
Abstract
Background:
Apolipoprotein E (APOE) is closely related to Alzheimer’s disease and other age-related diseases. In recent years, several studies have shown an interaction of APOE by age on brain volume. However, validation in larger cohorts is required.
Objective:
We explored the age-related effect of APOE on brain volumes in a community-dwelling cohort.
Methods:
Inhabitants in Shunyi District in Beijing aged≥35 years were invited to join this study from 2013 to 2016. The baseline assessments, APOE genotyping and brain magnetic resonance imaging were performed. Neuroimaging small vessel disease characteristics and brain volumes (global measures, cerebral lobes, hippocampus, brainstem, and subcortical nuclei) were acquired. The general linear model was used to analyze the interaction of APOE genotypes by age on brain volumes, and the age of 60 years was chosen as a cut-off value for stratification analysis.
Results:
A total of 1,105 subjects were enrolled in the final analysis with a mean age of 56.18 (9.30) years, and 37.7% were men. APOE ɛ3/ɛ3 carriers account for 71.8%, ɛ2 (+) 14.0%, and ɛ4 (+) 14.2%. Compared with APOE ɛ3/ɛ3, a significant protective effect for APOE ɛ4 (+) on brain parenchyma fraction (β = 0.450, p = 0.048) was observed in subjects aged≤60 years; in participants aged > 60 years, a negative effect for APOE ɛ4 (+) on hippocampus (β = 1.087, p = 0.021) was found.
Conclusion:
Our study reveals that APOE ɛ4 has differential effects on cerebral structures in different stages of lifespan, suggesting its complicated biological function and underlying antagonistic pleiotropy.
INTRODUCTION
Population aging is a global issue confronted by both developed and underdeveloped countries. Aging-related diseases, such as Alzheimer’s disease (AD), have posed challenges on public health care expenditure and social development [1], and the prevalence of dementia is projected to triple by 2050 [2]. Apolipoprotein E (APOE) is the strongest genetic risk factor for late-onset AD with the age of onset dominantly above 65 [3]. APOE ɛ4 heterozygotes increase the risk of AD by 3.7 folds and APOE ɛ4 homozygotes have a 12-fold increased risk compared with the ɛ3/ɛ3 genotype, while APOE ɛ2 is a protective genetic factor for AD [3]. APOE gene is located in chromosome 19, with three alleles of APOE ɛ2, ɛ3, and ɛ4 encoding three isoforms including APOE2 (Cys112; Cys158), APOE3 (Cys112; Arg158), and APOE4 (Arg112; Arg158) [4]. ApoE is a 34-kDa glycoprotein composed of 299 amino acids, and plays an important role in lipid metabolism, neurotrophy, synaptic functions, innate immunity, and blood-brain barrier integrity maintenance [4].
\enlargethispage 2pt In a lot of previous studies, APOE ɛ4 was found to be associated with greater rates of hippocampal atrophy and memory loss in AD patients [5, 6] and non-demented community-dwelling adults [7–10]. These findings suggest that APOE starts to modulate cerebral structures and functions long before the emergence of clinically overt symptoms. Furthermore, it is speculated that the effect of APOE might be exerted throughout the lifespan as early as starting from the fetal period [11].
In recent years, more and more studies have found an interaction between APOE genotype and age, suggesting that the influence of APOE might depend on the age of individuals [12]. In 2008, Mueller et al. showed significant effects of APOE ɛ4 on specific hippocampal zones in participants with older age, but not in the younger subjects [13]. A significant interaction of APOE genotype by age on grey matter volume in left calcarine, left insula, and left medial frontal gyrus in mild cognitive impairment (MCI) patients was reported by Chen in 2015 [14]. In 2018, Cacciaglia group found significant interactions between APOE ɛ4 homozygotes and age in the grey matter volume of the right hippocampus, caudate, precentral gyrus and cerebellar crus, and lower grey matter volume in homozygotes were observed after the fifth decade of life [15]. In 2020, Martí-Juan et al. found that nonlinear interaction between APOE ɛ4 allele load and age modulated the local three-dimensional hippocampal surface changes in cognitively intact individuals [16]. However, concerning the limited number of previous research, there are still knowledge gaps in the exact interaction between APOE genotype and age, requiring validation in larger cohorts. Whether the effect of APOE on human beings depends on different age stages needs to be put more emphasis on to better understand the underlying pathogenetic mechanism of APOE gene.
In this study, we aimed to investigate the prevalence of APOE genotypes in a large Chinese community-dwelling population, and the age-related effect of APOE genotype on brain volumes was studied. We hypothesized that APOE genotype might have differential effects on brain volumes in different age stages.
METHODS
Study participants
All participants were enrolled from Shunyi Study, an ongoing community-based cohort study conducted in Shunyi District which is a suburban area near Beijing. Inhabitants aged 35 years and older who could live independently were invited in this study from June 2013 to April 2016. All participants signed an informed consent form. The research was approved by the Medical Review Ethics Committee of Peking Union Medical College Hospital (Reference number: B-160).
Of the 1,586 individuals participating in this study, 70 participants with prior stroke, 251 without brain imaging data, and 143 without APOE genotype data were excluded. A total of 17 participants with APOE ɛ2/ɛ4 were excluded considering the contradictory effects of APOE ɛ2 and ɛ4 on AD.
There were 1,105 participants entered into the final analysis. Fig. 1 shows the flow chart of this study.
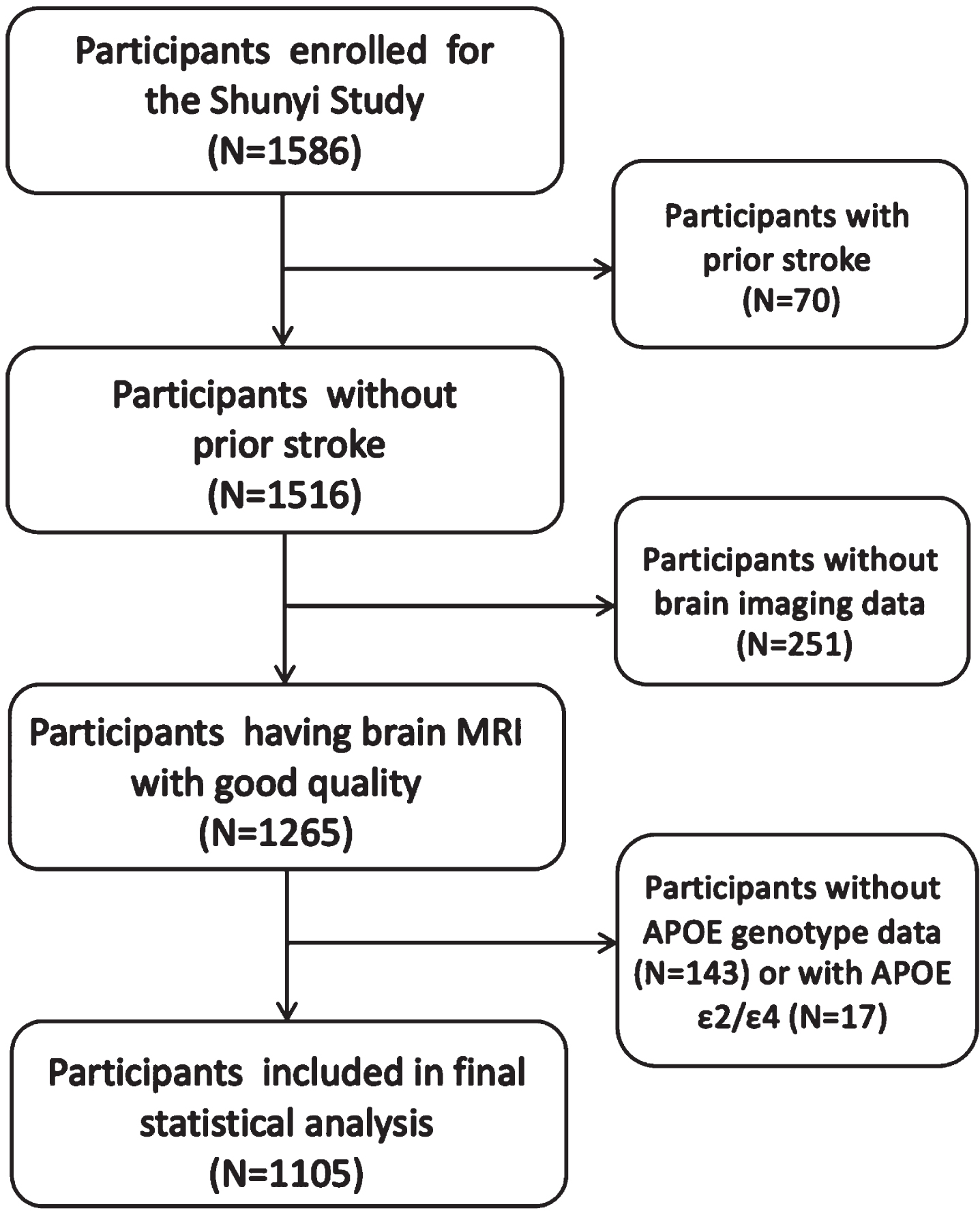
Study flow chart. MRI, magnetic resonance imaging; APOE, Apolipoprotein E.
Data collection
Demographic and clinical data
The baseline assessments for each participant included structured questionnaires, physical examinations, laboratory tests, and cognitive function test. The definition of cerebrovascular risk factors was as follows: hypertension was defined as systolic blood pressure≥140 mmHg or diastolic blood pressure≥90 mmHg; diabetes mellitus was defined as fasting serum glucose≥7.0 mmol/L or glycated hemoglobin (HbA1c) ≥6.5%; hyperlipidemia was defined as total cholesterol > 5 mmol/L or low-density lipoprotein cholesterol≥3.62 mmol/L; smoking status was categorized as current smoker and non-current smoker. The Mini-Mental State Examination (MMSE) was used to assess cognitive function.
APOE genotyping
Genomic deoxyribonucleic acid (DNA) of each participant was extracted from whole blood samples using a standard protocol. The three alleles of APOE, including ɛ2, ɛ3, and ɛ4, were defined by two single nucleotide polymorphisms (SNPs) including rs429358 and rs7412 [17]: ɛ2, rs429358 (T) and rs7412 (T); ɛ3, rs429358 (T) and rs7412 (C); ɛ4, rs429358 (C) and rs7412 (C). APOE ɛ4(+) was identified as APOE ɛ3/ɛ4 and APOE ɛ4/ɛ4; APOE ɛ2(+) was identified as APOE ɛ2/ɛ2 and APOE ɛ2/ɛ3. APOE ɛ2/ɛ4 were not included in this study for the reason mentioned above.
MRI acquisition and postprocessing
Brain magnetic resonance imaging (MRI) was performed using a single 3-T Siemens Skyra scanner (Siemens, Erlangen, Germany). Three-dimensional T1-weighted image (T1WI), T2-weighted image (T2WI), fluid-attenuated inversion recovery (FLAIR), and susceptibility-weighted imaging (SWI) were obtained. The detailed parameter settings of MRI sequences have been published in previous studies of Shunyi cohort [18].
White matter hyperintensities (WHMs) were defined as symmetrical hyperintensities on T2WI and FLAIR sequences and isointensities or faint hypointensities on T1WI sequences. WHMs were automatically segmented by the lesion growth algorithm as implemented in the LST toolbox (http://www.statistical-modeling.de/lst.html) for SPM at κ = 0.15. Lacunes were defined as round or ovoid focal lesions with a diameter ranging from 3–15 mm, usually in basal ganglia or white matter, representing the same signal characteristics as cerebrospinal fluid (CSF) on all sequences. Cerebral microbleeds (CMBs) were the round or ovoid structures with black or very hyperintense signal observed on SWI sequences, ranging from 2–10 mm in diameter, devoid of hyperintensity on TIWI and T2WI sequences, and at least half was surrounded by brain parenchyma [19]. Enlarged perivascular spaces (PVS) were CSF-like signal lesions with a diameter < 3 mm, located in perforating artery territories, which could be linear, ovoid, or round and the boundary was clear. The four-level severity score of PVS located in white matter (PVS-WM) are as follows: degree 1, < 10 PVS in the total white matter volume; degree 2, > 10 PVS in the total white matter volume and < 10 PVS in the slice containing the greatest number of PVS; degree 3, 10–20 PVS in the slice containing the greatest number of PVS; degree 4, > 20 PVS in the slice containing the greatest number of PVS. Enlarged PVS in basal ganglia (PVS-BG) were rated as follows: degree 1, < 5 PVS; degree 2, 5–10 PVS; degree 3, > 10 PVS but still numerable; degree 4, innumerable. Severe PVS-WM or PVS-BG was defined as degree≥2 and degree≥3 respectively [20].
An automated segmentation using three-dimensional TIWI was performed by CIVET pipeline [21] and yielded the following volumetric measures: the intracranial volume (ICV), total brain volume, whole-brain gray matter volume (GMV), whole-brain white matter volume (WMV) and cortical gray matter volume. The brain parenchymal fraction (BPF) was defined as: BPF = (WMV+GMV)×100/ICV. Meanwhile, we chose a comprehensively wide range of human brain structures to study the interaction effects between APOE and age, which were extracted by using the FIRST algorithm embedded in FSL (FMRIB Software Library, v5.0) [22], including cerebral lobes (frontal, parietal, temporal, and occipital lobes), brainstem, hippocampus, and subcortical nuclei (caudate, pallidum, putamen, and thalamus). Bilateral volumes were summed. Volumes of GMV, WMV, and cortical GMV were normalized using the following formula: volumenorm = volumeraw×100/ICV. Volumes of lobes, brainstem, hippocampus and subcortical nuclei were normalized as: volumenorm = volumeraw×1000/ICV.
Statistical analysis
Means or medians were used to show the average levels of quantitative data with or without normal distribution. Variance analysis or rank-sum test was used to compare quantitative variables with or without normal distribution between several groups. Chi-square test was used to compare categorical variables between different groups.
Interaction effect analysis: Interaction effect between APOE genotypes and age was evaluated by general linear model (GLM) with each normalized volume parameter as the dependent variables and age/10, gender, APOE genotype [APOE ɛ3/ɛ3, APOE ɛ2 (+), APOE ɛ4 (+)] and age×APOE genotype as the independent variables. False discovery rate was used for multiple comparison correction in the subtypes of cerebral lobes and subcortical nuclei which contain more than one structure.
Stratification analysis
The effect of APOE genotypes on brain volume was analyzed by GLM, with each normalized volume parameter as the dependent variable, and age, gender, and APOE genotypes as the independent variables in different age groups. Since APOE is known as a key gene related with some age-related diseases like AD, and the year of 60 is defined by World Health Organization as the start of old age, we chose the age of 60 years as the cut-off value for stratification analysis. False discovery rate was used for multiple comparison correction in the subtypes of cerebral lobes and subcortical nuclei which contain more than one structure.
SAS version 9.4 (SAS Institute, Inc., Cary, NC, United States) was used for statistical analyses and the p-value < 0.05 were considered statistically significant.
RESULTS
Demographic information
The detailed demographic information of participants enrolled in the final analysis is summarized in Table 1. A total of 1,105 subjects were included in this study, with a mean age of 56.18±9.30 years and 37.7% were men. Of all participants, carriers of APOE ɛ3/ɛ3 account for 71.8%, ɛ2 allele carriers account for 14.0%, and ɛ4 allele carriers account for 14.2%. The average age of APOE ɛ3/ɛ3, APOE ɛ2 (+), and APOE ɛ4 (+) was 56.29±9.57, 56.41±8.28, and 55.39±8.88 years. The distribution of age, gender, education years, prevalence of classical cerebrovascular risk factors, and the results of MMSE were analyzed among three APOE genotype groups, and there was no statistically significant difference observed, except for the prevalence of hyperlipidemia (Table 1).
Characteristics of the Study Population
APOE, Apolipoprotein E; MMSE, Mini-Mental State Examination; WMHs, white matter hyperintensities; CMBs, cerebral microbleeds; PVS, perivascular spaces; BPF, the brain parenchymal fraction; GMV, whole-brain gray matter volume; WMV, whole-brain white matter volume. *p < 0.05.
Neuroimaging small vessel disease characteristics and brain volume in different APOE genotype groups
Neuroimaging small vessel disease characteristics and brain volumes of all participants and different subgroups divided by APOE genotype are shown in Table 1. No significant differences in neuroimaging small vessel disease characteristics (WMHs, CMBs, lacunes, and severe PVS), global measures (total brain volume, BPF, GMV, WMV, and cortical GMV), brainstem, hippocampus, and subcortical nuclei (caudate, pallidum, putamen, thalamus) were observed.
Interaction between APOE genotype and age
The interaction of APOE genotype by age on cerebral structures was analyzed. As Table 2 demonstrates, there was an interaction between APOE ɛ4(+) and age on hippocampus was observed (p = 0.045). No significant interaction of APOE genotype by age was found on other brain structures. The detailed results of each independent variable in this general linear model are provided in Supplementary Table 1.
The interaction of APOE genotype by age on cerebral structures
APOE, Apolipoprotein E; WMHs, white matter hyperintensities; BPF, the brain parenchymal fraction; GMV, whole-brain gray matter volume; WMV, whole-brain white matter volume. *p < 0.05.
Effect of APOE genotype on cerebral structures stratified by age
The detailed results of the effect of APOE genotype on different cerebral structures stratified by age are shown in Tables 3 and 4. Compared with APOE ɛ3/ɛ3, there was a significant protective effect of APOE ɛ4(+) on BPF (β = 0.450, p = 0.048) in subjects aged≤60 years; in participants aged > 60 years, APOE ɛ4(+) had a negative effect on hippocampal volume (β = –1.087, p = 0.021) (Table 3). No significant effect of APOE ɛ2(+) on cerebral structures was found (Table 4).
Effect of APOE ɛ4 (+) on cerebral structures stratified by age
APOE, Apolipoprotein E; WMHs, white matter hyperintensities; BPF, the brain parenchymal fraction; GMV, whole-brain gray matter volume; WMV, whole-brain white matter volume. *p < 0.05.
Effect of APOE ɛ2 (+) on cerebral structures stratified by age
APOE, Apolipoprotein E; WMHs, white matter hyperintensities; BPF, the brain parenchymal fraction; GMV, whole-brain gray matter volume; WMV, whole-brain white matter volume. *p < 0.05.
The comparison of demographic information, APOE genotype distribution and MRI measures between different age groups are provided in Supplementary Table 2, and the results of each independent variable in the general linear models are provided in Supplementary Tables 3 and 4.
DISCUSSION
To our knowledge, this is the first large-scale study focusing on the distribution of APOE genotypes and its age-related effect on cerebral volumetric structures in Chinese community-dwelling adults. Our study revealed that in subjects aged≤60 years, APOE ɛ4 had a potential protective effect on brain parenchyma fraction; and in participants aged > 60 years, APOE ɛ4 had a negative effect on hippocampal volume. Our findings suggest that APOE ɛ4 has differential influences on cerebral structures in different age stages, providing a novel perspective for exploring the pathogenic mechanism of APOE gene in further study.
In this study, participants carrying ɛ4 allele accounted for 14.2%, ɛ3 allele 71.2%, and ɛ2 allele 14.0%. The distribution of APOE alleles varies significantly across the world [23]. As previous epidemiologic studies showed, the frequency of APOE ɛ3 in Asia was about 85%, the frequency of APOE ɛ4 in China was 10–25%, and the frequency of APOE ɛ2 was 7–19% worldwide [23]. In the past research conducted in community-dwelling populations, the frequency of APOE ɛ4 varied significantly, about 17–55.5% [7, 25].
APOE plays an important role in lipid metabolism, neuroplasticity, synaptogenesis, membrane preservation, and blood-brain barrier maintenance [11], and is the major genetic risk factor for several age-related diseases, such as AD and cerebral amyloid angiopathy. In AD patients, APOE ɛ4 has been found to be related to hippocampal atrophy and cognition decline [26]. And as previous literature showed, dementia is the ultimate consequence of long-time progress of AD pathology, which could begin without any clinical symptoms. Therefore, whether APOE gene begins to modulate the brain structure before the onset of clinical symptoms should be taken into consideration. Up to now, a number of studies focusing on the effect of APOE genotype on brain structure in community-dwelling cohorts have shown that APOE ɛ4 is correlated negatively with brain volumes, especially in regions connected to AD pathogenesis, suggesting that APOE gene begins to exert its influence before the clinical manifestations [7–9].
However, the average age of the participants in the above-mentioned studies was relatively old (about 68.4–73.6 years), and in research conducted in younger cognitively unimpaired individuals or MCI patients whose mean age was mostly younger than 60 years, an interaction of APOE ɛ4 by age on cerebral structures was found. In 2008, Mueller et al. observed a significant effect of APOE ɛ4 on hippocampal zones in older cognitively intact populations (aged 61–85 years) but not in the younger (aged 28–60 years) [13]. In 2015, Chen et al. found there was a significant interaction of APOE genotype by age on grey matter volume in the left calcarine, left insula, and left medial frontal gyrus in MCI patients, suggesting that the interaction between APOE ɛ4 and age might accelerate the morphological and pathological transition from MCI to AD in APOE ɛ4 carriers [14]. In 2018, Cacciaglia et al. observed lower grey matter volume of the right hippocampus, caudate, precentral gyrus, and cerebellar crus in healthy APOE ɛ4 homozygotes after the age of 50 years [15]. In 2021, Martí-Juan et al. found specific regions on the hippocampal surface where hippocampal shape was modulated by significant linear and quadratic interactions of APOE ɛ4 by age [16]. In our study, we found that in subjects aged≤60 years, AOPE ɛ4 was a protective factor for brain parenchyma volumes, and in those aged > 60 years, APOE ɛ4 had a negative effect on hippocampal volume. These findings all suggest that APOE ɛ4 might have differential effects on brain structures in different age groups.
By now, one of the potential hypotheses of the interaction between APOE genotype and age is the genetic antagonistic pleiotropy (AP), which is the most widely accepted mechanism for the evolutionary origin of aging [27]. It presumes that if a gene can control more than one character (pleiotropy), the effects of these characters may be different or even conflicting at different age stages, and if one character is beneficial for growth and fertility, another may be detrimental in late life (antagonistic). There are some examples of AP in human beings. Some genes playing an indispensable role in early life period might be detrimental in late life, which could accelerate aging and even lead to self-destruction. For example, insulin/insulin-like growth factor 1 (IGF-1) signaling, which is essential for growth and development, could increase the risk of cancer and accelerate the aging process [28, 29]; inflammation is indispensable to defense against microbes and degrade damaged tissue, but it is also the widely recognized mechanism of aging and age-related diseases [30]. Other evidence of human genetic AP has been found in genes associated with coronary heart disease, cancer, cystic fibrosis, etc. [31].
APOE gene is also considered to be the evidence of AP [11]. Despite the notorious pathogenic effect of APOE ɛ4 and the protective impact of APOE ɛ2 on AD late in life, they might play differential roles in the early-life stage. Studies have found that neonates and infants with APOE ɛ4 showed better prognosis compared with those APOE ɛ2 carriers after a cardiac surgery [32]. And young APOE ɛ4 carriers performed better in specific cognitive tasks compared with non-APOE ɛ4 carriers [33, 34]. What’s more, it has been found that APOE ɛ4 might be associated with female reproductive advantages and could promote fertility, suggesting its evolutionary adaption [35].
Researchers have raised a model of AP of APOE. It is postulated that in the early life stage, APOE ɛ4 carriers achieve a similar or even better cognitive performance by recruiting task-related brain regions or additional regions beyond task-related areas (It is supported by the evidence that in cognitively intact adults, APOE ɛ4 carriers have higher activations in task-negative and task-positive areas [36]); with age increasing and the AD disease burden accumulating, the APOE ɛ4 carriers’ recruitment fails to compensate and the clinical symptoms begin [12]. Similarly, in our study, APOE ɛ4 carriers had a larger brain parenchyma volume in younger adults, which might result from the compensatory recruitment of more brain structures to support daily cognitive activities. However, as age increases, the disease burden sufficiently accrued, leading to the minimization of volume-preserving effects of APOE ɛ4 allele, and reversely, resulting in volume loss of hippocampus, which harbors sensitive neurons functioning in cognition.
There are some limitations in our study. First, although Shunyi Study is a large-scale cohort study, considering the inevitable familial aggregation, there might be some selection bias in APOE genotype distribution. Moreover, the effect of APOE is the complex consequence of racial, metabolic, and gene-environment interactions, and other potential confounding factors should be taken into consideration in further studies. Thirdly, considering the nature of cross-sectional study, causality needs to be further validated in the future prospective study.
In conclusion, our study is the first large-scale study in which the distribution of APOE genotypes and its age-related effect on cerebral structures in Chinese community-dwelling adults were explored. Our study shows that APOE ɛ4 has differential effects on cerebral structures in different lifespan. These results suggest the underlying complicating biological function of APOE ɛ4, supporting its antagonistic pleiotropy, and we hope it could lay a foundation for further studies focusing on the pathogenetic mechanism of APOE and its hereditary association with age-related diseases.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
This work was supported by Science Innovation 2030 — Brain Science and Brain-Inspired Intelligence Technology Major Project Task 5 (No. 2021ZD0201101) and Chinese Academy of Medical Sciences (CAMS) Innovation Fund for Medical Sciences (CIFMS) (No. 2021-I2M-C & T-B-004).
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
Jun Ni and Yicheng Zhu are Editorial Board Members of this journal but was not involved in the peer-review process nor had access to any information regarding its peer-review.
DATA AVAILABILITY
The datasets generated during and/or analyzed during this study are available from the corresponding author on reasonable request.