
Abstract
Background:
Chronic psychological stress (PS) hinders the treatment of diabetes-associated cognitive decline (DACD). However, the impact of chronic PS on the risk of developing DACD remains unclear. There is growing evidence that gut flora interventions are promising targets for treating stress-related diseases.
Objective:
We examined whether chronic PS triggers or exacerbates the onset of DACD in rats and aimed to elucidate whether ZiBuPiYin recipe (ZBPYR) prevents and treats chronic PS-aggravated DACD by dynamically maintaining the components of the gut microbiota.
Methods:
We performed chronic PS (restraint, rotation, and congestion) on ZDF rats to establish a model. Cognitive function was evaluated by behavioral experiments, and activation of the hypothalamic-pituitary-adrenal axis was detected by ELISA. Weekly feces from rats were collected for 16 S RNA sequencing.
Results:
We found that chronic PS promoted cognitive abnormalities and exacerbated DACD phenotypes. Additionally, chronic PS altered intestinal flora diversity, dynamically elevating the abundance of Alistipes and Coprococcus; enriching Module 1 (Dorea, Blautia, Ruminococcus) and Module 48 (Blautia); and inhibiting Module 20 (Lactobacillus, SMB53), and Module 42 (Akkermansia). ZBPYR significantly alleviated hyperglycemia and cognitive impairment in chronic PS-aggravated DACD rats and dynamically reduced the abundance of Alistipes and Coprococcus; significantly enriched Module 3 (Ruminococcus) and Module 45 (Lactobacillus, Coprococcus, SMB53); and suppressed Module 2 (Lactobacillus), Module 16 (Turicibacter, Trichococcus, Lactobacillus, 02d06, Clostridium), Module 23 (Bifidobacterium), and Module 43 (Clostridium).
Conclusion:
ZBPYR might prevent and treat chronic PS-aggravated DACD by dynamically regulating Lactobacillus, Alistipes, and Coprococcus.
Keywords
INTRODUCTION
Chronic psychological stress (PS) is a significant risk factor for the development of type 2 diabetes mellitus (T2DM). Activation of the hypothalamic-pituitary-adrenal axis (HPA) is considered to be the main stress pathway involved in this association [1]. Diabetes-associated cognitive decline (DACD) is a typical neurological complication of T2DM and mainly manifests as cognitive deficits, inattention, and learning and memory impairment. A dysregulation of the HPA axis, brain insulin resistance, and an oxidative stress are the major factors that contribute to its pathogenesis [2, 3]. However, whether chronic PS is a risk factor for DACD and the mechanism deserve further investigation.
According to emerging evidence, the gut microbiota play a crucial role in DACD [4]. The gut microbiota in patients with cognitive impairment exhibits decreased diversity, with increased abundance of Firmicutes and decreased abundance of Bacteroidetes [5]. Transplantation of fecal microbes from DACD mice to germ-free mice induces cognitive impairment, whereas fecal microbiota transplantation from mice without DACD reverses the detrimental effects on cognitive function [6]. An alteration in beta-diversity and host gut bacteria was associated with this effect [6]. Furthermore, it has been proposed that the gut and brain communicate bidirectionally via the microbiota-gut-brain axis [7, 8]. Recently, increasing evidence suggests that there is also a bidirectional interaction between PS and the microbiome, and altered gut flora was found during PS [9]. The composition and diversity of bacterial communities have been shown to be altered by chronic water avoidance stress and repetitive restraint stress [10]. Maternal separation stress in rats leads to altered microbiota composition in pups [11]. The relationship between PS and gut microbiota remains to be further studied. By acting on gut microbes, probiotics can relieve somatic and behavioral stress responses [12]. High dietary fiber helps alleviate diabetes and cognitive dysfunction by regulating gut microbiota [13, 14]. Thence, targeting the gut microbiota using probiotics, drugs, and dietary improvements will be a new type of treatment that could improve DACD aggravated by chronic PS.
In over 2,500 years, traditional Chinese medicine (TCM) has been developed and advocated for treating many diseases in China. Multiple studies show that the gut microbiota may be involved in the effects of TCM formulations including GQD [15] and AMC [16]. This suggests that TCM is capable of modulating the gut microbiota as an underlying mechanism to treat disease. A formula consisting of twelve herbs (ZiBuPiYin recipe, ZBPYR) is originated from the ancient formula Zicheng Tang used for the treatment of cognitive disorder [17]. Studies have demonstrated that the herbs used in this formula, such as Panax ginseng C. A. Mey. [18, 19], Dioscorea polystachya Turcz. [20–22], Poria cocos (Schw.) Wolf [23], Paeonia lactiflora Pall. [24, 25], Salvia miltiorrhiza Bunge [26], Polygala tenuifolia Willd. [27, 28], and Santalum album L. [29], can significantly improve blood glucose levels and behavioral and cognitive functions. Furthermore, it has also been used clinically for more than 10 years and has shown good efficacy. ZBPYR contains the herbs Panax ginseng C. A. Mey. [30, 31] and Salvia miltiorrhiza Bunge [32], which have been clinically demonstrated to treat disease, possibly via regulation of the intestinal flora.
The gut microbiome is an important system in the human body and has a significant impact on behavior. Research have suggested that gut microbiota may cause an imbalance in the HPA axis and thus cause disruptions in the brain neuroendocrine system, which can affect the body’s sensitivity to stress. Modulating the gut microbiota may offer a new way to treat stress-related disorders [33]. ZBPYR may prevent and treat DACD aggravated by chronic PS through the dynamic maintenance of intestinal flora. To verify the hypothesis that alterations of the intestinal microbiota by ZBPYR can prevent and treat DACD symptoms aggravated by chronic PS, male obese Zuker diabetic fatty (ZDF) rats were allocated to the diabetes group, the chronic PS-aggravated DACD group, and the ZBPYR intervention group, while age-matched Lean Zucker (LZ) rats were the control group. The dynamic effects of ZBPYR on the gut microbiota were assessed by 16 S rRNA sequencing.
MATERIALS AND METHODS
Animals
LZ (fa/+) rats and ZDF (fa/fa) rats, 6 weeks of age, were acquired at Vital River Laboratories (Beijing, China). Experiments were carried out in conformity with the National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by the Animal Ethics Committee of Nanjing University of Chinese Medicine (ACU170606). The animals were acclimatized for 7 days in a 12-h light and dark experiment center with free access to water and food before experiments.
ZBPYR
ZBPYR is composed of 12 Chinese Herbal Medicines, as shown in Table 1. All herbs were purchased from the Sanyue Chinese Traditional Medicine Co., Ltd. (Nantong, China). Preparation and administration of ZBPYR were consistent with those described in preceding studies [17, 34]. We previously studied the chemical composition, quality control criteria, and pharmacokinetics of ZBPYR. We initially identified 155 chemical components in ZBPYR [35].
The ingredients of ZBPYR
Experimental design
According to body weight (BW) and random blood glucose (RBG), animals were assigned to four groups (n = 4 per group): LZ group (normal control), ZDF group (ZDF control), ZDF_P group (chronic PS-aggravated DACD), and P_Z group (chronic PS-aggravated DACD combined with ZBPYR administration). ZDF_P and P_Z groups were given restriction, rotation, and congestion stimulation for six weeks as previously described [34]. BW and levels of RBG were measured from weeks 1 to 8 of the experiment. At the end of 6 weeks of chronic PS, the oral glucose tolerance test (OGTT) and insulin tolerance test (ITT) were carried out. Behavioral experiments were performed on the last 7 days of the experiment, and the steps were as follows: the open field test (OFT) on day 1, MWM test on days 2–7, and the experimental methods were referred to the previous study [34]. After the experiment, the levels of corticosterone were measured. The schematic diagram of the experimental groups and experimental operations is shown in Fig. 1.
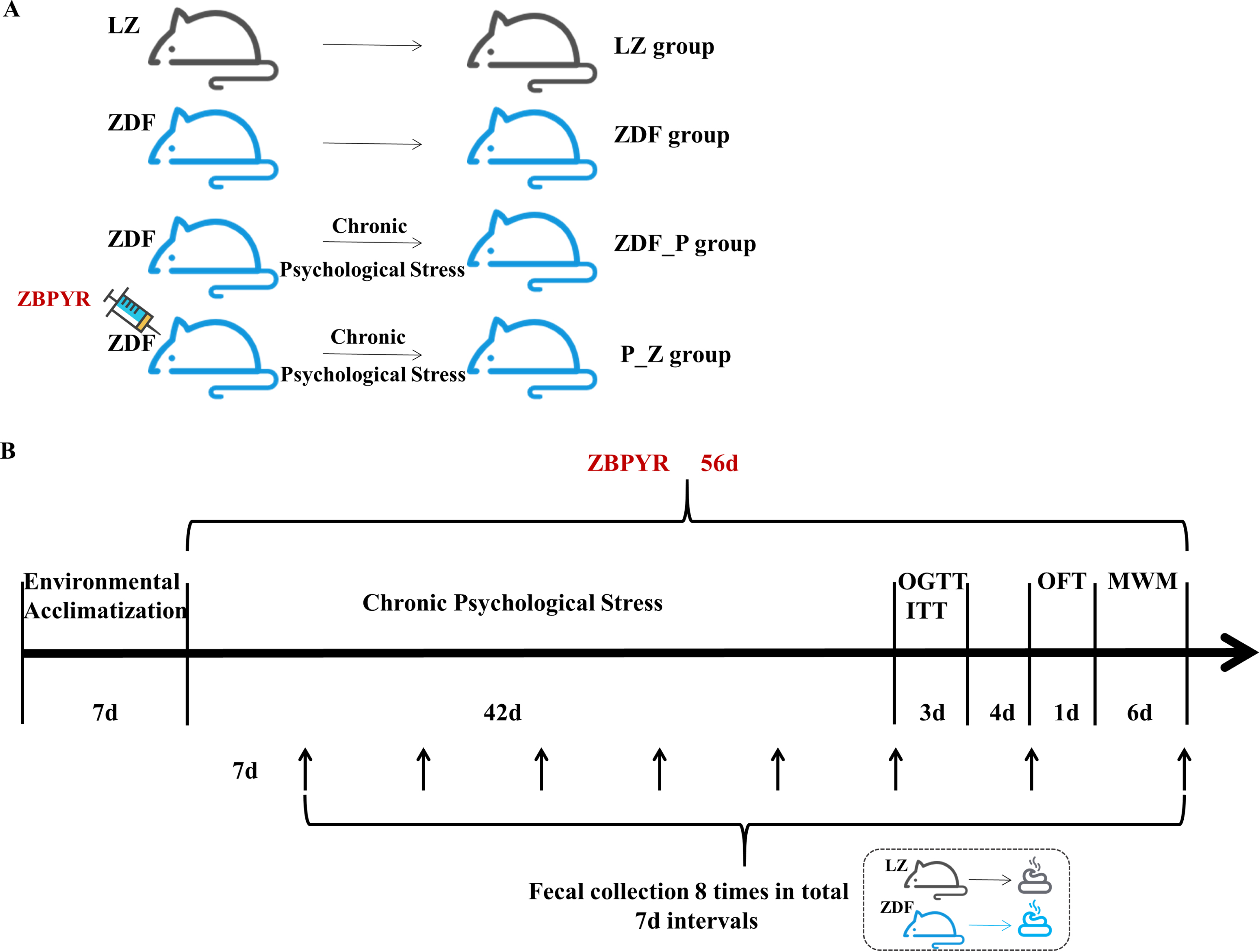
Experimental process. A) The experimental groups. B) The experimental operations.
Gut microbiota analysis
We studied the microbiomes of animals at 1-week intervals starting from week 1 onward to week 8 of the experiment. Fecal microorganisms were analyzed as described previously [36]. The V3-V4 region of the 16 S rRNA gene was sequenced using Illumina MiSeq. The sequencing data were further processed and evaluated via quantitative microbial ecology analysis (QIIME, v1.8.0) and the R software package (v3.2.0). In an ecosystem, an adapted ecological guild consists of species with similar functions and requirements. The same guild species have been found to be co-abundant. The SparCC algorithm was used to calculate the correlations between the core operations taxonomic units (OTUs). The core OTUs refer to the OTUs present in over 20% of the samples. The SparCC correlation coefficient matrix clustered the 340 core OTUs into 51 co-abundance groups (Modules). The average of the relative abundance of OTUs in a module is the abundance of that module.
Statistical analysis
Data are presented as means±SD and statistically analyzed by GraphPad Prism 8.0 software (GraphPad Software, San Diego, CA). Significant differences among different ages were analyzed by repeated ANOVA, and univariate analysis of significant differences among groups by one-way ANOVA followed by Tukey’s post hoc test. Wilcoxon’s signed-rank test was used to compare two groups. Spearman’s correlation analysis was used to test the correlations between physiological characteristics data and genera. A p < 0.05 was considered significant.
RESULTS
ZBPYR alleviates hyperglycemia, inhibits HPA axis activation, and improves behavioral and cognitive function
To establish a model of DACD aggravated by chronic PS, ZDF rats were subjected to chronic PS (restraint, rotation, and congestion). We analyzed the BW (Supplementary Table 1) and RBG (Supplementary Figure 1) data of rats in the Supplementary Materials. Chronic PS significantly and dynamically promoted weight gain as well as elevated blood glucose levels in rats. In the OGTT and ITT, the blood glucose levels and the area under the curve (AUC) of ZDF_P rats were higher than those of ZDF rats (Fig. 2A–D). ZBPYR significantly improved RBG and enhanced insulin sensitivity after 8 weeks of administration.

ZBPYR significantly improved glucose tolerance, insulin sensitivity and the levels of corticosterone in chronic PS-aggravated DACD rats. A-B) OGTT. A) Blood glucose levels, B) OGTT AUC. C, D) ITT. C) Blood glucose levels, D) ITT AUC. E) Corticosterone levels. Data are presented as means±SD. *p < 0.05, **p < 0.01, ***p < 0.001 compared to LZ; #p < 0.05, # #p < 0.01, # # #p < 0.001 compared to ZDF; and Δ p < 0.05, ΔΔ p < 0.01 compared to ZDF_P. ZBPYR, ZiBu PiYin Recipe; PS, psychological stress; DACD, diabetes-associated cognitive decline; OGTT, oral glucose tolerance test; AUC, area under the curve; ITT, insulin tolerance test.
To verify whether the HPA axis was activated, we measured serum corticosterone levels using ELISA. Corticosterone levels were higher in ZDF_P rats compare to ZDF rats (Fig. 2E). Administration of ZBPYR significantly decreased the levels of corticosterone, suggesting that ZBPYR regulates HPA axis activation.
ZDF rats exposed to chronic PS displayed a variety of characteristic anxiety-like and depression-like behaviors, such as hyperactivity, hair standing up, and lethargy. The OFT assessed spontaneous locomotor and anxiety-like behavior in rats. The number of entries in center, time in center and vertical numbers were reduced in ZDF_P rats compared to ZDF rats (Fig. 3A–D). The behavior of P_Z rats was restored after ZBPYR administration, indicating that ZBPYR improved anxiety-like behavior in rats given chronic PS.
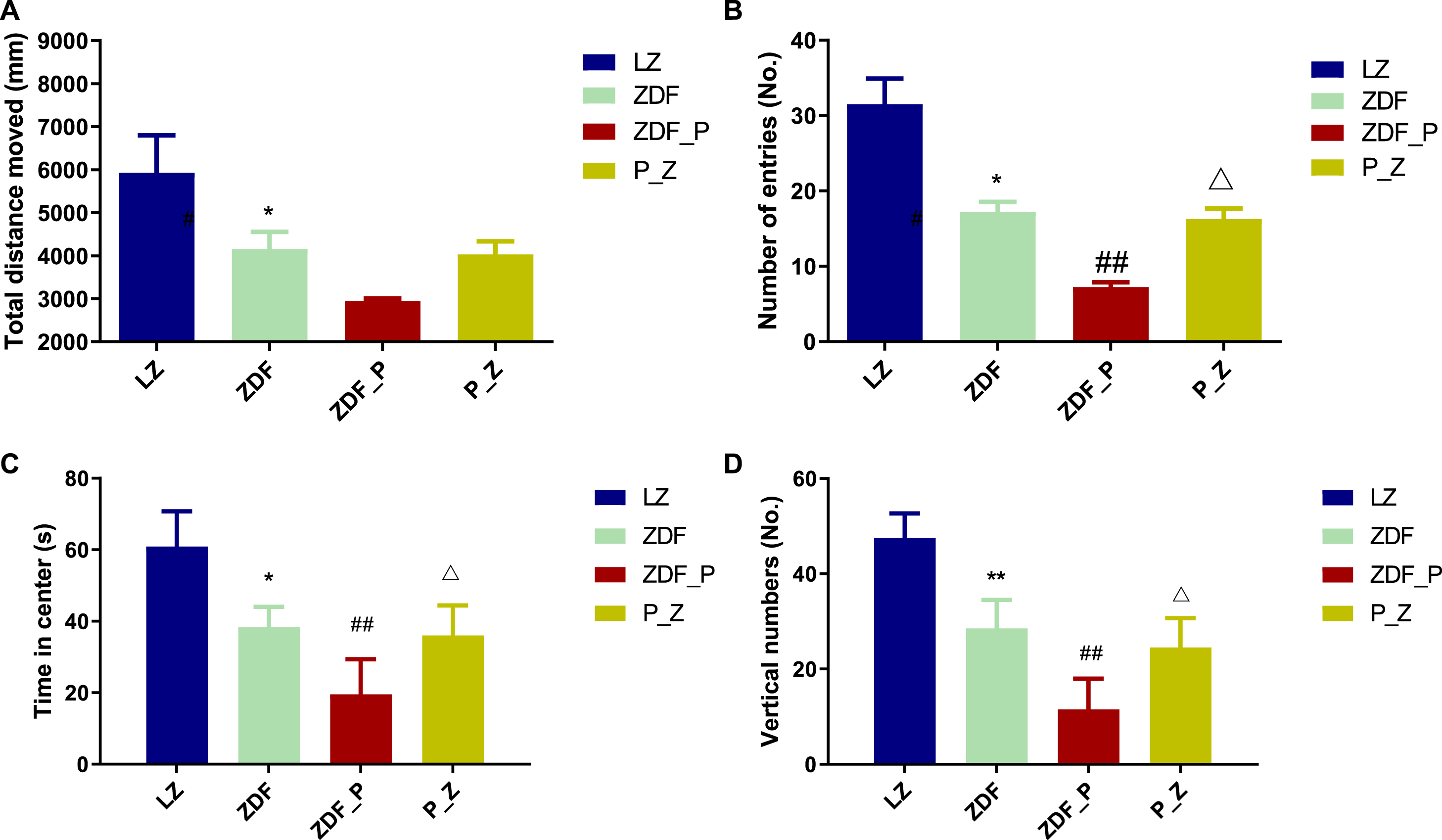
ZBPYR improved the performance of chronic PS-aggravated DACD rats in the OFT. A) Total distance. B) Numbers of entries into the center. C) Time in center. D) Vertical numbers. *p < 0.05, **p < 0.01 compared to LZ; # #p < 0.01 compared to ZDF; and Δ p < 0.05 compared to ZDF_P. OFT, open field test.
In the last week of the experiment, MWM was performed to assess the learning and memory capacity of rats. Compared with ZDF rats, the ZDF_P group had a longer escape latency on day 5 (Fig. 4A), a longer time to find the original platform position (Fig. 4B), and a shorter time to stay in the quadrant of the original platform position (Fig. 4C). Administration with ZBPYR significantly shortened the time for the rats to find the location of the original platform and prolonged the time that the rats stayed in the quadrant of the original platform (Fig. 4B, C). In the search for visible platform experiment, the data did not differ significantly (Fig. 4E, F), indicating that T2DM and PS did not affect the locomotor ability and visual acuity of ZDF rats. All these results suggested that ZBPYR significantly ameliorated cognitive decline in the P_Z group.
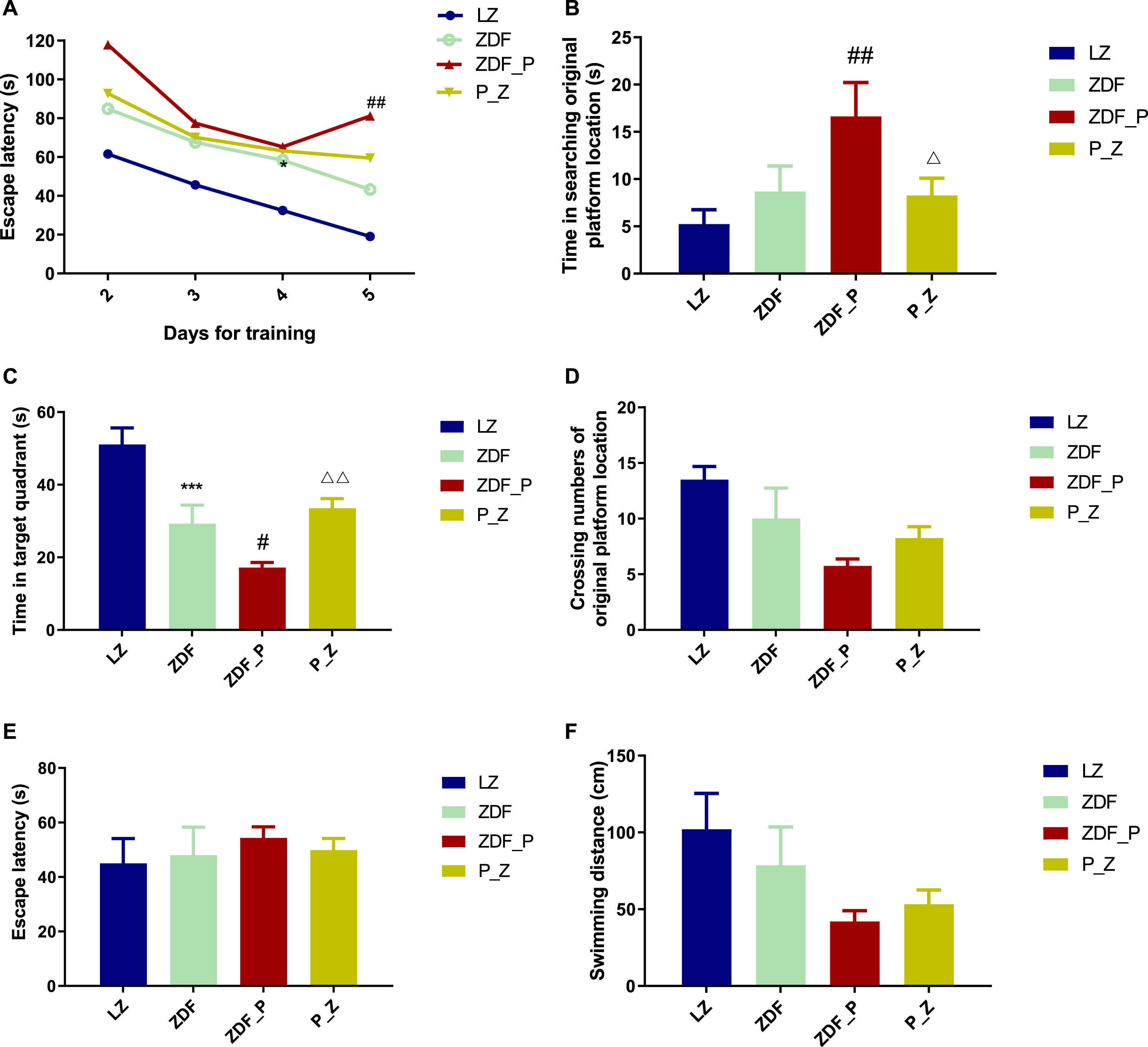
ZBPYR improved the performance of chronic PS-aggravated DACD rats in the MWM test. A) Escape latency in the orientation navigation test. B-D) The spatial exploration test. B) time in searching original platform location, C) time in target quadrant, and D) crossing numbers of original platform location. E, F) The visible platform test. E) escape latency, and F) swimming distance. ***p < 0.001 compared to LZ; #p < 0.05, # #p < 0.01 compared to ZDF; and ΔΔ p < 0.01 compared to ZDF_P. MWM, Morris water maze.
Dynamic changes of the gut microbiota by ZBPYR
To analyze the dynamic gut microbiota alteration in DACD rats aggravated by chronic PS administered with ZBPYR, we performed Illumina sequencing on fecal samples collected during experimental weeks 1–8 (that is, in rats aged 8–15 weeks) in the LZ, ZDF, ZDF_P, and P_Z groups. Altogether, we obtained 7,786,460 high-quality sequences and 2,419 OTUs from 128 samples, with an average of 60,831±11,342 reads. During the 1–8 weeks of the experiment, nine bacterial phyla were reserved for analysis (Fig. 5A, Supplementary Figure 2). The predominant bacterial phyla were Firmicutes and Bacteroides, and the predominant bacterial genus was Lactobacillus. The pattern (representing the relative positions of 32 sample points at different times) of microbiome trajectories (Fig. 5B–D) showed that the microbial tracks of ZDF and ZDF_P rats were similar, while those of the P_Z rats were opposite. The alpha diversity of the intestinal flora showed that ZBPYR significantly and dynamically decreased Simpson’s and Shannon’s diversity indices at weeks 7–8 of the experiment, indicating that ZBPYR significantly decreased diversity with the extension of administration time (see Supplementary Figure 3). The relative abundance of phyla and genera in the sample groups at eight time points (Fig. 5E, Supplementary Figures 4 and 5) was used to further evaluate changes in the intestinal microbiome. Not surprisingly, the high and low abundance variables changed greatly. All the bacterial phyla and genera changed significantly with time. Phylum-level profiling demonstrated that chronic PS increased Proteobacteria abundance in 15-week-old rats, while ZBPYR dynamically increased Firmicutes abundance and decreased Actinobacteria abundance in rats. Genus-level analysis revealed that chronic PS significantly increased Alistipes and Coprococcus abundance dynamically, and increased Proteus abundance in 15-week-old rats, while ZBPYR dynamically decreased Alistipes and Coprococcus abundance in 13- to 15-week-old rats, decreased Holdemania and Proteus abundance in 15-week-old rats, and increased Blautia abundance in 12–13-week-old rats. Thus, chronic PS-induced changes in intestinal flora trajectories in DACD rats were similar to those in the ZDF group, while ZBPYR dynamically altered intestinal flora in chronic PS-aggravated DACD rats.
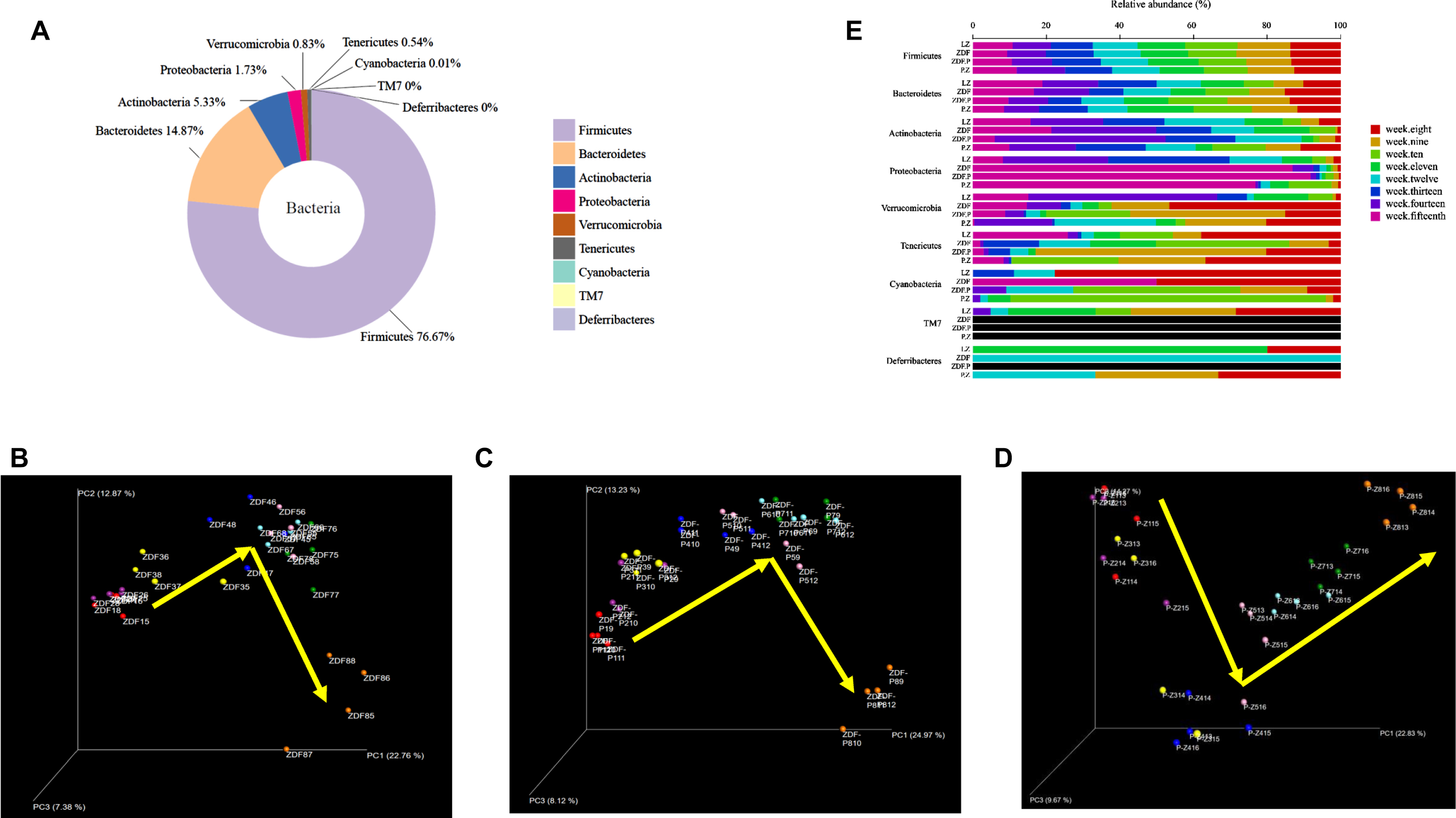
Pattern of changes in the structure of fecal microbial composition in rats during weeks 1–8 of the experiment. A) Analysis of fecal microbial composition. B-D) Correlation trajectories of fecal microbial changes over time assessed by unweighted UniFrac PCoA principal coordinate analysis B) for ZDF, C) for ZDF_P, and D) for P_Z. E) Relative abundance distribution of fecal bacterial composition at each age group (8–15 weeks of age) of rats at major phyla levels.
Changes in Modules correlate with dynamic changes in BW and glucose
In order to find the key phenotypes related to the efficacy of ZBPYR, we performed co-abundant network analysis methods. The 340 core OTUs were clustered in 51 Modules by SparCC correlation (Supplementary Table 2). In the ZDF group, 23 Modules increased, and 9 Modules decreased compared to the LZ group (Supplementary Figure 6). 8 Modules increased and 6 Modules decreased in ZDF_P compared to ZDF (Fig. 6). In P_ Z, 6 Modules were increased, and 13 Modules were decreased compared to ZDF_P (Fig. 7).
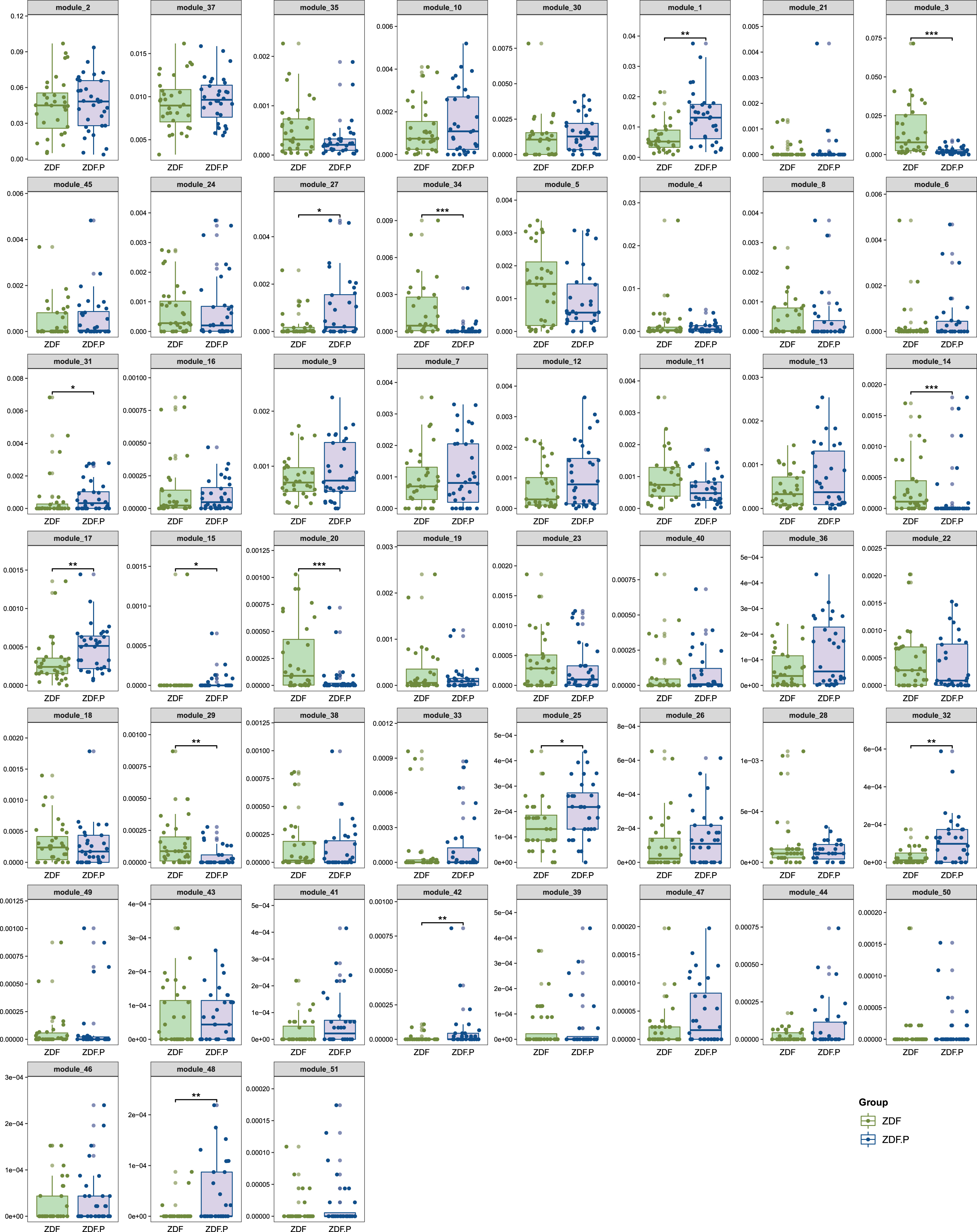
Altered modules by ZDF and ZDF_P. *p < 0.05, **p < 0.001, ***p < 0.001.

Altered modules by ZDF_P and P_Z. *p < 0.05, **p < 0.01.
In ZDF_P, enriched Module 1, Module 17, and Module 48 were significantly positively correlated with BW (Fig. 8A–C), enriched Module 25 was negatively correlated with BW (Fig. 8D), and inhibited Module 14, Module 20, Module 29, and Module 42 were significantly negatively correlated with BW (Fig. 8E–H). Module 48 was significantly positively correlated with RBG (Fig. 8I), and Module 29 was significantly negatively correlated with RBG (Fig. 8J). Module 1 contained one OTU from each of genera Dorea, Blautia, and Ruminococcus. Module 14 contained one OTU from unErysipelotrichaceae. Module 17 contained an OTU from unLachnospiraceae and unClostridiales. Module 20 contained ten OTUs from unRuminococcaceae, two OTUs from Lactobacillus and unClostridiales, one OTU from SMB53. Module 25 contained an OTU from Lactobacillus. Module 29 contained three OTUs from unClostridiales. Module 42 contained one OTU each from Akkermansia and unPeptostreptococcaceae. Module 48 contained an OTU from Blautia and unLachnospiraceae.
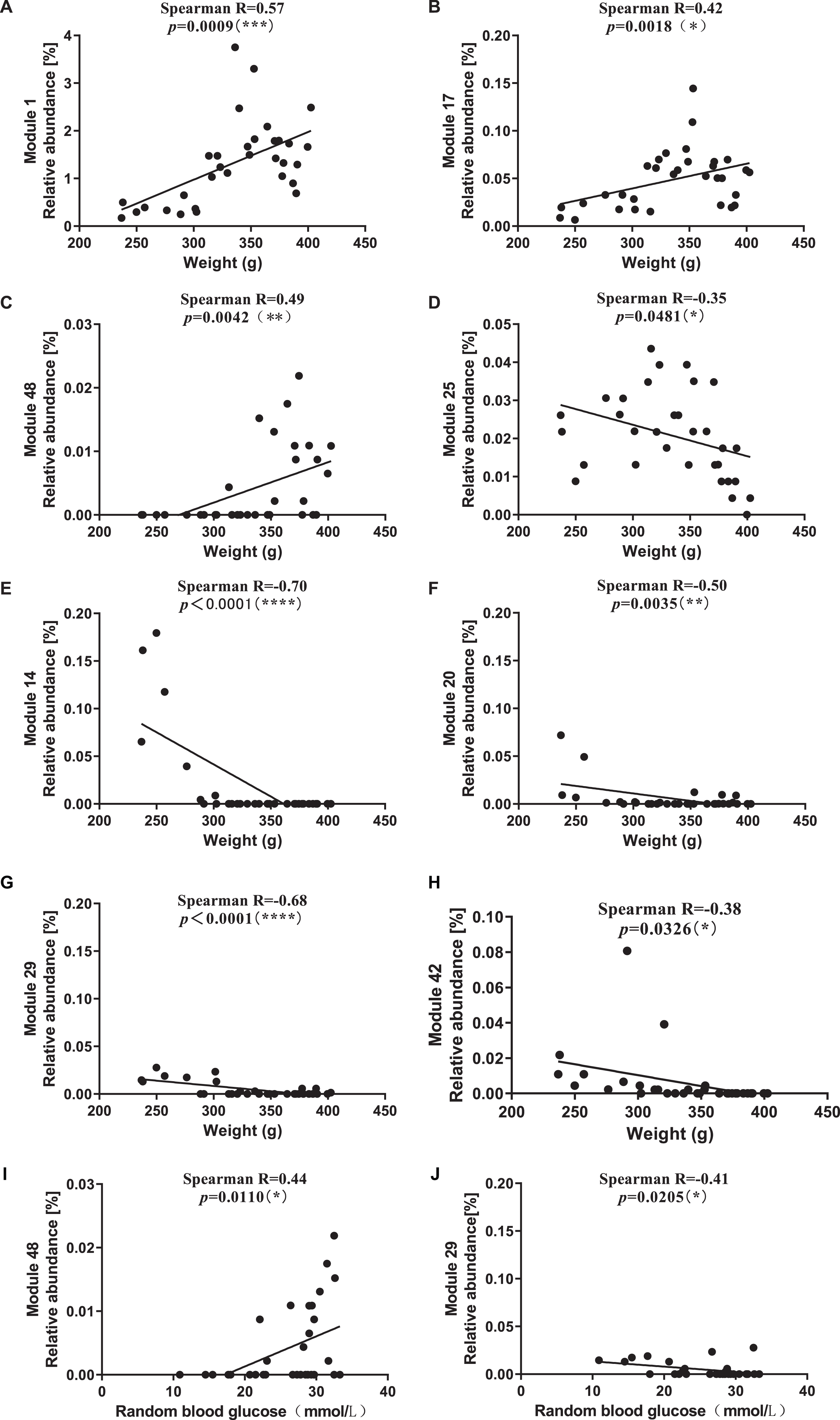
Correlation between modules related to chronic PS and physiological characteristics of rats. A-H) Modules and BW. A) Module 1, B) Module 17, C) Module 48, D) Module 25, E) Module 14, F) Module 20, G) Module 29, H) Module 42. I, J) Modules and RBG. I) Module 48, J) Module 29. BW, body weight. RBG, random blood glucose.
Among the six Modules significantly enriched by ZBPYR, Module 3 and Module 45 were significantly correlated with elevated BW (Fig. 9A, B). Module 3 included two OTUs from unClostridiales, and an OTU from Ruminococcus. Module 45 contained seven OTUs from unRuminococcaceae, four OTUs from unClostridiales, two OTUs from Lactobacillus, and an OTU from Coprococcus, SMB53, and unLachnospiraceae. Among the thirteen Modules significantly inhibited by ZBPYR, Module 13 and Module 17 were significantly positively correlated with BW (Fig. 9C, D), Module 16, Module 23, Module 43, and Module 46 were significantly negatively correlated with BW (Fig. 9E–H), and Module 2 was significantly positively correlated with RBG (Fig. 9I). Module 13 contained one OTU from unLachnospiraceae. Module 16 contained eleven OTUs from unClostridiaceae, six OTUs from unPeptostreptococcaceae, five OTUs from Trichococcus, four OTUs each from Lactobacillus and Clostridium, three OTUs each from unRuminococcaceae and unClostridiales, two OTUs from 02d06, one OTU each from Turicibacter, unS24-7, and un[Mogibacteriaceae]. Module 23 contained one OTU each from Bifidobacterium and unPeptostreptococcaceae. Module 43 contained one OTU each from Clostridium and unClostridiaceae. Module 46 contained two OTUs from unClostridiaceae; and Module 2 contained three OTUs from Lactobacillus.
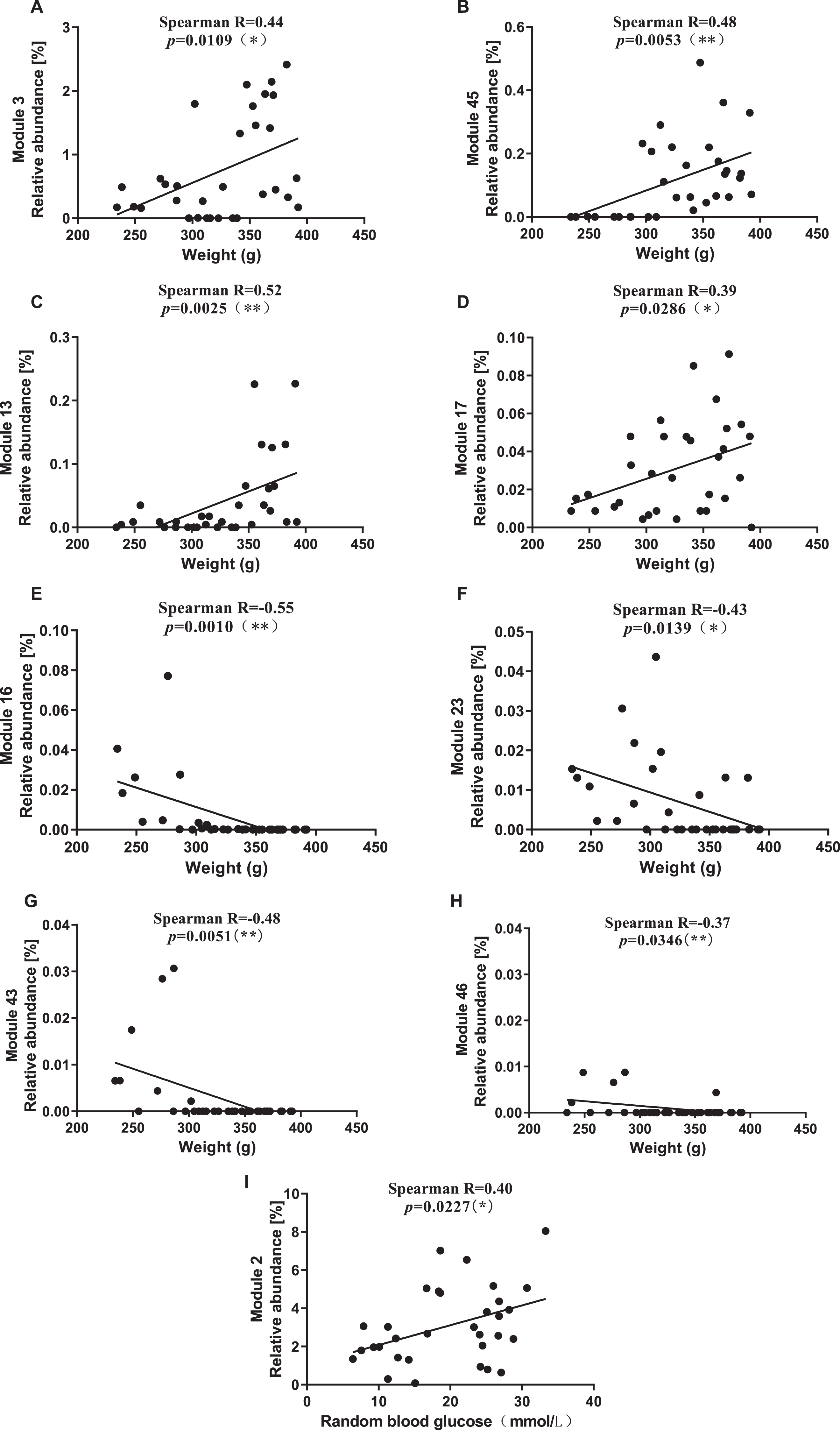
Correlation between modules related to ZBPYR intervention and physiological characteristics of rats. A-H) Modules and BW. A) Module 3, B) Module 45, C) Module 13, D) Module 17, E) Module 16, F) Module 23, G) Module 43, H) Module 46. I) Module 2 and RBG.
We compared the key Modules of the four groups and attempted to find common Modules among them. These Modules may not simply respond to pharmacological treatment, but are likely to be related to the alleviation of DACD aggravated by chronic PS. The common modules in the four groups were Module 17 and Module 25, and the representative genus was Lactobacillus.
DISCUSSION
This is the first research to observe the dynamic intervention influence of TCM on the gut microbiota of rats with chronic PS-aggravated DACD. In 8 weeks of animal experiments, chronic PS promoted and exacerbated DACD phenotypes, consistent with previous studies showing that stress negatively affects cognitive performance [12, 37], while ZBPYR significantly ameliorated hyperglycemia and cognitive function, consistent with our previous animal studies [3, 34].
In our animal experiments, chronic PS and ZBPYR were observed to dynamically alter the gut microbial structure. After administration of chronic PS, the abundance of Alistipes and Coprococcus were significantly and dynamically elevated, while the abundance of Proteus was significantly elevated in 15-week-old rats. One study has found a negative correlation between Alistipes and MoCA scores in patients with mild cognitive disorder [38]. An increased abundance of Alistipes is found in chronic fatigue syndrome [39]. Some patients with elevated Coprococcus levels are more likely to benefit from prebiotic supplementation [40]. Proteus can cause neuroinflammation by producing lipopolysaccharide, with increased abundance of the genus in patients with Alzheimer’s disease [41]. Infants exposed to higher stress tend to have higher Proteus. ZBPYR dynamically reduced the abundance of Alistipes and Coprococcus in 13–15-week-old rats, significantly reduced the abundance of Holdemania and Proteus in rats of 15 weeks age. Holdemania is a gram-positive anaerobic genus [43], and exhibits moderate correlation with anxiety, mainly in major depressive disorder patients [44]. These data show that the above-mentioned genera may play a role in ZBPYR alleviating PS-aggravated DACD.
Chronic PS significantly increased functional groups Module 1, Module 17, and Module 48, which contain OTUs from Dorea, Blautia, and Ruminococcus. The above genera are shown to be associated with high-fat diets [45]. A higher abundance of Dorea is found in stress-prone Wistar-Kyoto rats [46]. Moreover, the genus Dorea may promote precise regulation of the entire microbial ecosystem, leading to downregulation of intestinal mucosal inflammation. This effect may be related to the regulation of stress response [47]. As a genus of the phylum Firmicutes and the family Lachnospiraceae, an increased trend of Blautia has been reported in mice subjected to chronic stress [48]. Blautia is positively correlated with stress indicators dopamine beta hydrolase and tryptophan 2,3-dioxygenase [49]. In many central nervous system diseases, abnormally abundant Blautia has been observed [50]. An increased abundance of Ruminococcus is found in mice exposed to chronic restraint stress [51–53]. Ruminococcus has been found to be positively associated with four cognitive functions (semantic fluency span, digit span backward, memory domain, and recognition test) in elderly patients with mild cognitive impairment [54]. Modules, which were significantly suppressed by chronic PS, contain OTUs from Lactobacillus, Akkermansia, and SMB53. Unpredictable restraint stress decreases the abundance of the anti-inflammatory bacteria Lactobacillus [55, 56]. One study shows that oral administration of Lactobacillus in chronically stressed animals improves behavior, and cognition [57]. Lactobacillus johnsonii BS15 can be used to treat memory loss caused by PS [58]. There is evidence that Akkermansia effectively improves glucose tolerance, dyslipidemia, and intestinal barrier dysfunction and relieves the disorder of learning and memory in Alzheimer’s disease [59]. Akkermansia was significantly reduced and positively correlated with behavioral indicators of depression and anxiety in socially defeated animals [60, 61]. The SMB53 genus is increased in mice that spontaneously develop diabetes [62].
Module 3 and Module 45, which were significantly enriched after ZBPYR intervention, contain OTUs from Ruminococcus, Lactobacillus, Coprococcus, and SMB53. Module 2, Module 13, Module 16, Module 23, Module 43, and Module 46, which were significantly suppressed after ZBPYR intervention, contain OTUs from Lactobacillus, Trichococcus, 02d06, Clostridium, Bifidobacterium, and Turicibacter. Several reports have suggested that increased abundance of Lactobacillus is critical to the obesity epidemic. Likewise, reports have illustrated high Lactobacillus abundance in T2DM contributes to the chronic inflammation [36]. Nevertheless, a large body of literature still defines Lactobacillus as a probiotics. Trichococcus currently contains nine species, and in general, species of the genus Trichococcus can degrade a wide range of carbohydrates [63]. Participants with diabetes have a higher relative abundance of 02d06 [64]. Clostridium can induce diarrhea [65] and infection [66], and the genus contains several highly pathogenic species that produce toxins [67]. It has been suggested that the ratio of Clostridium to Lactobacillus may be associated with metabolic disturbances in chronically unpredictable mildly stressed animals [67]. One study has revealed increases of Clostridium species in chronically stressed animals [68]. Turicibacter is associated with high-fat diets [69, 70]. The family Trichodermaceae also increased with high-fat diets [46, 71]. Lactobacillus and Bifidobacterium improve cognitive performance in elderly mice through modulation of the intestinal microbiota [72, 73]. A correlation between gut flora abundance and corticosterone was not observed in our results, which is consistent with previous studies findings that certain gut flora abundances were independent of corticosterone, showing that biological effects of stress may go beyond changes in corticosterone [57]. The lack of observation of other stress indicators is a shortcoming of our study. Another weakness is that our results are based on a small sample size. In addition, the diversity of intestinal flora is easily affected by environmental, genetic, gender and other factors, which may be responsible for the differences in analysis results [52]. In the future study, we will supplement and improve our methods for evaluating animal cognitive behavior, such as using the elevated plus maze (EPM) test to detect anxiety-like behavior in animals. Furthermore, we would continue to observe in depth whether the hyperglycemia and cognitive decline of the ZDF rats without chronic PS can be better ameliorated after ZBPYR administration compared to ZDF rats with chronic PS. It would be interesting if we could study whether chronic PS can cause cognitive decline and gut flora interventions in the LZ rats and the function of ZBPYR on LZ rats with chronic PS. The study would reflect the majority situation nowadays because a huge population is under chronic PS. Additionally, further validation in a larger number of animals or a human cohort is needed.
Notably, Module 17 and Module 25, representing the genus Lactobacillus, were common to all four groups and were affected by ZBPYR intervention. Lactobacillus could be a target of ZBPYR for preventing and treating chronic PS-aggravated DACD. ZBPYR exerted a good modulatory effect on the gut microbiota and improved both hyperglycemia and cognitive functions, which might have been caused by the multiple chemical components of ZBPYR, such as ginsenosides, Salvia miltiorrhiza polysaccharides, cryptotanshinone, and Poria cocos polysaccharides. Panax ginseng C. A. Mey. is now often used as a dietary supplement, and clinically it can also treat diabetes and inflammatory diseases [74]. Ginsenosides are its primary active components and have various biological effects, such as anti-obesity and anti-hyperglycemia effects. It has been found to maintain energy balance, regulate intestinal ecological dysbiosis, improve intestinal integrity and metabolic inflammation, and accelerate the recovery of the probiotic Lactobacillus [75–77]. Salvia miltiorrhiza polysaccharides, cryptotanshinone, and Poria cocos polysaccharides can enhance the function of probiotics Lactobacillus and improve disorders of glucolipid metabolism and insulin resistance [78–80]. These findings imply that ginsenosides, Salvia miltiorrhiza polysaccharides, cryptotanshinone, and Poria cocos polysaccharides may be the main active components in ZBPYR that alter the intestinal microbiota. Further work to figure out which are the most useful components of the ZBPYR that attenuate the hyperglycemia and cognitive decline as well as alter the intestinal flora is important.
Conclusions
We found that chronic PS can activate the HPA axis, affect behavior, and promote cognitive abnormalities, and that the intervention of ZBPYR ameliorated these disturbances. Microbiomic analysis showed that at the genus level, chronic PS significantly and dynamically elevated the abundance of Alistipes and Coprococcus; enriched Module 1 (Dorea, Blautia, Ruminococcus) and Module 48 (Blautia); and inhibited Module 20 (Lactobacillus, SMB53), and Module 42 (Akkermansia). These genera may play a significant role in DACD aggravated by chronic PS, and Lactobacillus, Alistipes, and Coprococcus may be important targets of ZBPYR in preventing and treating chronic PS-aggravated DACD (Fig. 10).

Schematic showing the effect of chronic PS and ZBPYR for genera and Modules (representative genera), respectively. Red arrows represent upregulation, green arrows represent down-regulation.
Footnotes
ACKNOWLEDGMENTS
This work was financially supported by The Key Project of National Natural Science Foundation, China (81230084), Traditional Chinese Medicine Inheritance and Innovation “Hundreds and Tens of Thousands” Talent Project (Qihuang Project) ([2022] No. 6), the Natural Science Research of Jiangsu Higher Education Institutions (21KJB360003), the Open fund of Key Laboratory of Ministry of Education for TCM Viscera-State Theory and Applications, Liaoning University of Traditional Chinese Medicine (zyzx2107).