
Abstract
Background:
Sarm1 (Sterile alpha and TIR motif-containing 1) is a key protein that regulates neurodegenerative pathologies. Alzheimer’s disease (AD) is highly associated with neurodegenerative lesions and biorhythmic disturbances.
Objective:
This study aims to decipher the role of Sarm1 in AD-induced circadian rhythm disturbances and AD progression.
Methods:
Open field and water maze tests were used to assess the cognitive function of mice. Thioflavin-S staining was used to assess amyloid-β (Aβ) plaque deposition in the hippocampus and cortex. Rhythmic waveform of home cage activity and temperature was recorded to evaluate circadian rhythm. Expression of clock molecules including Bmal1 and Per2 in the hippocampus were analyzed using western blot and real-time PCR. Further, HT22 cells with Sam1 knockout were treated with Aβ31–35 treatment to initiate circadian rhythm disorder in the cellular level to assess the changes in Bmal1 and Per2.
Results:
Our data suggested that Sarm1 deficiency rescued cognitive disorder, decreased Aβ plaque deposition in the hippocampus and cortex, inhibited astrocyte activation, improved circadian rhythm, altered clock molecule expression in the cortex and hippocampus in APP/PS1 mice.
Conclusion:
Sarm1 attenuates circadian rhythm disturbances and reduces AD progression. These data support the potential use of Sarm1 as a therapeutic target to improve circadian rhythm to impede AD progression.
INTRODUCTION
Circadian rhythm disturbances exemplified by sleep disorders are highly associated with a plethora of health impairments in the elderly [1, 2], including hypertension and cardiac insufficiency [3], respiration symptoms [4], physical disabilities [5], and decreased immune functions [6]. Recent studies have underlined circadian rhythm disturbances as significant contributors to poor cognitive and psychological functioning and depression [7]. As a result, the daily self-care ability of the elderly can be severely compromised, causing a mounting healthcare cost [8]. Particularly for patients with Alzheimer’s disease (AD), circadian rhythm disturbances are common, which is strongly linked to the functional disabilities and cognitive deterioration in AD patients [9, 10]. Over the years, substantial efforts have been devoted to the improvement of circadian rhythm disorders to delay the progression of AD [11, 12], which makes understanding major genetic factors associated with circadian rhythm disturbance imperative.
Sterile alpha and TIR motif containing 1 (Sarm1), also known as Myd88-5, is a protein of 724 amino acid in length that has recently emerged as a novel regulator of neuronal morphogenesis [13] and neuroinflammation [14], both of which are essential in the progression of neurodevelopmental disorders. Studies have indicated that Sarm1 regulates cognitive behaviors [15]. Transgenic mice with Sarm1 knockdown showed an association between Sarm1 expression and cognitive flexibility [16], social behaviors and associative learning. It has also been revealed that Sarm1 is an appealing axonal-specific therapeutic target [17]. While being kept normally in an inactive state by the association of the N-terminal armadillo domain with the C-terminal TIR domain, Sarm1 can be activated by a pathologically triggered intramolecular rearrangement during axonal degeneration, which leads to dimerization of the TIR domains [18]. Sarm1 upregulates in response to disease or injury, resulting in the NAD + depletion by catalyzing NAD + hydrolysis and the distal axonal segment degeneration. It is worth noting that Sarm1 knockdown has been shown to prevent neural degeneration and demonstrate therapeutic efficacies against neuropsychiatric diseases like autism [19] and brain traumatic injury [20], suggesting that Sarm1 inhibitors will likely be efficacious in treating other neurological diseases suchas AD.
Herein, we set out to evaluate the role of Sarm1 in the progression of AD to unveil the potential of Sarm1 inhibition as a therapeutic strategy in AD. We constructed Sarm1 knockout mice and performed cognitive and behavioral tests of the mice and analyzed clock molecules including Bmal1 and Per2 in hippocampus and HT116 cells with circadian rhythm disruptions.
MATERIALS AND METHODS
Animals
All animal experiments were approved by the People’s Hospital of Xinjiang Uygur Autonomous Region. This study was performed in strict accordance with the NIH guidelines for the care and use of laboratory animals (8th edition, NIH). APP/PS1 (APPswe, PSEN1dE9) (#034832) and Sarm1–/– (#018069) mice were purchased from Jackson Lab (Bar Harbor, ME) and bred with APP/PS1 het mice and Sarm1–/– mice to obtain APP/PS1 het and Sarm1–/+ mice. Then mice were self-bred with APP/PS1 het and Sarm1–/+ mice to obtain APP/PS1 het Sarm1+/+ (AD-WT), APP/PS1 het Sarm1–/– (AD-KO), and APP/PS1 neg Sarm1+/+ (Control) mice. Male mice at 16 months of age were used in behavioral tests, rhythm-related experiments, and tissue collection.
Open field test and water maze experiment
Open field test and water maze experiment were used to assess the locomotor and cognitive activity of mice. The distance mice moved was recorded and the recorded path into bouts of walking (> 8.5 cm/s) and episodes of lingering or resting was segmented to determine the percent of time walking and corresponding walking speed. Level of anxiety was correlated with the time spent in the center 50% of area and mean distance to center. All measurements from different days were averaged to acquire a single value per mouse.
Long-term spatial memory and learning were tested using Morris water maze (MWM) experiment [21]. Briefly, opaque water stained with tempera paint (25±0.5°C) was used to fill in a circular pool of 120 cm in diameter with spatial cues. A platform with a 9 cm in diameter was placed at 1 cm from water surface in the northwest quadrant of the pool. Animals were trained for four days in MWM prior to test. Each test lasted one minute, or once the animal arrived on the platform for 3 s. After tests, animals were dried with a towel and placed into a cage. Distance traveled, time to find the platform, and average speed were calculated based on recordedvideos.
Aβ assay (Thioflavin-S immunofluorescence)
Coronal sections were washed and treated with 0.3% Triton X-100 for 30 min. Then blocking buffer (5% normal goat serum in phosphate-buffered saline (PBS)) was incubated with the sections for 1 h. Tissue sections were then stained with 0.05% thioflavin-S in PBS or 8 min, followed by washing with 50% ethanol. Images of the slides were acquired using a laser-scanning confocal microscopy system (Nikon Instruments, Melville, NY, USA).
Home cage activity and body temperature measurement
Home cage activity and body temperature, which are two of the metrics of circadian rhythm, were monitored continuously with the Activity Monitoring System after mice were inserted with G2 E-mitter probes (Mini Mitter, Bend, OR) was subcutaneously on the dorsum of the neck. Data were recorded every 6 min. Daily actogram of 28 days was summarized in an Excel file and summed (in measuring home cage activity) or averaged (in measuring body temperature) as 1 h bins.
RT-qPCR
Total RNA was isolated from the harvested hippocampus by TRIzol reagent (ThermoFisher Scientific, Waltham, MA, USA) followed by reverse transcription using the Prime-Script RT reagent kit (ThermoFisher Scientific). Real-time PCR was completed using the SYBR Green qPCR Master Mix (ThermoFisher Scientific) with the following primers acquired from GenePharm (Shanghai, China) with the following sequences: Gapdh, forward, 5′-ACAG CCGCATC TTCTTGTGCAGTG-3′ and reverse, 5′-GGCCTTG ACTGTGCCGTTGAATTT-3′. Per2, forward, 5′-TGTGCGATGATGATTCGTGA-3′ and reverse, 5′-GGTGAAGGTACGTTT GGTTTGC-3′ Bmal1, forward, 5′-CCACCTCAGA GC CATTGATACA-3′ and reverse, 5′-GAGCAGGTT TAGTTCCACTTTGTCT-3′. The reactions were run in Applied Biosystems 7500 (Applied Biosystems, US) with triplicated wells. The expression level was determined by the ΔCt (ΔCt = Ct of target gene – Ct of Gapdh), and the Ct (cycle threshold) was defined as cycles number required for the fluorescent signal to cross the threshold that was determined manually. The expression level of target gene arose from the formula = 2–ΔCt. Then the relative expression level of a target gene in each sample was normalized to the mean value of the expression level in control group.
Western blot
Western blot was used to quantify protein expression levels in the mouse hippocampus or cells cultured in vitro. Briefly, tissue and cells were homogenized in lysis buffer and the supernatant was resolved by electrophoresis and transferred onto polyvinylidene fluoride membranes. After being blocked in 2% milk, membranes were incubated with primary antibodies overnight at 4°C, followed by chemiluminescence after incubation with corresponding secondary antibodies. Primary antibodies used in this study include: Gfap (47084, 1 : 3000; Cell Signaling Technology, Danvers, MA); human APP (MABN10, 1 : 2000; Millipore, Billerica, MA); CTFs (A8717, 1 : 1000; Sigma); Bmal1 (ab235577, 1 : 1000; Abcam, Cambridge, MA); Per2 (PA5-100107, 1 : 2000; Invitrogen, Waltham, MA); and Gapdh (#2118, 1 : 5000; Cell Signaling Technology). And the gray values for the bands were analyzed by Image J. The target protein level was normalized to corresponding gray value of Gapdh, then the relative expression level of a target gene in each sample was normalized to the mean value of the expression level in control group.
Cell culture and shRNA knockdown
HT22 mouse hippocampal cells were cultured in DMEM medium with 10% fetal bovine serum (Gibco, Grand Island, NY) penicillin and streptomycin (Sigma) in a 5% CO2 incubator at 37°C. The pLKO.1 expression vectors for shRNA knockdown were shNC (Sigma, SHC016) and shSarm1 (Sigma, TRCN0000175300), and transfected using Lipofectaminetrademark LTX (Invitrogen, Carlsbad, CA). Cells were treated with Aβ31–35 to initiate circadian rhythm disruption.
Statistical analysis
The significant difference was analyzed using two-tailed t-test, two-way or one-way ANOVA with Tukey’s post-hoc test in GraphPad prism 7 as described in the figure legends. Error bar represents mean±standard deviation (SD). The statistical significance was determined when the p values was less than 0.05.
RESULTS
Sarm1 deficiency rescued cognitive disorder in APP/PS1 mice
To evaluate the effects of Sarm1 knockdown on locomotive and cognitive capabilities of AD mice, open field and water maze analyses were performed on mice with or without Sarm1 knockdown. Our data shows that sarm1 deficiency significantly alleviated cognitive impairment in AD mice, as revealed by the higher amount of time spent in center (Fig. 1A), times of central entries (Fig. 1B) and percentage of central/total time (Fig. 1C) compared to wild-type AD mice in the open field test. However, similar speed of movement was observed in the control, wild-type and knockout groups, suggesting that locomotive ability was not significantly affected by AD and Sarm1 knockdown (Fig. 1D). Similarly, in the water maze experiment (Fig. 1E-I), with representative swimming trajectory shown in Fig. 1E, our data showed that escape latency (Fig. 1F) was higher in AD mice with Sarm1 knockdown compared to wild-type AD mice. Consistently, AD mice with Sarm1 knockdown also demonstrated higher time spend in the target quadrants (Fig. 1G) and times of crossing the platform (Fig. 1H), suggesting lowered cognitive impairment. The speed of mice in water maze experiment was also not significantly different among the three groups, confirming that AD and Sarm1 knockdown did not prominently affect locomotive ability of mice.
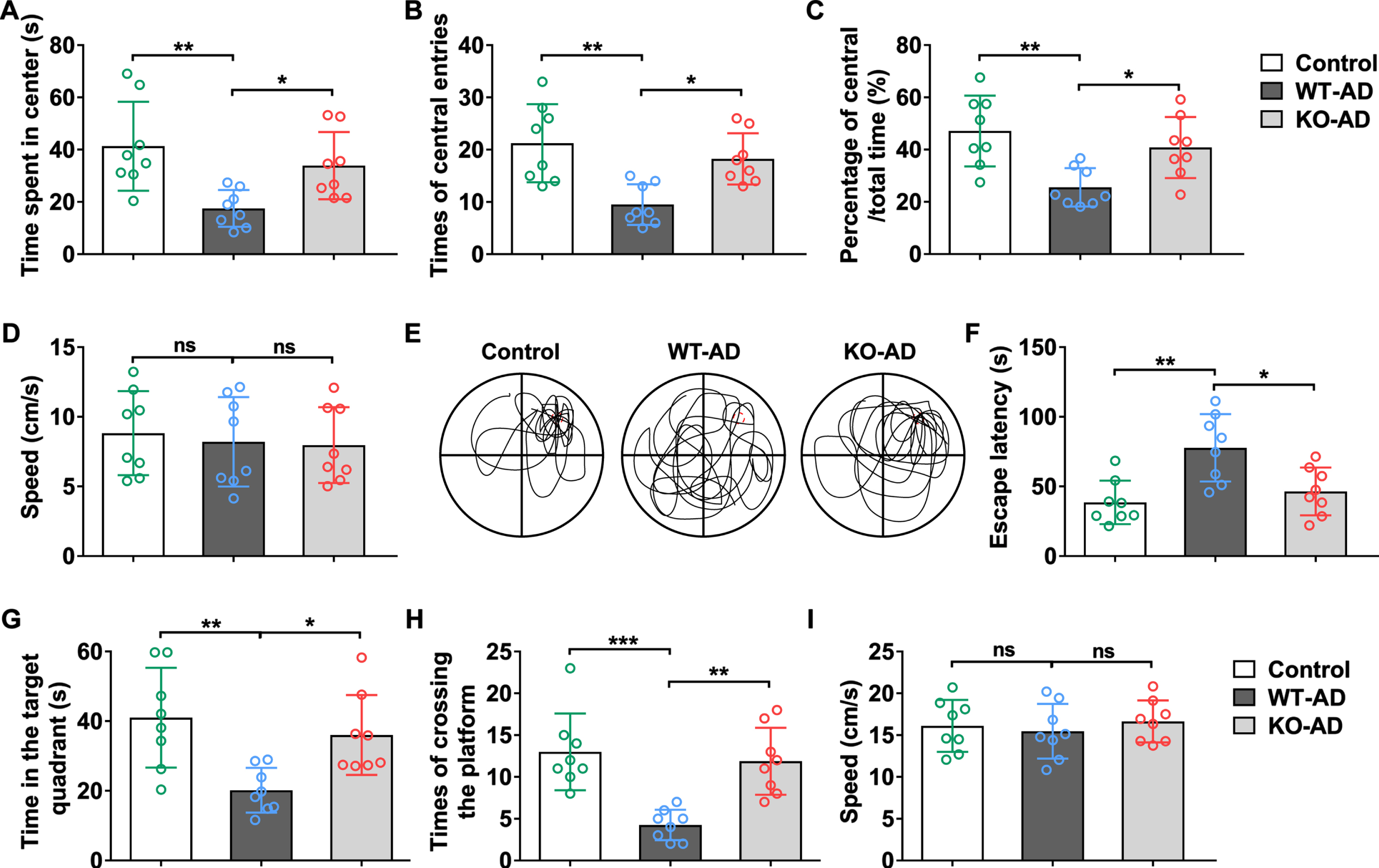
Sarm1 deficiency rescued cognitive disorder in APP/PS1 mice. A-D) Time spent in center (A), times of central entries (B), percentage of central/total time (C), and average speed (D) in open-field test. E-I) Representative swimming trajectories (E), escape latency (F), time in the target quadrant (G), and times of crossing the platform (H) and average speed (I) in Morris water maze test. 16 months old male mice were used in these tests. n = 8. The significance was analyzed by one-way ANOVA. Error bar represents mean±SD. *p < 0.05, **p < 0.01, ***p < 0.001. ns, not significant.
Sarm1 deficiency decreased Aβ plaque deposition in cortex and hippocampus of APP/PS1 mice
Quantification of plaque number, plaque area, and average plaque size in cortex (Fig. 2A–C) and hippocampus (Fig. 2D–F) tissues based on the Thioflavin-S staining suggested > 50% reduction in plaque number and area and > 20% reduction in average size in the Sarm1 knockdown mice. To demonstrate Aβ plaque reduction in the molecular level, we analyzed the expression of the human amyloid precursor protein (hAPP) and c-terminal fragments (CTFs) in the cortex (Fig. 2G–I) and hippocampus (Fig. 2J–L), showing reduced protein level of hAPP and CTFs in mice with Sarm1 deficiency. Taken together, these data suggested that Sarm1 knockdown alleviated AD symptoms.

Sarm1 deficiency decreased Aβ plaque deposition in cortex and hippocampus of APP/PS1 mice. Plaque number (A, D), plaque area (B, E), and average plaque size (C, F) in cortex and hippocampus of each group at 16-month-old male mice. n = 8. The significance was analyzed by one-way ANOVA. The protein level of human APP and C-terminal fragments (CTFs) in cortex (G-I) and hippocampus (J-L) of 16-month-old APP/PS1 mice. n = 4. The significance was analyzed by two-tailed t-test. Error bar represents mean±SD. **p < 0.01, ***p < 0.001. nd, not detected.
Sarm1 deficiency decreased astrocyte activation in cortex and hippocampus of APP/PS1 mice
We also analyzed astrocyte activation by immunoblotting on Gfap in cortex and hippocampus samples, showing significant increase in AD mice while sarm1 deficiency decreased Gfap level, which revealed the reduction of astrocyte activation in sarm1 knock out mice (Fig. 3A, B).

Sarm1 deficiency decreased astrocyte activation in cortex and hippocampus of APP/PS1 mice. The protein level of astrocytes marker Gfap in cortex (A) and hippocampus (B) of 16-month-old APP/PS1 mice. n = 4. The significance was analyzed by one-way ANOVA. Error bar represents mean±SD. *p < 0.05, ***p < 0.001.
Sarm1 knockout improves circadian rhythm disorder of APP/PS1 mice
Time-course changes in home cage activity (Fig. 4A, B) and body temperature (Fig. 4C, D) were documented to study the circadian rhythm of APP/PS1 mice. Our data revealed that rhythm disturbances in home cage activity and body temperature during the light/dark (LD) and dark/dark (DD) cycles were alleviated in AD mice with Sarm1knockdown.
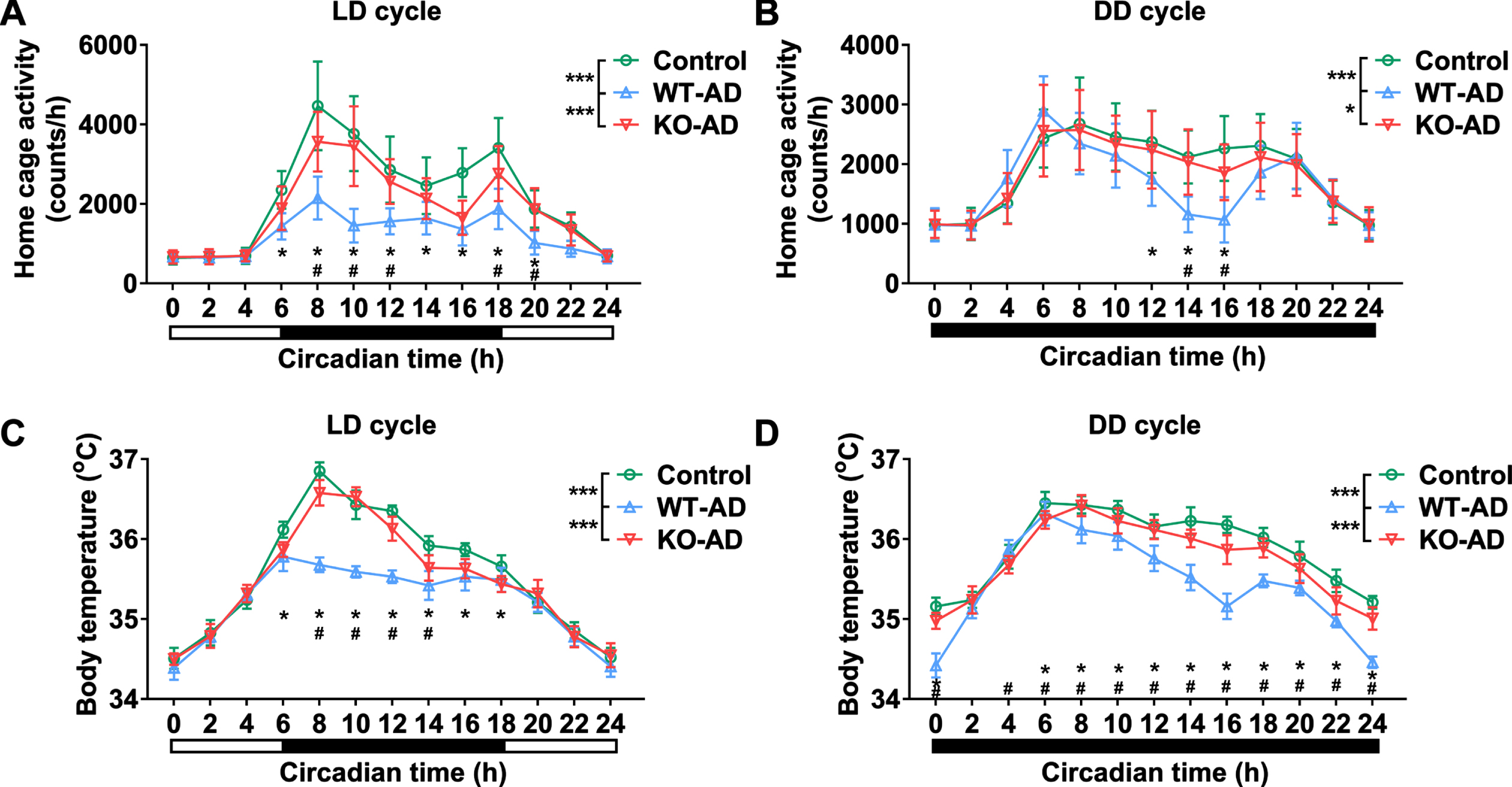
Sarm1 knockout improves circadian rhythm disorder of APP/PS1 mice. Rhythmic waveform of home cage activity (A, B) and body temperature (C, D) in APP/PS1 male mice of 16-month-old in light/dark (LD) (A, C) and dark/dark (DD) (B, D) cycle. n = 8. The significance was analyzed by two-way ANOVA. Error bar represents mean±SD. *p < 0.05, represents the significance of WT-AD group versus Control group. #p < 0.05, represents the significance of WT-AD group versus KO-AD group.
Sarm1 deficiency altered clock molecule expression in the hippocampus of APP/PS1 mice
We next examined the expression of clock molecules, including Bmal1 and Per2 in mRNA (Fig. 5A, B) and protein levels (Fig. 5C–E) in mice at different circadian time. Expectedly, while wild-type AD mice demonstrated significant deviation from the normal control mice in Bmal1 and Per2 levels, Sarm1 knockdown rendered the expression levels of clock molecules similar to normal control mice. This data corroborated that Sarm1 regulates circadian rhythm at the molecular level in AD mice.

Sarm1 deficiency altered clock molecule expression in the hippocampus of APP/PS1 mice. A, B) The mRNA levels of Bmal1 (A) and Per2 (B) in the hippocampus of 16-month-old APP/PS1 male mice at different circadian time. C-E) The protein levels of Bmal1 (C, E) and Per2 (D, E) in the hippocampus of 16-month-old APP/PS1 male mice at different circadian time. E) Representative results of Bmal1 and Per2 protein level at CT20 in hippocampus revealed by western blotting. n = 4. The significance was analyzed by two-way ANOVA. Error bar represents mean±SD. *p < 0.05, **p < 0.01, ***p < 0.001, represent the significance of WT-AD group versus Control group. #p < 0.05, # #p < 0.01, # # #p < 0.001, represent the significance of WT-AD group versus KO-AD group.
Sarm1 deficiency altered clock molecule expression in vitro
Given the regulation of clock molecules in vivo by Sarm1, we proceeded to check whether Sarm1 knockdown using shRNAs could exert similar effects in HT22 cells in vitro. Similar to observations in vivo, our data showed that Bmal1 and Per2 mRNAs (Fig. 6A, B) and proteins (Fig. 6C-E) were indeed affected by Sarm1 knockdown different circadian time.
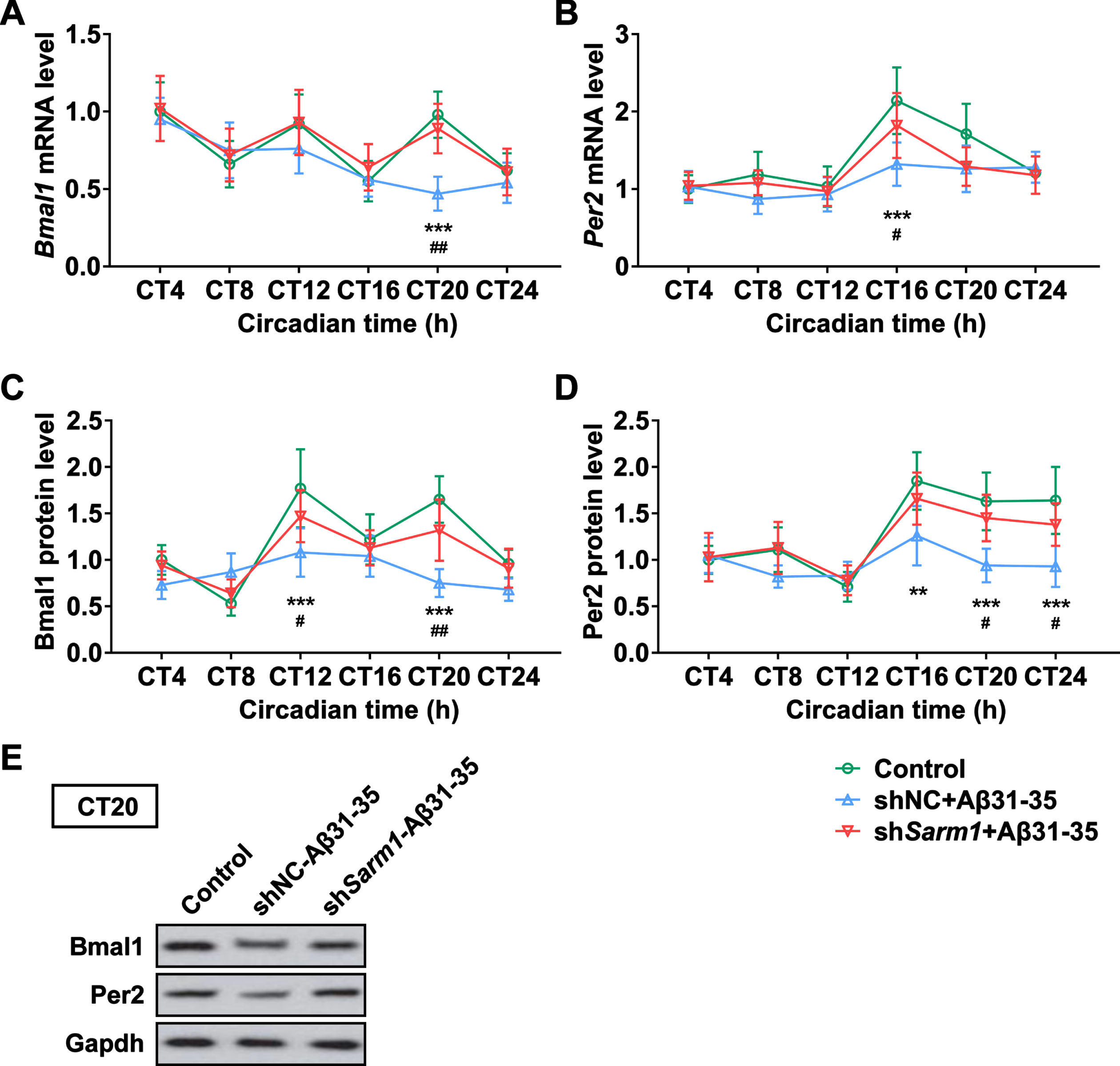
Sarm1 deficiency altered clock molecule expression in vitro. A, B) The mRNA levels of Bmal1 (A) and Per2 (B) in HT22 cells treated with Aβ31– 35 (5μM) at different circadian time. C-E) Western blot analysis of the Bmal1 (C, E) and Per2 (D, E) expression in the protein level in HT22 cells treated with Aβ31– 35 (5μM) at different circadian time revealed by western blotting. E) Representative results of Bmal1 and Per2 protein level at CT20. n = 4. The significance was analyzed by two-way ANOVA. Error bar represents mean±SD. **p < 0.01, ***p < 0.001, represent the significance of WT-AD group versus Control group. #p < 0.05, # #p < 0.01, represent the significance of WT-AD group versus KO-AD group.
DISCUSSION
Biological rhythms are evolutionary adaptations by organisms to the earth’s rotation [22]. Circadian rhythm disruption is a common consequence of aging and is even more pronounced in AD [9, 23]. The brain harbors the hypothalamic suprachiasmatic nucleus, which is the central biological clock that synchronizes physiological, biochemical, and behavioral rhythms [24]. Therefore, protection of neural degeneration is thought of as the key to preventing AD progression. Here we report that Sarm1 knockdown is efficacious in inhibiting AD progression by reducing circadian rhythm disturbance. This was demonstrated by the fact that AD mice with Sarm1 knockdown exhibited attenuated symptoms of circadian rhythm disturbance in behavioral and cognitive experiments. This translated to a reduction of Aβ deposition [25], indicating that Sarm1 knockdown impeded AD progression. Sarm1 is known to interact with signaling pathways involved in a large body of cellular functions, among which is the regulation of neural degeneration. Neurons in the brain preferentially expresses Sarm1, and Sarm1 deficiency was found to induce a reduction in TNF-α expression in the brainstem after being infected with the West Nile virus [26], suggesting a role of Sarm1 in neural immune response. Even in absence of viral infection, Sarm1 knockdown in mice induces alteration in inflammatory cytokine levels in the brain [27]. In vitro, Sarm1 also modulates neuronal morphogenesis, showing the ability to affect neuronal polarity, axonal outgrowth and dendritic arborization [28]. Our results confirms that Sarm1 is an important regulatory molecule in brain functions, and Sarm1 knockdown could serve as a therapeutic strategy with high potential in AD. In light of recent development of Sarm1 inhibitors [29, 30], we here propose that AD patients would likely benefit from those inhibitors through the restoration of circadian rhythm.
We also showed that Sarm1 knockdown regulates circadian rhythm in molecular levels, evidenced by the induced changes in expression of clock molecule Bmal1 and Per2 [31], which has not been reported before. However, the link between Sarm1 and the clock molecules remains to be established. Previous studies have indicated that Sarm1 affects TLR1 [32, 33] and MAPK [34] signaling pathways, which are important molecular mechanism that regulates neural inflammation and morphology [35]. Studies have shown that TLR1 and MAPK signaling pathways are important regulators of clock molecules [32, 36], and we hypothesize that the ameliorating effects of Sarm1 knockdown in AD mice stemmed from the regulation of these pathways. In addition, inflammation has been shown to be highly associated with circadian rhythm [37] and the role of Sarm1 in neural inflammation could play a significant part. Furthermore, since circadian rhythm disruption is associated with other neuropsychic disorders such as bipolar disorder and depression, it is worth investigating if Sarm1 inhibition has effects in those diseases.
Conclusions
In summary, our data demonstrated that Sarm1 knockdown can restore the disrupted circadian rhythm in AD mice in locomotive and cognitive experiments. While AD mice demonstrated deviation in the expression of clock molecules including Bmal1 and Per2 compared to normal control mice. Sarm1 knockdown was associated with circadian rhythm improvement and plaque reduction in mouse cortex and hippocampus. In vitro study also confirmed that Sarm1 knockdown in HT22 hippocampus cells also led to normalization of clock molecules. Our results potentiated Sarm1 inhibition as a viable pharmacological strategy to impede AD progression.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
The authors have no funding to report.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The corresponding author will provide original data upon reasonable request.