
Abstract
Background:
Cognitive reserve may protect against cognitive decline.
Objective:
This cross-sectional study investigated the association between cognitive reserve and physiological measures of cognitive workload in older adults with cognitive impairment.
Methods:
29 older adults with cognitive impairment (age: 75±6, 11 (38%) women, MoCA: 20±7) and 19 with normal cognition (age: 74±6; 11 (58%) women; MoCA: 28±2) completed a working memory test of increasing task demand (0-, 1-, 2-back). Cognitive workload was indexed using amplitude and latency of the P3 event-related potential (ERP) at electrode sites Fz, Cz, and Pz, and changes in pupillary size, converted to an index of cognitive activity (ICA). The Cognitive Reserve Index questionnaire (CRIq) evaluated Education, Work Activity, and Leisure Time as a proxy of cognitive reserve. Linear mixed models evaluated the main effects of cognitive status, CRIq, and the interaction effect of CRIq by cognitive status on ERP and ICA.
Results:
The interaction effect of CRIq total score by cognitive status on P3 ERP and ICA was not significant. However, higher CRIq total scores were associated with lower ICA (p = 0.03). The interaction effects of CRIq subscores showed that Work Activity affected P3 amplitude (p = 0.03) and ICA (p = 0.03) differently between older adults with and without cognitive impairments. Similarly, Education affected ICA (p = 0.02) differently between the two groups. No associations were observed between CRIq and P3 latency.
Conclusion:
Specific components of cognitive reserve affect cognitive workload and neural efficiency differently in older adults with and without cognitive impairments.
Keywords
INTRODUCTION
Cognitive reserve refers to the adaptability of the brain to cope with the effects of normal and pathological aging on cognitive functioning [1, 2]. Various factors, including advanced education, intellectually stimulating work activities, and a rich social life, influence cognitive reserve [1, 3]. Older adults with greater cognitive reserve experience less age-related cognitive decline and may have a reduced risk of dementia [4–7].
Cognitive reserve may also impact the cognitive workload needed to execute a task. Cognitive workload reflects the exerted mental and physical effort in response to the cognitive demands and time constraints of a task [8]. Cognitive workload increases linearly with task demand until the cognitive resources available to complete the task are depleted. When cognitive overload occurs, task performance will decrease [9]. Older age and age-related neurodegeneration may affect the availability of cognitive resources, resulting in increased cognitive workload to execute the task [9]. Such increased cognitive workload has been observed in older adults with normal cognition and cognitive impairments, even with equal performance on cognitive tasks [9]. The increased cognitive workload observed in both normal and pathological cognitive aging is likely to reflect either decreased neural efficiency or compensatory mechanisms to cope with the demands of the task [9].
Electro-encephalography (EEG) and pupillary recording are two relatively inexpensive and non-intrusive physiological measures with a temporal resolution sufficient to evaluate cognitive workload in real-time [9]. Event-related potentials (ERP) are very small voltages generated in pyramidal neurons of the cortex in response to specific events or stimuli recorded using EEG [10]. The P3 component is the third positive peak in the ERP waveform that occurs at about or slightly later than 300 ms after stimulus presentation [11]. The P3 is believed to be associated with neural efficiency and cognitive resource allocation during information processing, memory encoding, and updating of information [12, 13]. Working memory tasks are widely used in P3 ERP studies across the spectrum of cognitive impairments, as deterioration in working memory is one of the earliest cognitive dysfunctions observed in mild cognitive impairment (MCI) and is a reliable predictor of Alzheimer’s disease (AD) [14, 15]. Although P3 has been used in healthy populations to explain the neural substrates of cognitive reserve, very few studies have investigated the relationship between cognitive reserve and P3 in clinical populations [16]. While higher cognitive reserve correlated with P3 amplitude and latency in healthy older adults, no such relationship has been observed in MCI [17]. Changes in pupillary size have also been shown to correlate with cognitive workload in older adults with and without cognitive impairments [18–20], but no studies have evaluated the effect of cognitive reserve on pupillary response in people with cognitive impairments.
As cognitive reserve plays a crucial role in slowing down the progression of MCI and AD, the aim of this study was to compare the effect of cognitive reserve on physiological measures of cognitive workload between older adults with and without cognitive impairments. We hypothesized that higher cognitive reserve will be associated with lower cognitive workload, indexed by larger P3 amplitudes and decreased pupillary response. This relationship will manifest more profoundly in older adults with normal cognition compared to those with impaired cognition.
MATERIALS AND METHODS
Participants
Participants were recruited between 5/30/2018 and 02/25/2022 from the University of Kansas Alzheimer’s Disease Research Center (KU ADRC). Inclusion criteria were: 1) age 65 years or older; 2) understanding of all instructions in English; or 3) informed consent. Exclusion criteria were: 1) currently taking steroids, benzodiazepines, or neuroleptics; 2) history of any substance abuse; or 3) any contra-indications to EEG.
The Clinical Dementia Rating [21] and the Uniform Data Set 3.0 neuropsychological battery [22] were administered to determine cognitive status (normal or impaired). Cognitive status was determined in a consensus diagnostic conference. The MCI/AD diagnostic criteria were followed to identify participants with cognitive impairments [23, 24]. The team administering the cognitive tests, EEG, and pupillary recordings was blind to the cognitive status of participants. Participants with normal cognition had previously undergone a PET scan of the brain to rule out increased amyloid-β depositions. Three raters interpreted the PET scans to rule out elevated amyloid-β [25, 26]. The protocol has been published elsewhere [27].
This study was approved by the Human Subjects Committee at the University of Kansas Medical Center (#4461).
Participants with cognitive impairments had either mild or major neurocognitive disorder. The suspected clinical diagnosis was verified with medical records. Seventeen (59%) were categorized as mild neurocognitive disorder (MCI due to probable AD, n = 8; MCI of unknown or mixed etiology, n = 7; vascular MCI, n = 1; and MCI due to frontotemporal dementia, n = 1). Twelve (41%) were categorized as major neurocognitive disorder (AD, n = 6; dementia of unknown or mixed etiology, n = 4; and Lewy body dementia, n = 2). No participants were included with cognitive impairments due to traumatic or other causes.
Procedure
Demographic and clinical information
The demographic survey included age, sex, and hand dominance. All participants were right-handed. We screened for cognitive impairments using the Montreal Cognitive Assessment (MoCA) [28].
Cognitive Reserve Index questionnaire
The Cognitive Reserve Index questionnaire (CRIq) is a widely validated questionnaire of cognitive reserve [29]. The CRIq is composed of 20 items, which are categorized into three sections: Education, Working Activity, and Leisure Time. Total CRIq scores are calculated as the average of the section scores, each standardized and transposed to a mean = 100 and standard deviation of 15. Scores lower or equal to 70 are considered low; between 70 and 84 medium-low; between 85 and 114 medium; between 115 and 130 medium-high; and higher than 130 high.
N-back test
The n-back test involved participants pressing a mouse button with the right hand when a white letter that appeared on a black screen was the same as the letter presented n-places back [30, 31]. Participants completed the 0-, 1-, and 2-back test. The n-back tests reflect working memory, but higher levels of task demand also assess higher-order cognitive functions such as updating information and maintaining representations of recently presented stimuli [32].
Participants familiarized themselves with each test by practicing a random sequence of 3 targets and 7 nontargets until they felt comfortable to proceed with the actual test. Each n-back test consisted of 60 trials (target, 33.3%) that required a mouse click and 120 trials (nontarget, 66.7%) that did not require a response. This ratio was set in order to get >30 artifact-free trials for good-quality ERPs, assuming a low success rate on the 2-back test in individuals with cognitive impairments (Table 1). A lower probability of targets elicits a larger P3 [33] and creates a longer experiment, which might introduce fatigue effects in older adults. Therefore, the 1:3 ratio was chosen to get a trade-off between data quality and experiment time. The same ratio has been used in previous working memory studies using the 2-back test [34, 35]. All trials were presented in random order. Stimulus onset asynchrony was 2200 ms (presentation time of 500 ms and blank interstimulus interval of 1700 ms). We set a random jitter of +/-50 ms. The total duration of each n-back test was about 400 s. The main performance outcome measures were accuracy, defined as the number of correct responses to the 60 trials, and response time in seconds.
Comparison of clinical and behavioral variables between older adults with normal cognition (NC) and cognitive impairments (CI)
NC, normal cognition; CI, cognitive impairment; CRIq, Cognitive Reserve Index questionnaire; MoCA, Montreal Cognitive Assessment. Variables are described as mean±standard deviation; median (Q1 –Q3), or frequency (percentage). aIndependent t-test; bChi Square test; cWilcoxon Rank Sum test.
P3 event-related potential
While completing the n-back tests, participants were fitted with a high-density 256-electrode Geodesic Sensor Net from Magstim EGI. The Net Amps 400 amplifier was used with a bandwidth from DC to 2,000 Hz and input impedance larger than 1 GΩ. The electrode impedance was kept less than either 50 KΩ or 100 KΩ. Continuous EEG was digitized at 1,000 Hz and referenced to Cz without an online filter. All other EEG processes were done in EEGLab [36] and in ERPLab [37]. In the offline process, EEG data were band-pass filtered between 0.5 Hz and 30 Hz in Magstim EGI NetStation Tools software with a roll-off frequency of 0.3 Hz. Electrodes around the face were removed from data processing, leaving 183 channels to be analyzed. Bad channels were removed through visual inspection of data and automatic identification using joint probability methods with a z-score kurtosis cut-off of 6. Subsequently, independent component analysis was employed to detect and remove various ocular and movement artifacts or cardiovascular signals. Continuous EEG data were segmented into stimulus-locked ERPs with an epoch interval of 100 ms before to 1000 ms after stimulus onset. We used the baseline correction function rmbase() in EEGLAB toolbox to remove the mean of the baseline (–100 ms∼0 ms) from each epoch and channel. Epochs of incorrect and missed responses were manually removed. Signals from bad electrodes were interpolated using surrounding electrode data. All channels were off-line referenced to the linked mastoid average. ERP data containing more than 50% artifacts (which included noisy epochs and epochs of missed and incorrect responses) were not included in the analyses. Artifacts ranged from 0% to 26% (mean 9% ±7) of epochs for the 0-back test, 0% to 36% (mean 7% ±8) for the 1-back test, and 0% to 43% (mean 10% ±12) for the 2-back test. Thus, no participant was excluded from the ERP analyses. The measurement window of the P3 ERP was established a priori and ranged between 250 ms and 650 ms, based on our preliminary data [38]. We included the midline channels Fz, Cz, and Pz in our analyses. The P3 amplitude and latency at these electrode sites were calculated as the average of the clusters of surrounding channels [39].
Since the P3 difference waveform included both a negative and positive component (Fig. 1), we calculated the rectified area amplitude within the P3 time window of the task effect (target –nontarget). P3 latency was calculated as the 50% fractional area latency, which is the midpoint of the component that divides the area under the curve into two equal regions. Fractional area latency is typically recommended for difference waves of P3 components [40].
Index of cognitive activity
A remote infrared eye tracker (FX3, SeeingMachines, Inc, Canberra, Australia) was mounted right below the computer screen to record pupillary data of both eyes while completing the n-back tests. Pupillary size was recorded at 60 Hz using EyeWorks Record and analyzed using Eyeworks Analyze (Eye Tracking, Inc, Solana Beach, CA). We required participants to focus on the screen which reduces potential artifacts from the light reflex and eye movements on pupillary recording. However, the pupil continues to oscillate irregularly even when controlling for ambient lighting. The Index of Cognitive Activity (ICA) decomposes the raw pupillary size into different wavelets of high and low frequency components of the signal [41]. This allows changes in pupillary size in response to cognitive workload to be separated from changes in pupillary size due to the light reflex [8, 42]. The ICA is calculated by dividing the number of rapid small pupillary dilations per second by the number of expected rapid pupillary dilations per second [41]. The values are then transformed using the hyperbolic tangent function. Blinks are factored out by linear interpolation of adjacent time spans to produce continuous values ranging between 0 and 1 [41]. We calculated missing data for each person, each eye, and each n-back test. Missing data ranged from 0 to 55% (mean 3±8%) for the left eye and 0 to 96% (mean 4±14%) in the right eye. Since the left eye produced more accurate recordings, we only reported the mean ICA of the left eye in the results. One person had more than 50% missing data in the left eye and was therefore removed from the analysis.
Data analysis
Unpaired t-tests, Wilcoxon Rank Sum tests, and Chi-square tests were used to compare differences in descriptive and behavioral variables between groups. Linear mixed models were employed to identify the relationship between CRIq and physiological outcomes. We used a random intercept term with a subject-specific coefficient to adjust for correlation between measures within subjects. For P3 amplitude and latency, we entered one main between-group effect of cognitive status (normal cognition, cognitive impairment), and two main within-group effects of task demand (0-, 1-, 2-back) and electrode sites (Fz, Cz, Pz). For ICA, we used one main between-group effect (cognitive status) and one main within-group effect (task demand). We entered CRIq scores as covariate and CRIq by cognitive status as interaction effect into the model. Effect sizes (ES) of significant interaction effects were reported as the beta estimate divided by the standard error. We first ran the linear mixed model with main and interaction effects. We subsequently ran the model with only main effects. The total CRIq score was our main independent variable; however, we explored CRIq Education, Work Activity, and Leisure Time in separate linear mixed models as well. Each model was adjusted for age and sex. Residuals of all outcome variables were normally distributed. p values of 0.05 or less were considered significant. All analyses were performed in SAS, version 9.4.
RESULTS
Participant characteristics
We consented 29 older adults with cognitive impairment (age: 74±6, 11 (38%) women, MoCA 20±7) and 19 with normal cognition (age: 74±6; 11 (58%) women; MoCA 28±2). Participants with cognitive impairment scored lower on the MoCA and all behavioral outcomes, except for accuracy on the 0-back test (Table 1). No differences were found in CRIq total or subscores between groups. CRIq scores ranged from 81 to 158 in the normal cognition group: one participant (5%) scored low; 3 (16%) scored medium-low; 9 (47%) scored medium-high; and 6 (32%) scored high. Scores in the group with cognitive impairment ranged from 104 to 146: nine (32%) scored medium; 10 (34%) scored medium-high; and 10 (34%) scored high.
P3 event-related potential waveforms
The grand average waveforms of the targets and non-targets at electrode sites Fz, Cz, and Pz for each n-back test are shown in Supplementary Figure 1. The task effect, depicted as the difference wave and scalp maps of targets –nontargets of the P3, between the normal cognition and cognitive impairment groups at the three channels for each n-back test is shown in Fig. 1.
Outcomes of the physiological variables are described in Table 2. The results of the statistical analyses are reported below.
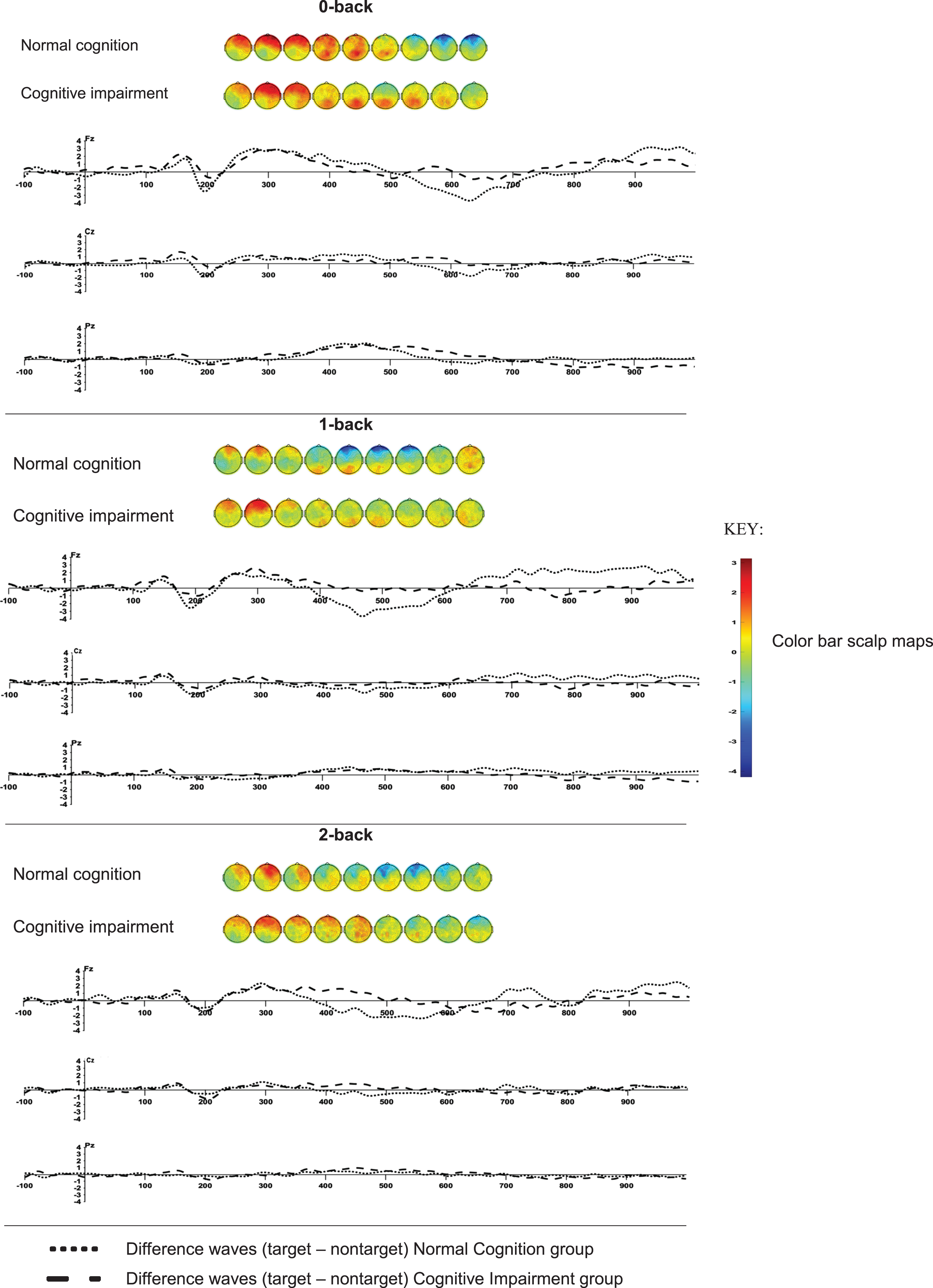
Scalp maps (from 250 to 650 ms in 50 ms increments) and difference waves (target –nontarget) of the P3 event-related potential of the 0-back, 1-back, and 2-back tests in the groups with normal cognition (n = 19) and cognitive impairments (n = 29).
P3 amplitude
The interaction effect of CRIq total score by cognitive status was not significant (Supplementary Table 1). Likewise, no main effect of CRIq total score on P3 amplitude was found (Supplementary Table 2). Task demand (F = 4.46; p = 0.01) and electrode site (F = 16.15; p < 0.0001) correlated with P3 amplitude (Fig. 1, Supplementary Table 2). Using Fz as a reference, P3 amplitude was higher at Cz (t = 2.42; p = 0.02) and lower at Pz (t = –3.24; p = 0.001). P3 amplitude was higher in the 2-back test compared to the 0-back test (t = 2.24; p = 0.03).
CRIq subscores were also entered in separate linear mixed models. A significant interaction effect (F = 4.98; p = 0.03) was found for CRIq Work Activity by cognitive status (Supplementary Table 1). Participants with normal cognition showed a positive association between CRIq Work Activity and P3 amplitude (ES = 2.50; t = 3.04; p = 0.003) compared to the null effect, whereas no difference was found between the slope of the regression line and the null effect in the cognitive impairment group (ES = 0.67; t = 0.69; p = 0.49).
CRIq Work Activity also showed a significant main effect with P3 amplitude (F = 4.12; p = 0.004), with higher scores reflecting increased P3 amplitude (Supplementary Table 2).
Education and Leisure Time did not affect P3 amplitude.
Comparison of physiological variables between older adults with normal cognition and cognitive impairments for each n-back test
NC, normal cognition; CI, cognitive impairment; Variables are described as mean±standard deviation.
P3 latency
CRIq total and subscores did not correlate with P3 latency (Supplementary Tables 1 and 2). Only task demand (F = 4.46; p = 0.01) and electrode site (F = 16.14 p < 0.0001) were significantly associated with P3 latency (Supplementary Table 2). P3 latency was significantly shorter at Cz (t = –2.62; p = 0.009) compared to Fz. P3 latency was also longer in the 2-back task (t = 2.96; p = 0.003) compared to the 0-back test.
Index of cognitive activity
Whereas no significant interaction effect of CRIq total score by cognitive status was found on ICA (Supplementary Table 1), the main effect of CRIq total score (F = 3.04; p = 0.03) was significant (Supplementary Table 2). Higher CRIq total scores were associated with lower ICA (t = –2.14; p = 0.03). In addition, task demand affected the ICA (F = 7.59; p = 0.009). ICA was lower in the 2-back compared to 0-back (t = –3.56; p = 0.0006).
The associations between the CRIq subscores and ICA were also explored in separate linear mixed models. The interaction effect of CRIq Education by cognitive status (F = 5.62; p = 0.02) was significant (Supplementary Table 1). Higher CRIq Education scores correlated with lower ICA in older adults with normal cognition (ES = –1.5; t = –1.62; p = 0.11), whereas a positive association was found in older adults with impaired cognition (ES = 1.5; t = 1.70; p = 0.09). Likewise, CRIq Work Activity affected ICA differently in older adults with normal cognition and those with cognitive impairment (F = 5.69; p = 0.02). Larger scores on the Work Activity subscale were associated with decreased ICA, but only in adults with normal cognition (ES = –2.5; t = –3.15; p = 0.002) and not in those with impaired cognition (ES = –0.78; t = –0.73; p = 0.47).
Leisure Time did not affect ICA.
DISCUSSION
The aim of this study was to evaluate the association between cognitive reserve and physiological measures of cognitive workload in older adults with and without cognitive impairment. In particular, Work Activity of the CRiq affected P3 ERP and ICA differently between older adults with normal cognition and those with cognitive impairment.
Total scores on the CRIq did not impact physiological measures of cognitive workload differently between older adults with and without cognitive impairment. In contrast, Gu et al. found distinct mediation processes of cognitive reserve and P3 ERP in older adults without and with cognitive impairments [17]. Whereas higher cognitive reserve affected the efficiency of neural processing in normal cognitive aging, no such effects were found in patients with amnestic MCI [17]. Differences in calculation of the composite measure of cognitive reserve may have accounted for the discrepancies in results. Whereas our study used the total score of the CRIq, Gu et al. used a composite score of cognitive reserve derived from CRIq and the IQ test of the Wechsler Adult Intelligence Scale [17].
However, in support of Gu et al. [17], Work Activity and Education subscores of cognitive reserve correlated differently with physiological measures of cognitive workload in older adults with and without cognitive impairment. Older adults with normal cognition who experienced an intellectually stimulating occupation exhibited increased P3 amplitude, reflecting more efficient post-stimulus categorization processing [43]. Similarly, Education and Work Activity differently mediated pupillary response, indexed by ICA, in older adults with and without cognitive impairment. Higher education and vocational experiences were associated with decreased cognitive workload in older adults with normal cognition. Previous studies also highlighted education and work as important cognitively stimulating life events that affect physiological measures of cognitive workload [44]. Education had a moderating effect on the relationship between hypometabolism and cognition, whereas previous work activities had a moderating effect on the relationship between cortical atrophy and cognition in AD [44]. These findings suggest that lifestyle choices in early- and midlife may have long-lasting implications on physiological processes of cognitive workload.
Pupillary response showed to be a sensitive measure of cognitive workload that is influenced by cognitive reserve. Participants with higher cognitive reserve exhibited lower cognitive workload to complete the working memory tasks. ICA is a measure of cognitive workload based on moment-to-moment changes in pupillary size [41]. Pupillary dilation in response to cognitive workload is mainly mediated through the locus coeruleus [45]. The locus coeruleus is the main supplier of noradrenaline in the brain and critical in the regulation of physiological arousal and cognition [46]. In vivo and postmortem imaging studies demonstrate that older adults with cognitive impairments exhibit decreased neuronal density and early tau accumulation in the locus coeruleus [47]. These neurobiological abnormalities in the locus coeruleus result in increased pupillary dilation during cognitive tasks, even in the absence of impairments in cognitive performance [20, 48]. The locus coeruleus noradrenergic system has also been implicated as a potential source generator of the P3 ERP [49, 50]. Although no studies have found evidence of tight coupling between stimulus-locked pupillary size and P3 [51, 52], there is a link between pre-stimulus pupillary size and subsequent P3 response [50, 52]. These results suggest that both pupillary response and P3 may reflect independent, but complementary processes of task-relevant decision-making. The relationship between cognitive reserve and noradrenergic-regulated physiological measures provides further support of the noradrenergic theory of cognitive reserve, which postulates that continuous upregulation of noradrenaline in the brain throughout the lifespan is paramount to building cognitive reserve [53]. Individuals who continuously engage in cognitively stimulating activities in their lifetime may exhibit an overall higher noradrenergic tone, building more resilient neurobiological networks that protect against age-related neurodegeneration. Our results support previous studies identifying the role of the locus coeruleus noradrenergic system as a key mediator of cognitive reserve [54].
Although no support is found for the effect of cognitive reserve on neural efficiency in older adults with cognitive impairment using EEG, other studies using functional MRI revealed protective mechanisms of cognitive reserve across the spectrum of cognitive aging [55]. Higher cognitive reserve was related to increased frontal activity in older adults with cognitive impairments compared to those with normal cognition [56]. However, the time window for modulation of cognitive reserve on brain reserve may be limited to individuals with MCI [57]. A previous study found that higher cognitive reserve in older adults with amnestic MCI was associated with lower macromolecular tissue volume across major white tracts, whereas no such relationship was observed in individuals with AD [58]. Likewise, older adults with MCI and high cognitive reserve manifested decreased cortical thickness in the right temporal and left prefrontal lobes, along with an increased fractional dimension in the right temporal and in the left temporo-parietal lobes compared to those with lower reserve [57]. Future studies combining the spatial accuracy of MRI and the temporal dynamics of EEG may elucidate the mediating effects of cognitive reserve between localized brain compensatory activity and neural efficiency across the cognitive aging spectrum.
Our study has several limitations. First, our study included participants with different etiologies and degrees of cognitive impairment, making it difficult to pinpoint the neurobiological mechanisms underlying the link between cognitive reserve and physiological measures of cognitive workload. Therefore, the study results cannot be interpreted according to a specific dementia type. However, our results also demonstrate a strong association between cognitive reserve and physiological measures of cognitive workload, regardless of disease pathology. Our methods could therefore be applied to different types of dementia. Second, our participants were recruited from a volunteer registry database. This selection bias shows in the relatively high average CRIq scores. Caution is therefore warranted extrapolating our results to the general population of older adults with and without cognitive impairments. Third, the cross-sectional design of our study prevents making any inferences about the causal relationships between cognitive reserve and physiological measures of cognitive workload. Finally, our results need to be validated in cognitive domains other than working memory.
Conclusions
Cognitive reserve directly affects physiological measures of cognitive workload across the cognitive aging spectrum. Higher cognitive reserve is associated with more efficient neural processing and decreased cognitive workload to accomplish working memory tasks. However, this relationship is only observed in older adults with normal cognition and in specific components of cognitive reserve. Advanced education and occupational attainment are modifiable lifestyle choices that may directly benefit cognitive reserve. Future longitudinal studies should investigate the causal relationship between cognitive reserve and physiological processes of neural efficiency in normal and pathological cognitive aging.
Footnotes
ACKNOWLEDGMENTS
This work was completed at the Hoglund Biomedical Imaging Center which is supported by the Forrest and Sally Hoglund and a High-End Instrumentation grant from the National Institutes of Health (S10 RR29577). The authors thank all participants for volunteering their time.
FUNDING
Research reported in this publication was supported by the National Institute on Aging of the National Institutes of Health under Award Number K01 AG058785 and P30 AG072973. This study was supported in part by a pilot grant of the KU Alzheimer Disease Research Center (P30 AG035982). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.