
Abstract
Background:
Prior experience in early life has been shown to improve performance in aging and mice with Alzheimer’s disease (AD) pathology. However, whether cognitive training at a later life stage would benefit subsequent cognition and reduce pathology in AD mice needs to be better understood.
Objective:
This study aimed to verify if behavioral training in mid-adulthood would improve subsequent cognition and reduce AD pathology and astrogliosis.
Methods:
Mixed-sex APP/PS1 and wildtype littermate mice received a battery of behavioral training, composed of spontaneous alternation in the Y-maze, novel object recognition and location tasks, and spatial training in the water maze, or handling only at 7 months of age. The impact of AD genotype and prior training on subsequent learning and memory of aforementioned tasks were assessed at 9 months.
Results:
APP/PS1 mice made more errors than wildtype littermates in the radial-arm water maze (RAWM) task. Prior training prevented this impairment in APP/PS1 mice. Prior training also contributed to better efficiency in finding the escape platform in both APP/PS1 mice and wildtype littermates. Short-term and long-term memory of this RAWM task, of a reversal task, and of a transfer task were comparable among APP/PS1 and wildtype mice, with or without prior training. Amyloid pathology and astrogliosis in the hippocampus were also comparable between the APP/PS1 groups.
Conclusion:
These data suggest that cognitive training in mid-adulthood improves subsequent accuracy in AD mice and efficiency in all mice in the spatial task. Cognitive training in mid-adulthood provides no clear benefit on memory or on amyloid pathology in midlife.
INTRODUCTION
Education in early life is strongly associated with preserved cognitive functions in aging and is associated with reduced risk of dementia. Early life education is thought to be a primary contributor for cognitive reserve [1]. High levels of education in children have been shown to associate with preserved cognitive ability in old age [2]. Formal education stretching past childhood has been shown to associate with lower disease-related cognitive decline, as well as degenerative aging that leads to reduced cognitive ability [3, 4]. Whether cognitive training in mid-adulthood provides benefits for subsequent learning and memory and for reducing AD-related brain pathology is understudied. As memory decline can start in midlife in early-onset AD [5] and mild cognitive impairment can be seen in mid-age [6, 7], it is important to understand whether cognitive stimulation in adulthood would be a useful approach to prevent or slow the decline. To address this question without confounding from earlier life experience, we proposed to use an AD mouse model that undergo cognitive training at mid-adulthood.
Early life training can provide cognitive benefit in later life. For example, training of a spatial memory task in the water maze in young mice (e.g., at 2 months old) is shown to make re-learning occur at a faster pace in mid-adulthood [8]. At the brain circuit and receptor level, we have shown that a brief aversive conditioning can change the requirement for N-methyl-D-aspartate (NMDA) receptors in the dorsal hippocampus or the requirement of the extent of the hippocampal formation in subsequent conditioning [9–11]. Prior training can also improve performance in Alzheimer’s disease (AD) mouse models. For example, in an AD model with overexpression of human amyloid-β protein precursor (AβPP), presenilin (PS), and tau proteins, AD mice that received repeated training every 3 months from juveniles to an older age show better spatial memory in the water maze at midlife, compared to AD mice without repeated training [12]. Together, these support the view that early life training is beneficial, while the benefit of later life training remains to be answered. To address this, we further proposed that the effect of cognitive training in mid-adulthood (at 7 months old) would be assessed at an early time point when learning and memory impairment would be seen in an AD mouse model [13, 14], but not in wildtype mice.
Amyloid-β plaques, as a result of protein aggregation, are a key pathology of AD, with early-onset familial AD being caused by mutations in AβPP, presenilin-1 (PS1), and presenilin-2 (PS2) [15–17]. The establishment of these plaques leads to the disruption of synaptic function, disturbance of neural connectivity and begins a cascade of events including alterations in tau that are associated with neuron death [15]. Amyloid-β pathology has been shown to associate with the risk of cognitive decline in humans [18, 19], albeit cognitive normal humans can have amyloid-β pathology [20, 21]. With high amyloid-β load affecting working memory and cognitive performance, the amount of deposition has been linked with performance in humans [22]. APP/PS1 mice which express the Swedish mutation of AβPP (APPswe) as well as the deletion of exon 9 of human presenilin 1 (PS1-dE9), is a commonly used AD mouse model [23]. We used this mouse line to test whether a battery of cognitive training in mid-adulthood at 7 months of age can improve subsequent learning and memory at 9 months old. This age was chosen to reflect mid-adulthood, prior to middle age, in humans [24]. We designed a sequence of training, from tasks requiring a brief session with spontaneous exploration to tasks requiring more intensive training with spatial navigation. These included a spontaneous alternation task in the Y-maze, a novel object recognition task, a novel object location task, and a series of training, reversal training, and transfer training in a radial-arm water maze (RAWM) and in an open field water maze. Short-term and long-term memory tests after training, reversal and transfer were also arranged to assess the spatial memory. As these mice show significant plaque burden in the hippocampus [23] and learning deficits [25–27] at 9 months old, we compared the behavioral performance and amyloid-β plaque load in the hippocampus at this age between animals receiving prior training at 7 months old or controls without training. Glial fibrillary acidic protein (GFAP)-positive astrocytes, which are highly reactive, are associated with amyloid-β plaques and inflammatory processes in AD, and have been shown to be significantly increased in the hippocampus of APP/PS1 mice from 6 months old [28, 29]. Hence, we additionally asked if prior training would ameliorate astrogliosis in the hippocampus of APP/PS1 mice.
We found benefits of prior training in reducing errors made in searching for the escape platform, an effect which was concluded as prior training ameliorated the number of errors made by APP/PS1 mice. Prior training also reduced the time needed for finding the platform in all mice. Our training protocol was sufficient to result in robust short-term and long-term memories after training. Prior training did not reduce plaque areas, counts, or astrogliosis in the hippocampus.
MATERIALS AND METHODS
Animals
Thirty-five hemizygous APP/PS1 transgenic (Tg) mice and wildtype (Wt) littermates were bred from feeder mice expressing the human Swedish mutation of AβPP and human presenilin-1 with an exon 9 deletion under the control of the Thy1 promoter (Jax 34829) [23]. Same sex, mixed genotypes were group-housed at 2 to 4 per cage. A 12 h light-dark schedule was maintained, and behavioral tests were conducted during the light phase. Three Tg animals remained in their home cages and received no handling or training as sedentary control for pathology. Thirty-two animals were handled for 2 min daily for 5 days a week before the age of 7 months. One group of mice received prior training at 7 months (Fig. 1A; n = 16; Wt = 8, 5 of which were male; APP/PS1 = 8, 5 of which were male) and the other group received handling for the matching number of days when prior training would take place (n = 16; Wt = 7, 5 of which were male; APP/PS1 = 9, 7 of which were male). Both groups received subsequent training at 9 months of age. One male Wt mouse died between the two training phases resulting in n = 16 for the prior training at 7 months and n = 15 for the subsequent training at 9 months. Animals had ad libitum access to food and water. All experiments were carried out under an approved Project Licence from the University of Edinburgh’s Animal Welfare and Ethical Review Body and UK Home Office. Experiments also received approval from the Experimental Request Team at the Bioresearch & Veterinary Service of the University of Edinburgh, which provided additional checks on animal welfare and ethics. All protocols were in accordance with the Home Office Animals Scientific Procedures Act 1986 (amended 2012). The study was designed according to the ARRIVE guidelines. All methods were carried out in accordance with relevant guidelines and regulations.
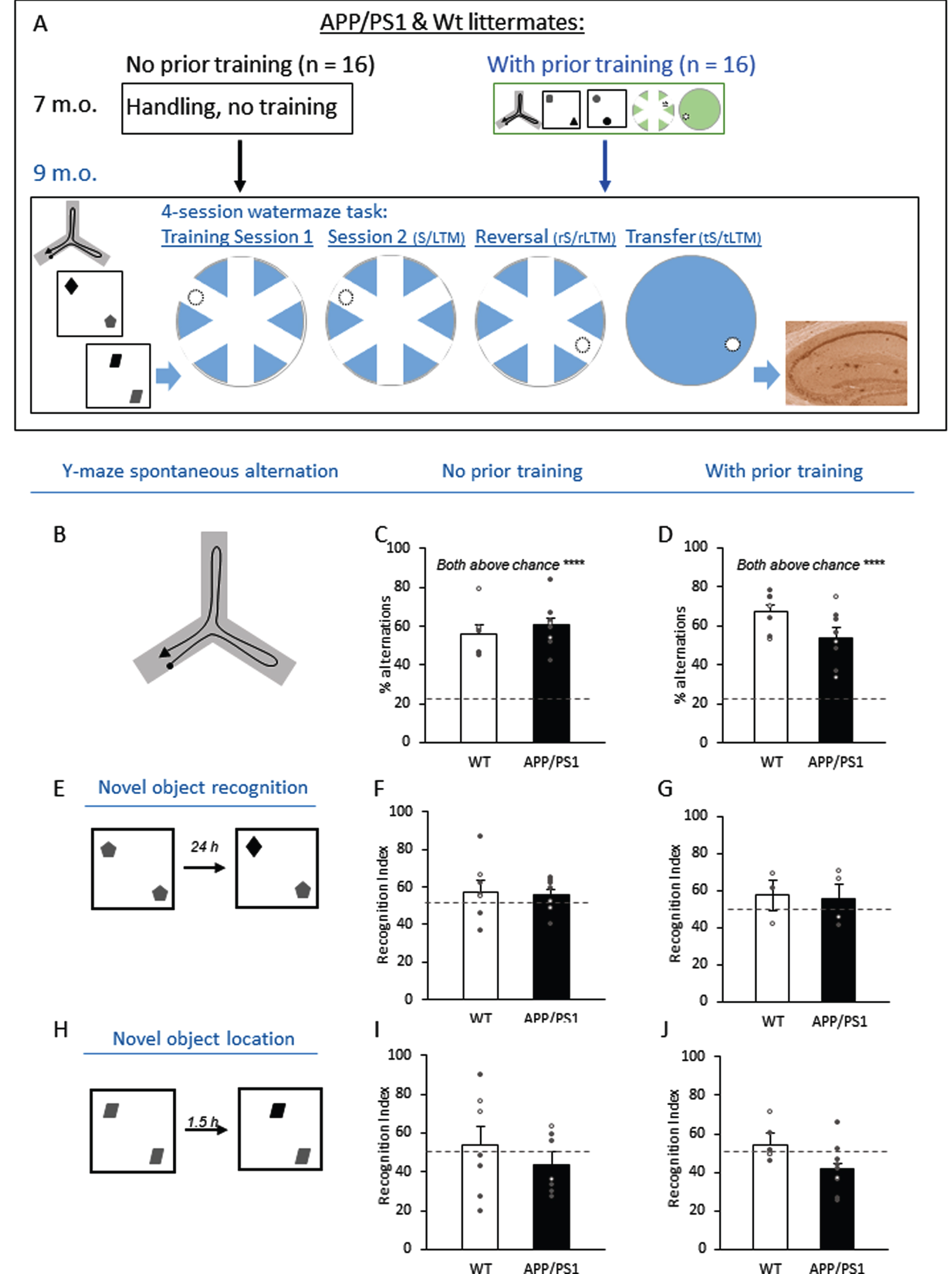
Experimental design and performance in the Y-maze, novel object recognition, and novel location tasks at 9 months. A) Two groups of 7-month-old APP/PS1 and wildtype littermates received handling (i.e., no prior training) or prior training of a spontaneous alternation test in a Y-maze, a novel object recognition task, a novel location recognition task, and a 4-session water maze task with training, short-term, and long-term memory (S/LTM) tests, reversal training, reversal STM and LTM (rS/LTM) tests, and transfer training, transfer STM and LTM (tS/LTM) tests. All mice were trained and tested at 9 months of age with similar tasks but with different objects or in a different visuospatial water maze. Results from the 9-month-old mice are reported in subsequent figures. B) Schematic diagram of the Y-maze with an example path (black arrow). C, D) All groups performed significantly above chance (22%, dashed line) and there were no significant genotype effects. E) Schematic diagram of the novel object recognition box with objects for encoding (left) and test (right). F, G) All groups performed insignificantly above chance (50%, dashed line) and there were no significant genotype effects. H) Schematic diagram of the novel object location box with objects for encoding (left) and test (right). I, J) All groups performed insignificantly above chance (50%, dashed line) and there were no significant genotype effects. Data are presented as mean±SEM. *p < 0.05, **p <0.01, *** p < 0.005, **** p < 0.001.
Apparatus
The Y-Maze was composed of 3 white acrylic arms (7×12×40 cm) and a central triangular area (equilateral 7 cm, height 12 cm). The box for object exploration was made of clear acrylic walls (40×40×30 cm) and the floor was lined with approximately 0.5 cm of fresh sawdust. Objects were placed in diagonally opposite corners (i.e., north-west and south-east) at 10 cm away from either wall for encoding. For the novel object recognition task at 9 months of age, the objects were a pair of metal peppershakers and glass saltshakers (approx. 4×4×8 cm). For prior training, the objects were a pair of red phone box models (5×5×9 cm) and white 3-pin plugs (5×5×5 cm). The identity and location of objects were counterbalanced within each group and kept similarly between groups. For example, half of the mice would have object 1 as the pair at encoding, with object 2 used as the novel object at testing, while the other half of the animals would have object 2 as the pair at encoding and object 1 as the novel object at testing. Objects used for novel object location were a pair of grey cylindrical tins (dia. 6 cm×10 cm) at 9 months old and a pair of blue-cap Pyrex bottles (50 mL) at prior training. The novel location would be at 10 cm away from the center of the north or south wall.
The radial-arm water maze (RAWM) was composed of 6 identical triangular acrylic inserts placed at equal distance in a 1 m diameter pool (depth 45 cm). This created 6 arms of equal length (35 cm) and width (16 cm) and a hexagonal center zone. To ensure the escape platform (11 cm in diameter, and 2 cm below the surface of the water) remained invisible to the animals, 20 mL of white non-toxic tempera paint was added in the water. For the first block of training, visibility of the platform was created by attaching a 10 cm stick with an inflated surgical glove to the platform. The temperature of the water was maintained at 23±1°C. Black, green, or vibrant yellow geometric cue cards were placed on white walls around the maze to provide visible spatial cues. Green triangles, a black cross, 4 black rectangles, and a yellow star were placed at 4 zones as spatial cues in the training at 9 months (Fig. 2A, inset), while a black circle, a stripy rectangle, a check-pattern rectangle, and a green triangle were used for the group receiving prior training at 7 months of age. The triangular acrylic inserts were removed for the transfer training and tests in the open water maze.
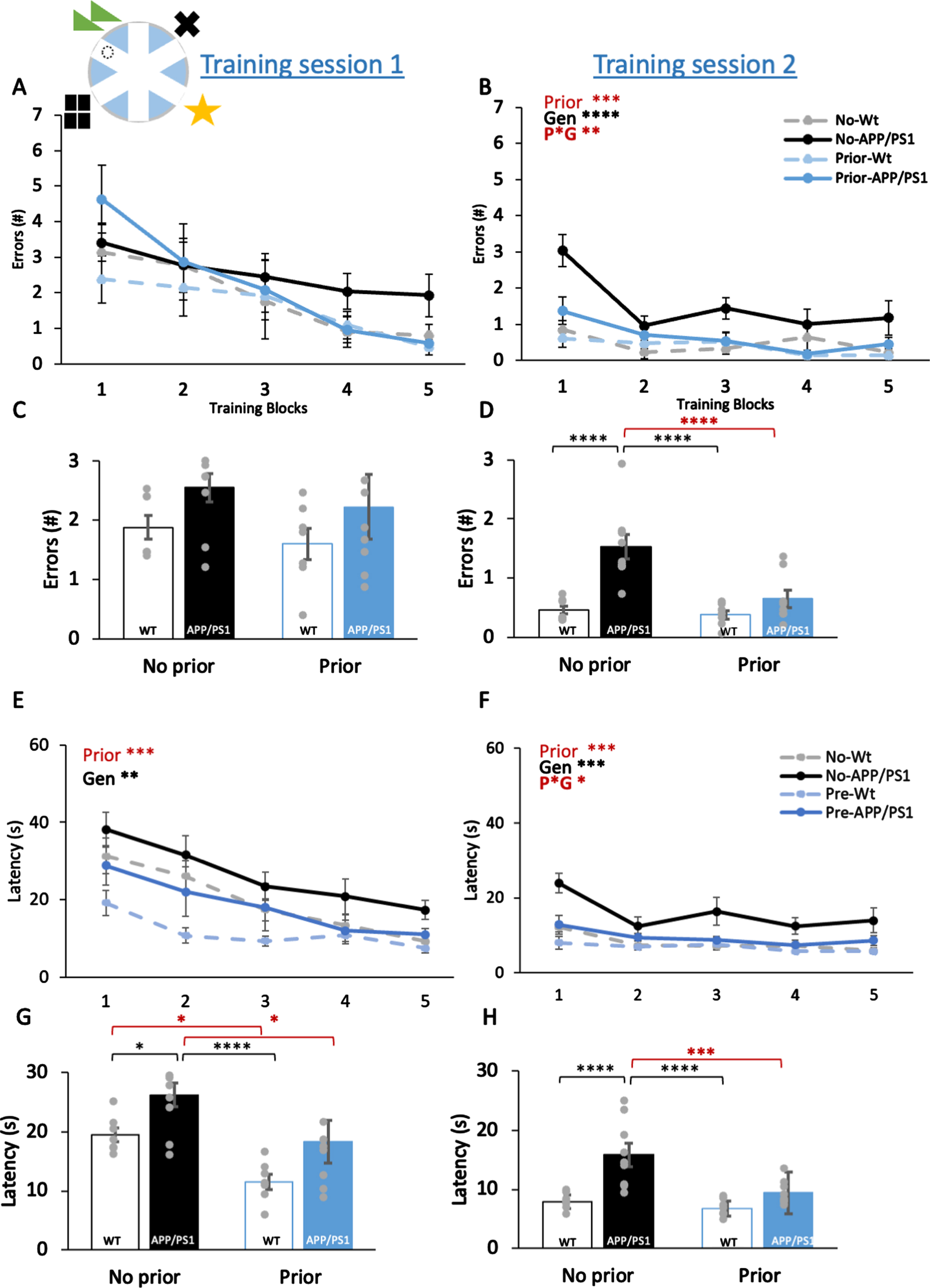
Prior training reduces errors in AD mice and latencies in both genotypes in the radial-arm water maze task. A) Inset: A schematic drawing of the RAWM, platform location (dashed circle) and surrounding visuospatial cues in geometric shapes. In training session 1, the number of errors reduced across 5 blocks but were not significantly affected by prior training or genotype in 9-month-old mice. B) In training session 2, the number of errors reduced across 5 blocks. Prior training (Prior) significantly reduced the errors, the genotype (Gen) and the Prior*Gen (P*G) interaction were significant. C) Averaged errors in session 1 were not significantly different among 4 groups. D) Averaged errors in session 2 were significantly higher in no prior training, APP/PS1 group than 3 other groups. E) In training session 1, latencies reduced across 5 blocks. Prior training and genotype effects were also significant. F) In training session 2, latencies reduced across 5 blocks. Prior training and genotype effects and the interaction were significant. G) Averaged latencies in session 1 were significantly different among 4 groups. No prior training, APP/PS1 group showed significantly higher latencies than 3 other groups. In wildtype groups, prior training significantly reduced the latency. H) Averaged latencies in session 2 were significantly different among 4 groups. No prior training, APP/PS1 group showed significantly higher latencies than 3 other groups. Data are presented as mean±SEM. *p < 0.05, **p < 0.01, ***p < 0.005, **** p < 0.001, § p < 0.06.
Behavioral procedures and measurements
The order of training APP/PS1 or wildtype mice was randomized and counterbalanced. It was kept the same for the following training tasks. The experimenters were blind to the genotype of the mice when conducting the studies. The behavioral indices were collected ‘blindly’ with the scorers not knowing the genotype of the mice. Errors for the RAWM tasks were scored manually as the animal could only be in a correct or incorrect arm at a given time. The water maze training and testing was recorded via an overhead camera, using ANY-maze version 6.30 (Stoelting Co., Wood Dale, IL) for latency and speed quantification. The Y-maze, NOR, and NOL were also recorded via an overhead camera. The sequence of alternation in the Y-maze was manually noted and the time spent with each object was manually timed with the experimenters remaining blind to the genotypes or object identity.
Spontaneous alternation in a Y-maze
After 5 days of handling, mice were placed in the center of the maze and allowed to freely explore it for 5 min. Behavior in the maze and arm entries were recorded via an overhead camera. One full alternation was scored when the mouse consecutively entered 3 different arms. The percentage of alternation was calculated by: (total number of consecutive alternations) / (the total number of entries -2) * 100.
Novel object recognition (NOR) task
Animals were habituated to the box without objects for 3, 10 min sessions across 3 days. On the next day, they received a 10 min encoding trial in which they were placed in the center of the box with two identical objects. After a retention interval of 24 h, mice received a 5 min test trial during which one of the two familiar objects was replaced with a novel object. The identity and the location of the objects being familiar, or novel, were counterbalanced between encoding and testing trials and between groups. There was no significant difference between the natural preference of the pairs of objects being used in the study. Exploration was defined as sniffing the object, nose directed at object within 2 cm, and rearing with paws on the object, but not sitting on top of the object. The recognition index for NOR was calculated as the percentage of time spent exploring the novel object over the total time spent exploring both objects.
Novel object location (NOL) task
As box habituation had already been conducted prior to NOR, NOL was conducted without further habituation. Mice were placed in the center of the same box in the presence of two identical objects for a 10 min encoding trial. After a retention interval of 90 min (or 3 h at prior training), a 5 min test trial was conducted during which the mice were placed in the box again but one of the objects had been moved to a novel location. The object and the location of relocated object were counterbalanced between groups. There was no significant difference in preference for either side of the location in this study. In both NOR and NOL tasks, objects were wiped with 70% ethanol between trials and before the first trial. The definition of exploration remained the same as NOR. The recognition index of NOL was calculated as the percentage time spent exploring the displaced object over the total time spent exploring both objects.
Four-session training and memory tests in the water maze (Fig. 1A)
Mice received 4 sessions of water maze training. Sessions 1–2 were designed to assess the animals’ spatial learning and memory. Session 3 was designed to test if APP/PS1 mice would make more errors to learn the changed location in reversal training and if prior training would improve it. Session 4 was designed to test if APP/PS1 mice would take longer to ‘transfer’ the spatial information to a modified environment and if prior training could facilitate transfer learning.
Procedures
Mice received 4 sessions of water maze training. In session 1 on day 1 in the RAWM, they would receive 3 training trials of swimming per block for 5 blocks. The inter-trial interval was 1–2 min; inter-block interval was 15–25 min. Session 2 on day 2 was a repeat of session 1, except that trial 12 was a short-term memory (STM) test. A long-term memory (LTM) test was performed on the next day (24 h after the first trial on the previous day).
In session 3 on day 3 in the RAWM, they would receive 3 training trials per block for 4 blocks with a new platform location at the opposite arm from session 2. The inter-trial interval was 1–2 min; inter-block interval was 15–20 min. This was called reversal learning and occurred 20 min after the LTM test. A reversal STM (rSTM) test was introduced at the 9th trial. A reversal LTM (rLTM) test was performed on the next day.
In session 4 on day 4 in an open water maze (i.e., the same maze without arm inserts), they would receive 3 training trials per block for 3 blocks with the same platform location as day 3. This was called transfer learning and occurred 20 min after the rLTM test. A transfer STM (tSTM) test was introduced at the 6th trial. A transfer LTM (tLTM) test was performed on the next day.
Training trials
The mouse would be released from a start location, swim and locate the escape platform, remain on it for 15 s, and be remove and dried by a towel on a heating mat. The trials were capped at 60 s. If the animal did not find the platform within 60 seconds, it was gently guided to the platform by the experimenter. The next trial would start at a different location. The start location of the start arm, except the arm with the platform would be randomized across trials. The platform location would be in the same position for a mouse, not repeated among 3 consecutive animals, and counterbalanced across groups.
Memory tests
The mouse would swim freely for 60 s (no platform) in the maze and be removed and dried.
For training measurements, errors, and latencies in finding the escape platform were recorded for sessions 1–3. An arm entry was defined as the whole body of the mouse passing through an arm entrance. Swimming speed in the first and last blocks of session 1 was further analyzed with ANY-maze version 6.30 (Stoelting Co., Wood Dale, II). STM, LTM, rSTM, and rLTM were assessed by measuring the percentage of time spent in the correct arm over time in all 6 arms. For training in session 4, latencies in finding the escape platform were recorded. tSTM and tLTM were assessed by the percentage time spent in the correct quadrant divided by the time spent in all quadrants and multiplied by 100.
Tissue preparation and immunohistochemistry
Mice were deeply anaesthetized with pentobarbital and perfused transcardially with 1×phosphate buffered saline (PBS) then 4% paraformaldehyde (PFA). Brains were extracted and kept in 4% PFA for 3 h. Brains were sectioned at 40μm thickness in sagittal sections using a vibratome (Leica, VT1000 S) and kept in 0.2% sodium azide PBS solution at 4°C. Sections encompassing the hippocampus were collected. Sections (3–4) at equal distance around 0.96–1.8 mm lateral to the midline were processed for immunohistochemistry and quantified.
Free-floating sagittal brain sections were washed in 1×PBS for 15 min, blocked with 1% H2O2 PBS for 10 min then washed in PBS for 3 times at 10 min each. Sections were incubated in 10% normal goat serum with 0.1% Triton™ x–100 (Sigma Aldrich) in PBS for 1 h and then in primary antibody overnight (mouse anti-β-Amyloid, 1–16, 6E10, 1:500, Biolegend, or rabbit polyclonal anti-GFAP, 1:3000, Dako). Following 3, 10 min washes in PBS, sections were incubated in the secondary antibody solution for 2 h (1:100 anti-mouse or anti-rabbit Biotin, Sigma Aldrich) and then in Avidin-Biotin complex (ABC elite, standard, Vectastain) for 30 min. Finally, 3.3’-diaminobenzine (DAB Substrate Kit, Peroxidase (HRP), with Nickel, Vector Laboratories) was applied to brain sections for 3 min. All sections were dehydrated and mounted on glass slides followed by coverslipping with dibutylphthalate polystyrene xylene (DPX, Sigma Aldrich).
Brain sections containing the hippocampus were imaged using a Carl Zeiss, Axio Scan.Z1, with an exposure time of 200us and 1.72μm depth of focus. Images were analyzed using StereoInvestigator™ and Neurological™ software to measure the percentage of areas with positive staining for both 6E10 and GFAP. An average was taken across these sections for an animal and the data from individual animals were used for statistical analyses. The images were quantified ‘blindly’ so that the experimenters did not know the genotype of the sections.
Statistical analyses
Data are presented as percentage of alternation for the Y-maze task, recognition index from the percentage of time in exploring the novel object for NOR, and recognition index from the percentage of time in exploring the new location for NOL. For training in the RAWM, errors and/or latencies were used for analyses. For memory tests, the percentage of time swimming in the correct quadrant was analyzed. Group data was presented as mean±SEM. Genotype of the data was revealed when all measurements were collected. Three-way ANOVAs were used to analyze the effects of prior training, genotype, training blocks, and their interactions. If the assumption of sphericity was adhered, sphericity-assumed statistics were reported. If it was violated, Greenhouse-Geisser correction was applied. Two-way ANOVAs were used to analyze the effects of prior training, genotype, and the interaction. Post hoc (Fisher’s Least Significant Difference, LSD) tests were conducted to identify the source of difference among groups that contributed to significant main effects and/or interaction in the RAWM task. Two-tailed, unpaired t-tests were used to verify the genotype effect or gender effect. A χ2 test was used to assess gender imbalance. Two-tailed, one-sample t-tests were used to verify whether the measurement was significantly different from chance. Correlations were analyzed via Pearson’s correlation. All analyses were done with IBM SPSS Statistics (v.25). Type 1 error was set at 0.05.
RESULTS
The effects of prior training and genotype in spatial learning and memory
Prior training did not affect working memory in the Y-maze or recognition memory with objects
In the Y-maze task (Fig. 1B), spontaneous alternation among 3 arms was measured and the percentage of sequential alternation among all turns was calculated. The genotype effect was not significant at 9-month-old in the group without prior training (Fig. 1C; t14 = 0.76, p = 0.46) or in the group with prior training (Fig. 1D; t13 = 2.05, p = 0.06). Critically, all performances were significantly above chance (22.22%; all t6–8 > 6.22, p < 0.001), which indicates that all groups performed this task well. These results suggest that both APP/PS1 and wildtype mice have an intact working memory.
In the novel object recognition task (Fig. 1E), time investigating both the novel and familiar objects were measured and the percentage of time for exploring the novel object was calculated as the recognition index. There was no significant effect of genotype in groups without prior training (Fig. 1F; t14 = 0.58, p = 0.57), or with prior training (Fig. 1G; t5 = 0.15, p = 0.89). Regardless of prior training, performances were not above chance (50%; all t2–8 < 1.68, p > 0.13). These results suggest an age-dependent decline (compared with data from 7 months old) as none of the groups were able to recognize the novel object after a 24 h retention delay at 9 months old.
In the novel object location task (Fig. 1H), time spent exploring the objects in the novel and familiar locations were measured and the percentage of time for exploring the novel location was calculated as the recognition index. Although the genotype effect was significant for the group without prior training (Fig. 1I; t14 = 2.23, p = 0.043) and not significant for the group with prior training (Fig. 1J; t12 = 0.88, p = 0.40), all groups’ performances were not significantly different from chance (50%; all t6–8 < 1.24, p > 0.08). These results suggest that none of the groups show a preference for the novel location after a 1.5 h retention delay.
APP/PS1 mice performed less well, while prior training in midlife improved performance in the radial-arm water maze
The number of errors (i.e., times mice entered the wrong arm that had no escape platform) was used to indicate the accuracy in learning in the RAWM (Fig. 2A, inset). Over the 5 blocks of training in session 1, mice showed significant reduction in errors made for searching the platform (Fig. 2A; F3.23,87.21 = 10.15, p < 0.001). However, there were no effects of prior training, genotype, or interaction (all F1,27 < 3.43, p > 0.08). All other two-way or three-way interactions were also insignificant (all F3.23,108 < 1.04, p > 0.38). This suggests that while all mice improve performance over training, mice with prior training can find the platform significantly quicker and learn the task more efficiently.
Across the 5 training blocks in session 2, a significant decrease in the number of errors was observed (Fig. 2B; F4,108 = 8.89, p < 0.001), suggesting all animals further improved their learning in the second session. Animals with prior training made significantly less errors (F1,27 = 9.88, p = 0.004), while APP/PS1 mice made more errors than wildtype mice (F1,27 = 19.44, p < 0.001). Importantly, the interaction between prior training and genotype was also significant (F1,27 = 6.81, p = 0.02), suggesting prior training reduces the impairment in learning cause by APP/PS1 mutations. The interaction between training blocks and genotype was significant (F4,108 = 2.99, p = 0.02), most likely due to APP/PS1 mice making more errors in early training blocks. No other two-way or three-way interactions were significant (all F4,108 < 1.637, p > 0.17). This suggests that prior training improves later learning in both wildtype and APP/PS1 mice, with a greater effect in the transgenic animals which initially performed poorer, an effect that is also observed in the latency of trials predominantly in the second session.
Errors in training sessions 1 (Fig. 2C) or session 2 (Fig. 2D) were averaged for group comparisons by post hoc tests. No significant group difference was found in session 1 (Fig. 2C; p = 0.1 –0.6), which is likely due to within-group variation and all animals needing to familiarize with the task requirement (e.g., registering the environment cues and learning the rule of the task) in the early phase. In session 2, APP/PS1 mice without prior training made more errors than 3 other groups (Fig. 2D; all p = 0.04 –0.001), while no significant difference was found among the 3 other groups (p = 0.2 –0.7). Toward the later stage of training, wildtype mice made very few errors and prior training did not improve the accuracy further. This would suggest that prior training at mid-adulthood provides benefit in accuracy in performing the spatial task in AD mice.
Latencies (s) in finding the escape platform were used to indicate the efficiency of performing the task. Over 5 training blocks in session 1, mice showed significant reduction in latency (Fig. 2E; F2.75,74.19 = 17.11, p < 0.001). Prior training and genotype effects were significant (F1,27 = 11.4 and 8.49 respectively, both p < 0.01). None of the interactions was significant (all F1,27 < 1.01, p > 0.38). This suggests that prior training improves efficiency regardless of the genotype from early on. Over 5 training blocks in session 2, a significant decrease in the latency was again observed (Fig. 2F; F4,108 = 7.69, p < 0.001). Animals with prior training were significantly quicker at finding the platform than animals without prior training (F1,27 = 9.69, p = 0.004). APP/PS1 mice took longer to find the platform than wildtype mice (F1,27 = 18.5, p < 0.001). The interaction between prior training and genotype was also significant (F1,27 = 4.68, p = 0.04). None of other interactions was significant (all F4,108 < 1.61, p > 0.18).
Latencies in training sessions 1 (Fig. 2G) or session 2 (Fig. 2H) were averaged for group comparisons by post hoc tests. Significant group differences were already apparent in session 1 (Fig. 2G). APP/PS1 mice without prior training took longer to find the platform than 3 other groups (all p = 0.001 –0.047). Prior training reduced latencies in wildtype (p = 0.03) and APP/PS1 (p = 0.02) mice. This would suggest that prior training at mid-adulthood provides benefit in efficiency in the spatial task in all mice, regardless of genotypes. In session 2 (Fig. 2H), APP/PS1 mice without prior training still took longer to find the platform than 3 other groups (all p = or< 0.001), while no significant difference was found among the 3 other groups (p = 0.2–0.5). The benefit of prior training in efficiency remained in AD mice throughout both sessions. The benefit subsided in the wildtype mice as they were very quick at finding the platform after successive training.
Swimming speed (m/s) was measured and no significant prior training or genotype effects on speed (m/s) were found during the first block (all F1,27 < 2.85, p > 0.1, data not shown) or the last block of session 1 training (F1,27 < 3.75, p > 0.06, data not shown). This suggests that the mice do not complete the task faster as a result of swimming faster, and that there is no genotype difference in the APP/PS1 mice.
The STM test in the RAWM (Fig. 3A) showed that all performed significantly above chance (16.67%, Fig. 3B; all t6–8 > 10.21, p < 0.008). There was no significant effect of genotype, prior training, or interaction in the STM test (all F1,27 < 3.46, p > 0.07). They also showed robust LTM with the performance significantly above chance (Fig. 3C; all t6–8 > 3.70, p < 0.008). The prior training effect, genotype effect, and interaction were all insignificant (Fig. 3C; all F1,27 < 2.3, p > 0.14). These suggest that good STM and LTM can be maintained in APP/PS1 mice after sufficient training.

Prior training improved performance in the transfer task but did not affect the reversal learning or various memory tests. A) A schematic diagram of the RAWM, platform location (dashed circle) and surrounding visuospatial cues in geometric shapes. B, C) In the short-term memory (STM) and long-term memory (LTM) probes, all groups performed significantly above chance (16.67%, dashed line). There were no significant effects of genotype or prior training. D) A schematic diagram of the RAWM in reversal configuration with the platform location (dashed circle) moved to the opposite arm. E) In the reversal training session, the number of errors decreased across the 3 blocks. There were no significant prior training or genotype effects. F, G) In the reversal short-term memory (rSTM) and long-term memory (rLTM) probes, all groups performed significantly above chance (16.67%). There were no significant effects of genotype or prior training. H) A schematic diagram of the open field water maze with platform location (dashed circle) and arms removed. I) In the transfer training session, all mice improved in latency (s) to reach the platform across the 3 blocks. Prior training (Prior) significantly improved latency and the genotype (Gen) effect was significant. J, K) In the transfer short-term memory (tSTM) and long-term memory (tLTM) probes, most groups performed significantly above chance (22.7%). No significant prior training or genotype effects. Data are presented as mean±SEM. *p < 0.05, **p < 0.01, *** p < 0.005, **** p < 0.001.
Reversal learning was comparable between APP/PS1 mice and prior training did not improve this
Reversal learning was carried out in the RAWM with the platform now relocated to the opposite arm to gauge the animals’ ability to reverse the learning (Fig. 3D). All mice gradually made less errors in finding the platform across training blocks (Fig. 3E; F1.51,54 = 22.57, p < 0.001). Prior training, genotype or interaction were all insignificant (all F1,27 < 0.97, p > 0.33), and there were no significant two-way or three-way interactions (all F1.51,54 < 2.89, p > 0.064).
Reversal short-term and long-term memory probe tests were also carried out and time spent in the correct arm was calculated as previously shown. All performances were significantly above chance (all t6–8 > 9.99 p < 0.021). No significant effects of prior training, genotype or interaction were seen for the reversal short-term probe (Fig. 3F; F1,27 < 2.54, p > 0.12) or the long-term probe (Fig. 3G; F1,27 < 0.97, p > 0.33). Together, these suggest that all the mice were able to learn and engage memory for the reversal task effectively, but there was no improvement on either learning or memory as a result of prior training.
Prior training improves transfer learning in wildtype and APP/PS1 mice
All animals were then trained in the open field water maze (Fig. 3H) and latency (s) to reach the platform in each trial was measured. This was averaged into three blocks of trials. Across the three blocks, all mice significantly improved (Fig. 3I; F2,54 = 6.8, p = 0.002). Both a significant effect of genotype and prior training were observed (F1,27 > 5.05, p < 0.03), but no significant interaction (F1,27 = 0.594, p = 0.448). There were no other significant two- or three-way interactions (all F2,54 < 1.12, p > 0.334). This suggests APP/PS1 mice do not find the platform as quickly as their wildtype littermates, but prior training is beneficial across groups in reducing the time needed for the transfer learning.
Transfer short-term (Fig. 3J) and long-term memory (Fig. 3K; tSTM and tLTM) tests were carried out and percentage time in the correct goal quadrant (%) was calculated. All performances were significantly above chance (25%, all t6–8 > 7.47, p < 0.048), except APP/PS1 mice without prior training in the short-term memory probe (t8 = 1.75, p = 0.118). However, prior training, genotype or interaction were all insignificant for tSTM (all F1,27 < 3.75, p > 0.06) and tLTM (all F1,27 < 0.71, p > 0.41). Together, these suggest that prior training improves transfer learning but does not additionally improve memory when good memory is already achieved without prior training.
The effects of prior training and genotype on brain pathology
Prior training did not ameliorate the amyloid-β pathology in APP/PS1 mice
The brain sections were stained for amyloid-β using 6E10 antibody (Fig. 4A). We first quantified the percentage of hippocampus area that is 6E10-positive and found, as predicted, APP/PS1 hippocampi were significantly occupied with amyloid-β (Fig. 4B; genotype effect, F1,27 = 97, p < 0.001). The prior training or the interaction between genotype and prior training were both insignificant (F1,27 < 0.79, p > 0.38). When counting the number of amyloid-β plaques, the same pattern of effects was observed (Fig. 4C; significant genotype effect, F1,27 = 43.94, p < 0.001; insignificant prior training and interaction, F1,27 < 2.38, p > 0.134). Both groups were not significantly different from the sedentary control in % of 6E10-positive area (Fig. 4B; t10 = –0.71, p = 0.5, for no prior training group; t9 = –1.98, p = 0.08 for the prior training group) or in plaque counts (Fig. 4c; t10 =–1.26, p = 0.24, for no prior training group; t9 =–0.26, p = 0.8 for the prior training group). When quantifying GFAP-positive areas, as an estimation of reactive astrocytes, in the hippocampus (Fig. 4D), none of the genotype, prior training, or interaction effects were significant (Fig. 4E; all (F1,27 < 2.382, p > 0.26). Both groups were not significantly different from the sedentary control in % of GFAP-positive area (Fig. 4E; t10 =–1.19, p = 0.26, for no prior training group; t9 =–0.27, p = 0.79 for the prior training group). Together, these suggest that prior training did not ameliorate brain pathology indicated by amyloid- β and GFAP.
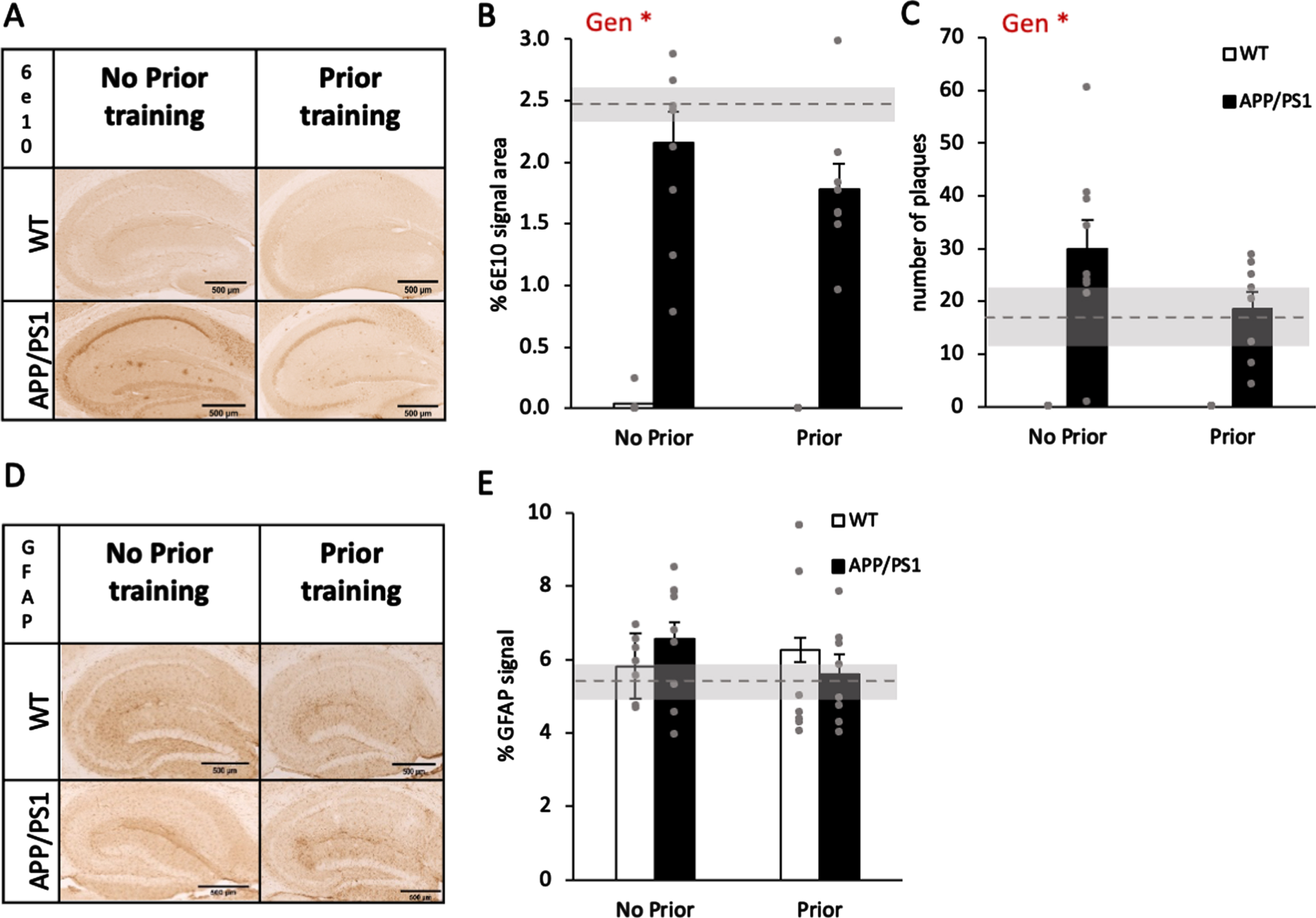
Prior training did not reduce AD-related pathology in APP/PS1 mice compared with sedentary controls. A) Representative images of 6E10 expression in the hippocampus of 9-month-old wildtype and APP/PS1 mice. B) The percentage of 6E10-positive area was significantly higher in APP/PS1 mice. There was no significant effect of prior training. Both APP/PS1 groups were not significantly below sedentary controls (2.48±1 SEM; dashed line and gray zone). C) The number of amyloid-β plaques in the hippocampus of APP/PS1 mice was higher than in wildtype mice but there was no significant prior training effect. Both APP/PS1 groups were not significantly above sedentary controls (17±1 SEM; dashed line and gray zone). D) Representative images of GFAP expression in the hippocampus of 9-month-old wildtype and APP/PS1 mice. E) There were no significant genotype or prior training effects. All groups were not significantly above sedentary controls (5.37±1 SEM; dashed line and gray zone). Data are presented as mean±SEM. *p < 0.05, ** p < 0.01, *** p < 0.005, **** p < 0.001.
Amyloid-β pathology correlates with several learning indices
To explore the correlation among learning indices (averaged errors in training session 1, in session 2 and in reversal training; latencies in transfer training, in training session 1 –first block, in training session 1 –last block, in training session 2 –first block, and in training session 2 –last block) and brain pathology (6E10- and GFAP-positive area), we performed Pearson correlation on a 10×10 matrix (Fig. 5A). To ensure sufficient samples for the exploration, data from all groups were put together and genotype or prior training effects were not tested. This allowed a full exploration into the relationships between all learning indices as well as the pathology measured.
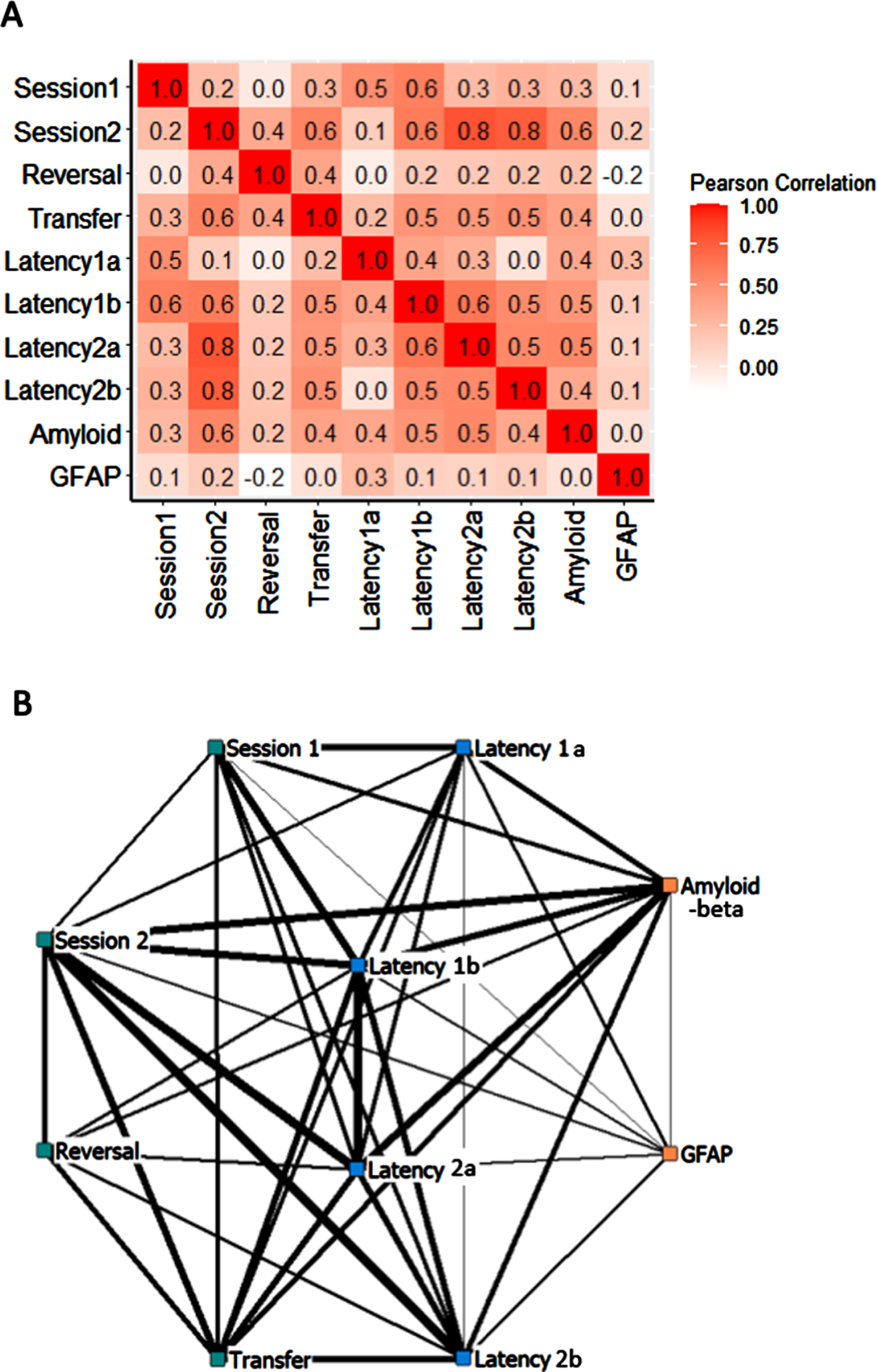
Strong correlation between relevant training sessions as well as between Amyloid pathology and several learning indices. A) Pearson correlation analyses between 8 learning indices and 2 pathology indices in 9-month-old mice. All r values in the cells > 0.4 were significant at p < 0.03. All r values in the cells > 0.5 were significant at p < 0.005. All r values in the cells > 0.6 were significant at p < 0.001. Sessions 1–2: averaged errors in training sessions 1–2 in the RAWM. Reversal: averaged errors in reversal training. Transfer: averaged latencies in transfer training. Latency 1a-1b: averaged latencies in the first block (1a) and last block (1b) of training session 1 in the RAWM. Latency 2a-2b: averaged latencies in the first block (2a) and last block (2b) of training session 2 in the RAWM. Amyloid: the percentage of 6E10-positive areas in the hippocampus. GFAP: the percentage of GFAP-positive areas in the hippocampus. B) A network graph summarizing correlation among indices. Weight of lines indicates higher correlation between variables.
Strongest correlations (r > 0.6) were found between errors in training session 2 and latencies in the 3 later training blocks (r = 0.61 –0.81, all p < 0.001). Strong correlations (r > 0.5) were found between errors in training session 1 and latencies in session 1 (r = 0.52–0.59, both p < 0.005), between errors in training session 2 and transfer latencies (r = 0.57, p < 0.005), and between latencies in the last block with the latencies in the previous 2 blocks (r = 0.52–0.54, both p < 0.005). Moderate correlations (r > 0.4) were found between transfer training and latencies in 3 earlier training blocks (r = 0.46–0.5, all p < 0.01).
Strong to moderate correlations were also found between amyloid pathology and learning. The percentage of 6E10-positive area is positively correlated with errors and latencies in training session 2 (r = 0.55, both r = 0.001). It was also positively correlated with latencies in session 1 (r = 0.48, p < 0.01), and with transfer training (r = 0.4, p < 0.03). Such correlations are likely driven by more amyloid pathology and poorer performance in APP/PS1 mice than in the wildtype mice. None of the correlation between GFAP-positive area and 8 learning indices was significant (all r < 0.26, p > 0.16).
To provide visualization of these positive correlations, a network graph was presented in Fig. 5B. Thicker lines represent stronger correlations. The network suggests that more errors in learning were correlated with longer latencies to find the target. Transfer learning ability was correlated with earlier learning performance, while reversal learning shows weaker correlation as such. More amyloid pathology in the hippocampus was correlated with more errors and longer latencies in intermediate phase of learning.
Performance during prior training at 7 months
In the Y-maze, the 7-month-old groups did not show a significant effect of genotype (Fig. 6A, B; t14 = 1.85, p = 0.09), and both groups’ performance was very significantly above chance (22.22%; both t7 > 11.45, p < 0.001). There was no significant effect of genotype in the NOR task (Fig. 6C, D; t14 = 1.16, p = 0.27), with both groups performing significantly above chance (50%; both t7 > 4.32, p < 0.003). In the NOL task, no genotype effect was seen (Fig. 6E, F; t14 = 1.03, p = 0.32) and neither of the groups performed above chance (50%, both t7 < 0.31, p > 0.31), suggesting these mice were unable to learn this task effectively. Together, APP/PS1 and wildtype mice learn and remember these tasks similarly.
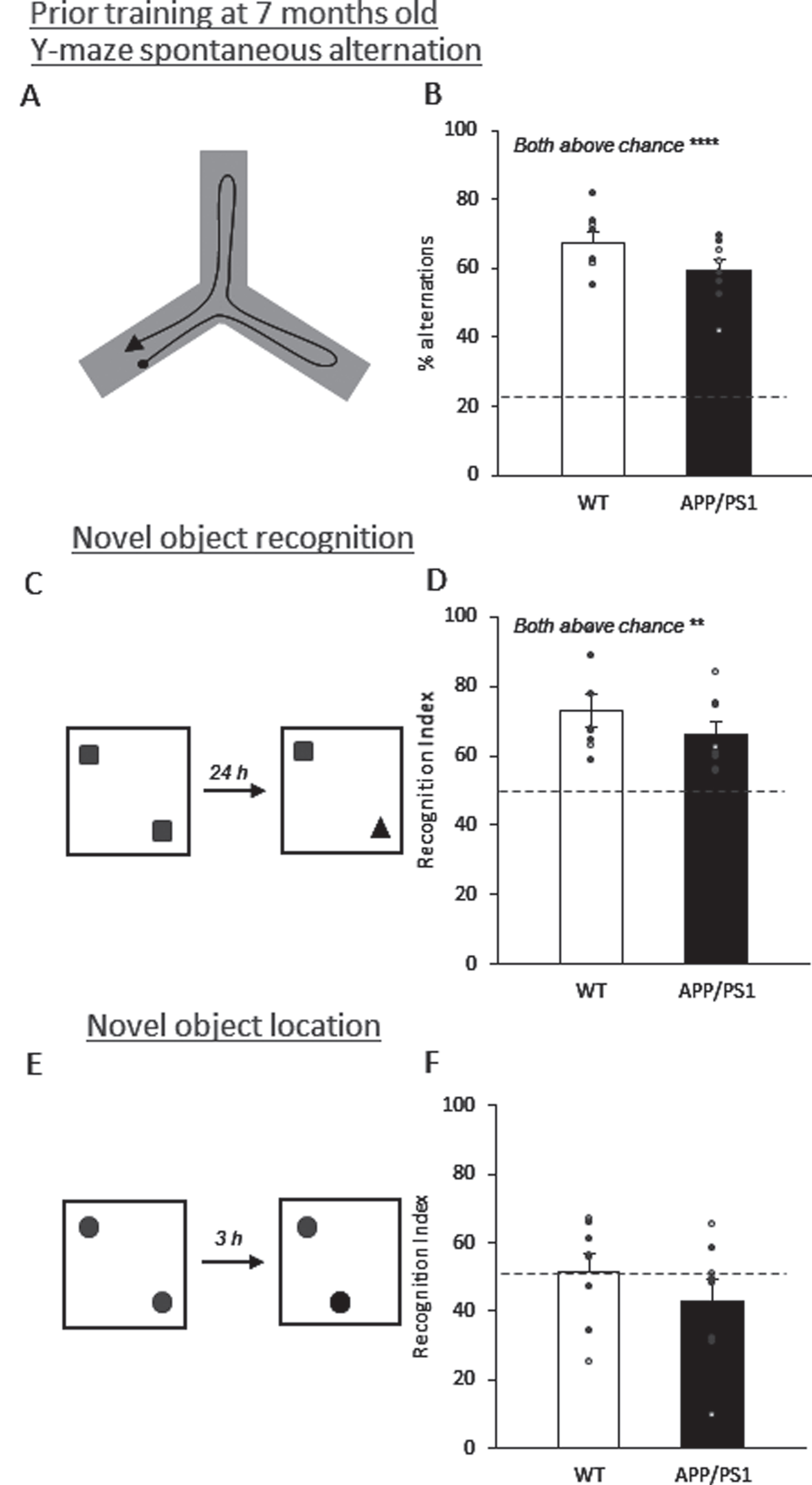
APP/PS1 mice performed indifferent from wildtype mice in the Y-maze, NOR, or NOL tasks during the prior training at 7 months of age. A) Schematic diagram of the Y-maze with an example path (black arrow). B) Both groups performed significantly above chance (22%) in the Y-maze. There was no significant genotype effect. C) Schematic diagram of the novel object recognition box with objects for encoding (left) and test (right). D) Both groups performed significantly above chance (50%) in the novel object recognition task. There was no significant genotype effect. E) Schematic diagram of the novel object location box with objects for encoding (left) and test (right). F) Both groups performed insignificantly above chance (50%) in the novel object location task. There was no significant genotype effect. Data are presented as mean±SEM.*p < 0.05,**p < 0.01,*** p < 0.005, **** p < 0.001.
APP/PS1 generally performed similarly to wildtype mice at 7 months in the watermaze task (Fig. 7A). No significant effect of genotype was found in the first or last training blocks (Fig. 7B-G; all t14 < 1.54, p > 0.15). When mice underwent reversal and transfer training (Fig. 7H-K), there were no significant genotype effects (both t14 < 1.94, p > 0.07). There were no significant effects in the reversal training (Fig. 7H, I; t14 = –1.07, p = 0.30) or the transfer training (Fig. 7J, K; t14 = –1.94, p = 0.08). Six probe trials were carried out for short-term and long-term tests after training, reversal training and transfer training, and no significant difference between genotype was observed (Fig. 7L; all t14 < 1.61, p > 0.13). All performances in the probes were above the level of chance (all t7 > 2.54, p < 0.038).
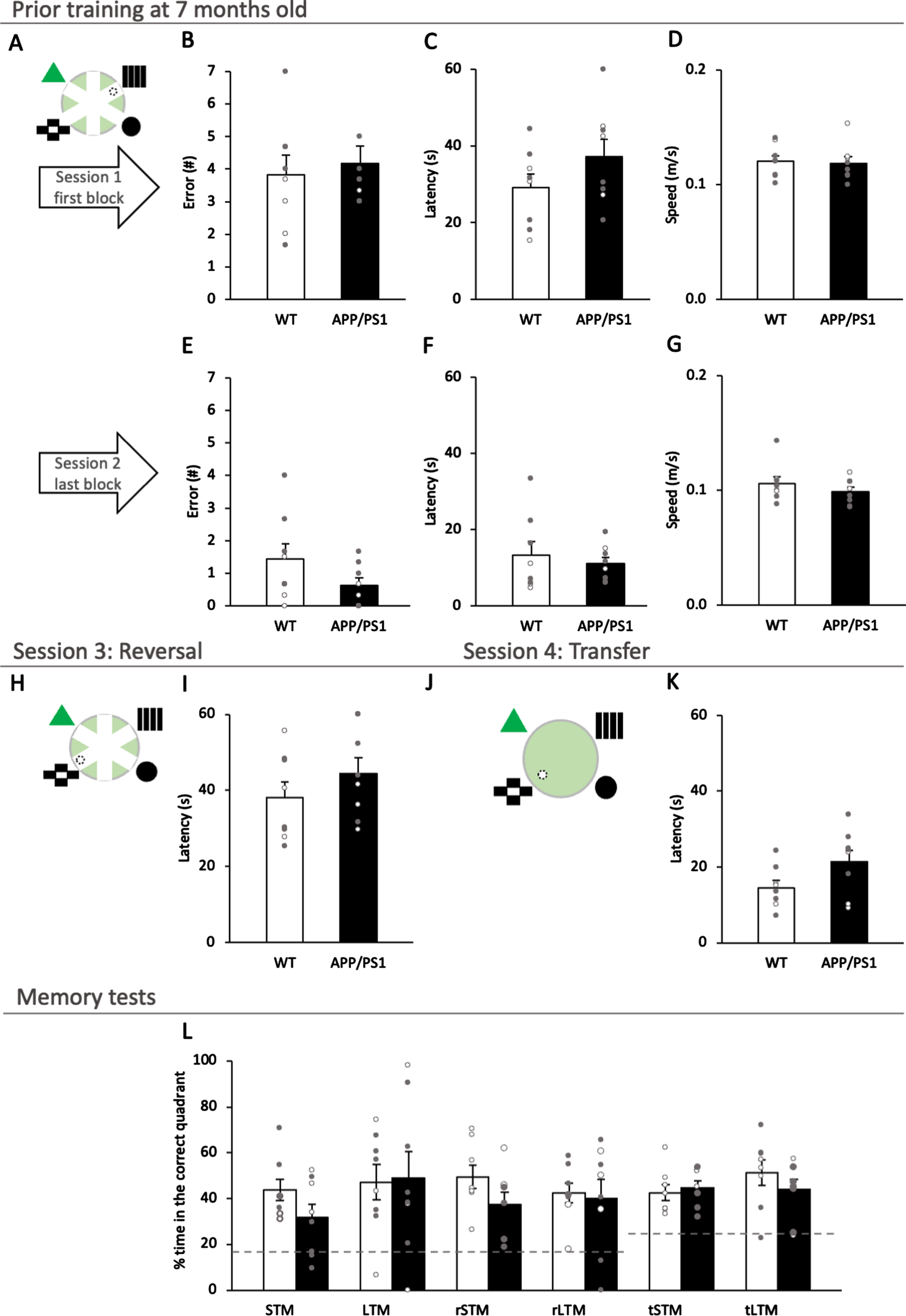
APP/PS1 mice performed indifferent from wildtype mice in the water maze tasks during the prior training at 7 months of age. A) Schematic diagram of the RAWM, showing platform location (dashed circle) and surrounding visuospatial cues in geometric shapes. B-G) There was no significant effect of genotype for either error, latency, or speed for both first and last training blocks in the RAWM. H) Schematic diagram of the reversed RAWM, showing platform location (dashed circle) moved to the opposite arm. I) There was no significant effect of genotype for latency to reach the platform in the reversal training. J) A schematic diagram of the open field water maze transfer task with platform location (dashed circle) and surrounding visuospatial cues. K) There was no significant effect of genotype for latency to reach the platform in the transfer training. L) Probe trials were carried out for short-term (STM) and long-term (LTM) memory after training, reversal training (rS/LTM), and transfer training (tS/LTM), all of which showed no significant difference between genotypes. Data are presented as mean±SEM. * p < 0.05, ** p < 0.01, *** p < 0.005, **** p < 0.001.
Gender effects
A chi-squared test was run to determine gender imbalance, and this effect was not significant (χ21,31 = 0.0026, p = 0.96). T-tests were used to determine any gender difference in the behavioral measurements. No gender effect was seen in all measurements except two. Both occurred at the very beginning of the RAWM task (session 1, block 1 of Fig. 2A and Fig. 2E), in which female making less errors (t29 = –2.99, p = 0.006) and were quicker (t29 = –3.01, p = 0.005) at finding the platform. However, this gender difference did not interact with prior training (F1,27 = 0.44, p = 0.51 for errors, F1,27 = 0.12, p = 0.73 for latencies), nor with genotypes (F1,27 = 0.39, p = 0.54 for errors, F1,27 = 0.04, p = 0.85 for latencies). Hence, the gender effect unlikely contributes to insignificant prior training or genotype effects in Fig. 2A or significant effects of these factors in Fig. 2E.
DISCUSSION
This study investigated which aspects of cognitive functions benefit from mid-adulthood prior training in AD and wildtype animals. We found that in the RAWM task, prior training improved accuracy in spatial training in AD mice, and improved efficiency in the task in both genotypes. Prior training enabled effective transfer from RAWM to open water maze. Short-term or long-term memory in the spatial, reversal, and transfer tasks were intact after sufficient training had taken place. No genotype effect was seen in spontaneous alternation, object recognition, or location recognition. Prior training did not reduce hippocampal amyloid load or astrogliosis. With all animals considered, there were significant correlations between hippocampal amyloid-β pathology and indices in the spatial water maze task.
Spatial learning and memory in APP/PS1 mice
People living with AD often experience difficulties with navigation and deficit in learning visuo-spatial associations [30, 31]. In genetic AD mouse models, poorer spatial learning and memory are often reported [27]. Using an open field water maze, APP/PS1 mice show an impairment in the acquisition of the spatial reference task [27, 33–38]. Here, we show that APP/PS1 mice making more errors and taking longer to find the platform. These significant genotype effects are unlikely due to change in motor ability, as the swimming speed is comparable between groups. This is in agreement with several findings [39, 40].
Some studies show impairment in memory retention in the APP/PS1 mice [41, 42], while our results do not support this. It is conceivable that memory retains after sufficient learning has taken place. Several factors can contribute to milder impairment seen in the current study. First, animals in this study are intensively handled and accustomed to the experimenters before the commencement of the behavioral procedures. Handling can ameliorate the anxiety level of the animals and improve cognitive performance [43, 44]. For example, C57BL/6 mice that were handled prior to water maze training show improved latency to find the platform and less variability [43]. Second, our training protocol may lead to stronger learning. This is evident in animals making minimal errors in finding the platform towards the later phase of training. It is also evident by a high proportion of time spent in the target arm (nearly 60%) in the non-reinforced memory tests. Third, wildtype littermates are used in this study. This rules out between-cohort differences, due to breeding background, and other environmental differences [45, 46].
From the current study, the poorer performance in the training phase and the transfer phase in the APP/PS1 mice may imply that the mechanisms involved in encoding are impaired. Encoding in the water maze, or other open field mazes, requires neural transmission through glutamatergic N-Methyl-D-aspartate receptors (NMDAr) [9, 48]. Toxic amyloid-β oligomers alter neurotransmitter release, glutamatergic receptor internalization, and inhibit long-term potentiation [49–51]. The affected NMDAr-related mechanism could be one of the mechanisms underlying the poor performance in AD mice in this study. As both short-term and long-term memories in APP/PS1 mice remain largely intact after initial, reversal, and transfer learning, this suggests that mechanisms involved in memory retrieval would likely be functional. As spatial learning requires the hippocampus [52–55], it is likely that hippocampal dysfunction is associated with impairment in the APP/PS1 mice [56]. The prefrontal cortex is required for reversal learning [57]. Intact reversal learning here would suggest that the prefrontal circuit remains functional in APP/PS1 mice at this age. The frontal circuit [58], related connectivity, and function [59] may become affected at later stages of AD. One limitation of our study is the use of a single AD mouse model. APP/PS1 (line 85) mice model amyloidosis, show synapse loss at 4 months old [60], synapse reduction and abolished LTP at both 6- and 12 months [61–63] and plaque-associated neuronal loss at 8–10 months old [64], and in old age at 16 months [65]. This line is limited by overexpression of AβPP and does not model tau pathology involved with overexpression of AβPP.
The benefit of prior training
Prior training reduces latencies in finding the platform from early training. This effect persists at later training and at transfer training. These support a role of prior training, albeit in mid-adulthood, in improving efficiency of performing the spatial task. Complexity of training has been shown to be beneficial for improving learning capacity [12]. It is possible that the intensity and diversity of training protocols developed in this study enables the animals to acquire effective navigating strategy that can be applied at later training. Contrasting to a recent study showing reduced flexibility in mice with cerebral β-amyloidosis using one prior training task [66], here we find that a mixture of tasks in prior training would contribute to effective subsequent learning.
Prior training can change the receptor and circuit mechanisms for subsequent learning [9, 67–70]. Whether there is a change of the underlying receptor mechanisms [67] between prior and subsequent training in midlife in the current study requires further investigation. Similarity between prior training and subsequent training in the open field water maze and in contextual fear conditioning is critical for detecting a change of circuit or receptor mechanisms in learning [9, 70]. This would imply that with common regimes in the prior training and subsequent training in the current study, the subsequent learning may become NMDAr-independent, which would warrant further investigation. It is also yet to address systemically whether similarity in training regimes or in training environment is important for the benefits observed here.
Prior training in this study also reduces errors in the AD mice in the spatial task. Environmental enrichment improves the performance of APPswe, PS1dE9, and APP/PS1 mice in spatial learning [71]. Physical activities through long-term treadmill training improve contextual fear memories in the APP/PS1 mice [72]. Environmental enrichment or exercise typically involves months of exposure at an earlier stage [71, 72] to show effects. Our approach would provide a step closer to modelling humans where prior experience involves more cognitively demanding learning, and a closer match to education and experience concepts in cognitive reserve in human studies [73]. We do recognize the limitation in extrapolating findings from rodents to humans [74]. Whether the exercising component or environmental enrichment component in the prior training will be sufficient for the benefits observed here will require future investigation. As the duration of physical exercise or environmental enrichment in our study is much shorter than the exercise or enrichment duration used to report benefits [75–78], it is conceivable that the cognitive aspect of prior training in our study plays a critical role. Here, we used overlapping tasks in prior training and subsequent training and tests. It is yet to address if completely different types of prior training that have minimal overlapping with subsequent cognitive assessment will be sufficient to provide benefits.
Intact working memory and recognition in APP/PS1 mice
One study shows impairment in spontaneous alternation in the Y-maze in APP/PS1 mice [79]. However, we have not observed such impairment and our finding is consistent with other studies [80, 81]. It is possible that after intensive handling in our study, the stress level is reduced [44] such that it rescues memory impairment in the spontaneous alternation task. It is important to note that the chance level of serial spontaneous alternation is 22.22% and a performance of 50–60% with small variance is highly significantly above chance. Even in some studies where AD mouse models showed poorer performance than the wildtype animals, the performance in AD mice is often significantly above chance [82].
The spontaneous alternation is proposed to reflect working memory and shows deficit in early phases of certain mouse models of AD [27]. Working memory requires the prefrontal cortex [57] and increasing prefrontal cortex function improves spontaneous alternation [83, 84]. With this notion, it again supports the view that 9-month-old APP/PS1 mice show intact prefrontal function.
Novel object recognition is apparent in both genotypes at 7 months of age, which is consistent with previous studies [85, 86]. Due to an age-dependent memory decline in this task at 9 months, no genotype or prior training effects are detected. It is likely that stronger training is needed for revealing the long-term recognition memory [87]. The same sampling duration that enables object recognition memory is insufficient for forming and/or retaining object location memory. The lack of significant memory in the wildtype animals limits the detection of genotype and prior training effects. Longer or multiple sampling phase or shorter testing delay is needed for revealing object location memory [34, 63].
Hippocampal pathology is unaffected by prior training
Prior training benefits efficiency and accuracy in the RAWM task but does not change amyloid-β-positive areas and amyloid-β plaque counts in the hippocampus. This is inconsistent with exercise studies that show amelioration of amyloidosis in AD mice. In APP/PS1 mice, prolonged running in young leading to reductions in amyloid-β plaque deposition [75, 89]. These effects are also seen in Tg2576 mice [90, 91]. Similarly, environmental enrichment in early life reduces amyloid-β burden in AD11, cDKO, and PS1/PDAPP mice [77, 92; but see 71, 93]. Late intervention using long-term environmental enrichment, also did not confer a benefit to amyloid-β burden in APP/PS1 mice [94]. Crucially, environmental enrichment alone is shown insufficient to confer a benefit of reducing amyloid load without further cognitive stimulation [92]. One study reports that young APP/PS1 mice show higher hippocampal GFAP coverage than wildtype mice, while older APP/PS1 mice show lower GFAP coverage than wildtype mice [95]. In our study, hippocampal GFAP is comparable across genotype with or without prior training. There are a few factors that contribute to a lack of benefits in brain pathology in this study. First, as described above, our study focused on cognitive training and the duration of exposure, compared to exercise or enrichment, is much shorter. It is possible that longer training will exert benefits. Second, we focused on addressing the research gap of mid-adulthood training. It is conceivable that the magnitude of benefit from prior training may reduce with age. Prior training may need to occur at an earlier stage of AD to delay pathology development [96]. Third, we focused on the hippocampus for its role in spatial cognition. It is possible that benefits may occur in other brain regions. Finally, we only used two markers as proxy of the pathology. Soluble oligomeric amyloid-β and synaptic loss are proposed to underpin cognitive dysfunction in AD [50, 51]. Future studies are required to investigate if prior training in mid-adulthood can ameliorate these.
In conclusion, we show a clear, albeit transient, cognitive benefit from prior training without a significant change in brain pathology in AD mice. Modelling cognitive deficits in AD animals with prior training experience can inform to what extent the cognitive stimulation during midlife can be beneficial and shorten the translational gap between studies from experience-deprived lab animals and studies from experience-enriched humans.
Footnotes
ACKNOWLEDGMENTS
We would like to thank BVS, University of Edinburgh for animal care and Ms. B. Goiatti McMahon for technical support.
For the purpose of open access, the author has applied a CC-BY public copyright license to any Author Accepted Manuscript version arising from this submission.
FUNDING
This work was supported by the Biotechnology and Biological Sciences Research Council (BBSRC Research Grant BB/M025128/1 to S-H.W.) and Alzheimer’s Research UK (Senior Research Fellowship to S-H.W.).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The datasets generated and analyzed during the current study are available from the corresponding author on request.