of training. Second, the follow-up point is 4 months after training in the present study, and dynamic changing of cognitive function may not be captured because the training may exhibit a delayed effect (longer than 4 months) or a temporary effect (improvement before 4 months, and consecutive decrease). Third, because the assessment of cognitive function was not as detailed as the assessment of brain structures, the relatively small sample size might not have been sufficient for demonstrating the effect of training on cognitive change.
There are limitations in the present study. First, because a control group without training was not included, there was no direct evidence excluding the time effect on cortical thickness of hippocampus. However, it is well acknowledged that the hippocampal atrophy becomes pronounced in patients with MCI, and hippocampal atrophy is impossible to reverse spontaneously. Therefore, we believe that the increased cortical thickness of hippocampus resulted from the training in the present study. Second, the drop-out and relatively low compliance of MCI patients probably limited the effect of training. Future studies could better clarify the training effect by enhancing the supervision and cooperation. Third, the present sample size and follow-up period may not have been sufficient for demonstrating the effect of training. Fourth, the present study did not include CSF or PET assessments and was not able to define “MCI due to AD", and the training effect may be limited by heterogeneity of MCI. Finally, the present study included a verum group and a placebo group only, and future studies including a control group without any training could better clarify the effect of olfactory training in MCI subjects.
Conclusion
Training with odors was associated with an increase of the cortical thickness of hippocampus but not olfactory bulb volumes in patients with MCI. Possibly, this may serve as an early intervention of preventing hippocampal atrophy. Future studies are needed to confirm the effect by applying a longer follow-up, enlarging the sample size, and enhancing the supervision. It is necessary to elucidate the underlying mechanism of how sniff training modifies the hippocampus.