
Abstract
Background:
Oxidative stress plays a major role in the progression of Alzheimer’s disease (AD)-related cognitive deficits.
Objective:
This study was done to determine the protective effects of coenzyme Q10 (CoQ10) and high-intensity interval training (HIIT) alone and in combination for eight continuous weeks, on oxidative status, cognitive functions, and histological changes in the hippocampus in amyloid-β (Aβ)-induced AD rats.
Methods:
Ninety male Wistar rats were randomly assigned to the sham, control, Q10 (50 mg/kg of CoQ10; P.O.), HIIT (high intensity: 4 min running at 85–90% VO2max, low intensity: 3 min running at 50–60% VO2max), Q10 + HIIT, AD, AD+Q10, AD+HIIT, and AD+Q10 + HIIT groups.
Results:
The results showed that Aβ injection reduced cognitive functions in the Morris water maze (MWM) test and recognition memory in the novel object recognition test (NORT), which was accompanied by a decrease in total thiol groups, catalase, and glutathione peroxidase activities, an increase in malondialdehyde levels, and neuronal loss in the hippocampus. Interestingly, pretreatment with CoQ10, HIIT, or both, could markedly improve the oxidative status and cognitive decline in the MWM and NOR tests, and hinder neuronal loss in the hippocampus of Aβ-induced AD rats.
Conclusion:
Therefore, a combination of CoQ10 and HIIT can improve Aβ-related cognitive deficits, probably through an amelioration in hippocampal oxidative status and prevention of neuronal loss.
Keywords
INTRODUCTION
Alzheimer’s disease (AD), as the commonest cause of dementia, is an irreversible neurodegenerative disease associated with many symptoms, such as loss of memory, declined cognitive functions, depression, impaired judgment, confusion, abnormal behavior, and psychiatric problems [1, 2]. Based on the World Alzheimer’s Report (2018), 50 million people are living with dementia all over the world [3], which will increase to 152 million by 2050, three times the present rate of dementia [4]. Although the etiology of AD is unknown environmental and genetic factors, mitochondrial haplotypes, age, and sex play a role in its development [5]. The key pathological hallmarks of AD are neuronal death, deposition of extracellular amyloid-β (Aβ) plaques and neurofibrillary tangles in the intracellular environment, and the loss of synapses [2, 6, 7].
In addition, oxidative damage plays a role in the pathogenesis of AD [8, 9]. Oxidative stress is due to an imbalance between the generation and accumulation of reactive oxygen species (ROS) [10]. Oxidative damage causes lipid peroxidation (LPO), reactive carbonyl, nitration, and nucleic acid oxidation showed an increase in vulnerable neurons in AD [11]. Aβ deposition is linked to incising the production of ROS [2] and disrupted mitochondrial function [10]. Oxidative stress is an early feature of AD [9, 12], leading to Aβ deposition. It also induces the accumulation of Aβ1 - 42 [13, 14]. In contrast, Aβ impairs mitochondrial respiration [15] and induces oxidative stress [9]. Oxidative damage leads to neurological problems, such as an impairment in cognition and memory; thus, using antioxidants is effective in the prevention and treatment of AD [15].
Recently, the role of nutritional supplements and physical activity in the prevention and treatment of neurodegenerative diseases has been considered. Coenzyme Q10 (2, 3-dimethoxy-5-methyl-6-decaprenyl-1,4-benzoquinone; CoQ10) as a vitamin-like lipophilic antioxidant compound [16], can be found in vegetables and animal foods, like trout, beef, broccoli, chicken, and soybeans [17].
It is an essential cofactor of the electron transport chain found in the inner mitochondrial membrane, in which it can transport electrons from complexes I and II toward complex III to produce energy to transfer a proton to the intermembrane space [18]. CoQ10 as a potent antioxidant can prevent free radical-related oxidative damage, including lipid oxidation within the mitochondrial membrane [19, 20]. It also is served as an antioxidant through the activation and increasing the expression of mitochondrial uncoupling proteins (UCPs) [20]. UCPs have antiapoptotic and antioxidant effects [20]. The CoQ10 levels in the body decrease with aging and due to some diseases, like AD. Accordingly, CoQ10 has recently been used as a supplement [17].
Exercise training is a potential lifestyle intervention to reduce the incidence of dementia and AD [21]. Regular exercise increases hippocampal neurogenesis [22], which is linked to hippocampus-dependent spatial learning and memory function [23]. Exercise may enhance synaptic transmission and plasticity in the brain [22, 23]. Furthermore, regular exercise can reduce Aβ deposition and oxidative stress in many brain regions, which may mediate the exercise-related preventive role against cognitive decline in AD [24]. Among the many reasons for neglecting exercise, “lack of time” is often cited as an excuse. Therefore, aerobic training was designed as high-intensity interval training (HIIT) [25, 26]. HIIT is training with repeated and brief bouts of vigorous activity (nearly 85 to 90– 100% of maximal speed or maximum oxygen consumption – VO2 max) associated with periods of rest or low-intensity exercise [27]. Although exercise can be regarded as a promising intervention strategy to mitigate AD-related Aβ deposition and cognitive impairment [24], little is known about the effects of HIIT on cognition and memory.
Accordingly, we examined the effects of HIIT combined with Q10 supplementation for eight weeks before AD induction on learning and memory abilities, histological (number of neurons) changes, and the levels of glutathione peroxidase (GPx), total thiol groups (TTG), catalase (CAT), and malondialdehyde (MDA) in the hippocampus.
MATERIALS AND METHODS
Animals
Ninety male Wistar rats (eight weeks old; 180±20 g) were purchased from the animal laboratory of Hamadan University of Medical Sciences and housed in standard cages for a week before the experiments. They remained under standard laboratory conditions at 22±2°C under a light/dark cycle of 12 h and could eat and drink water and food. The research protocol was approved by the Ethics Committee of the Hamadan School of Medical Sciences (IR.UMSHA.REC.1400.459) and performed considering the instructions of the National Institutes of Health on laboratory animal care (NIH Publication 80-23, 1996).
Drugs and chemical agents
The following chemicals were used: CoQ10 and hematoxylin (Sigma-Aldrich Co., USA), Aβ 1 - 40 (Tocris Bioscience Co., United Kingdom), ketamine (Rotex Medical Co., Germany), and xylazine (Alfasan Chemical Co., Holland).
Training protocols
The training protocol (HIIT) was performed for eight weeks and five days per week for 49 min per session, which was performed on a 4-line animal treadmill (Tajhiz Gostare Omide Iranian, Iran). To make the training groups familiar with the treadmill, rats were forced to run on the device at a speed of 10 m/min, with a slope of 0° and a span of 5 min. Following familiarization, to evaluate maximal oxygen uptake (
Protocol of HIIT¥
¥Slop of the treadmill remains 0°, during the entire steps/stages of the training. €To determine the training intensity (Vo2max), the exercise capacity test was performed at weeks 0 and 4.
Intraventricular Aβ injection and neurosurgery
Following eight weeks of HIIT training and receiving CoQ10, to induce the AD model, all the rats were anesthetized intraperitoneally with a mixture of ketamine and xylazine (100 and 10 mg/kg, respectively) [31, 32] and placed in a stereotaxic device (Stoelting Co., USA). Aβ1 - 40 (100μg) was dissolved in 100μL of PBS (vehicle solution) and underwent incubation at 37°C for three days before use, resulting in the development of Aβ fibrils toxic to the nervous system [31–34]. Drilling the holes was done in the skull over the ventricular area considering the following coordinates: 2 mm lateral to the midline, 1.2 mm posterior to bregma, and 4 mm ventral to the cortex surface [35]. Aβ was injected using a Hamilton syringe (Hamilton Laboratory Products, USA) (5μg/5μl/rat) in the lateral ventricle [34]. The recovery period (seven days) of the animals was chosen based on previous studies [8, 34].
Experimental design
A schematic of the study design is displayed in Fig. 1. Following adaptation for a week, animals were randomly allocated to nine groups (n = 8) as listed below:
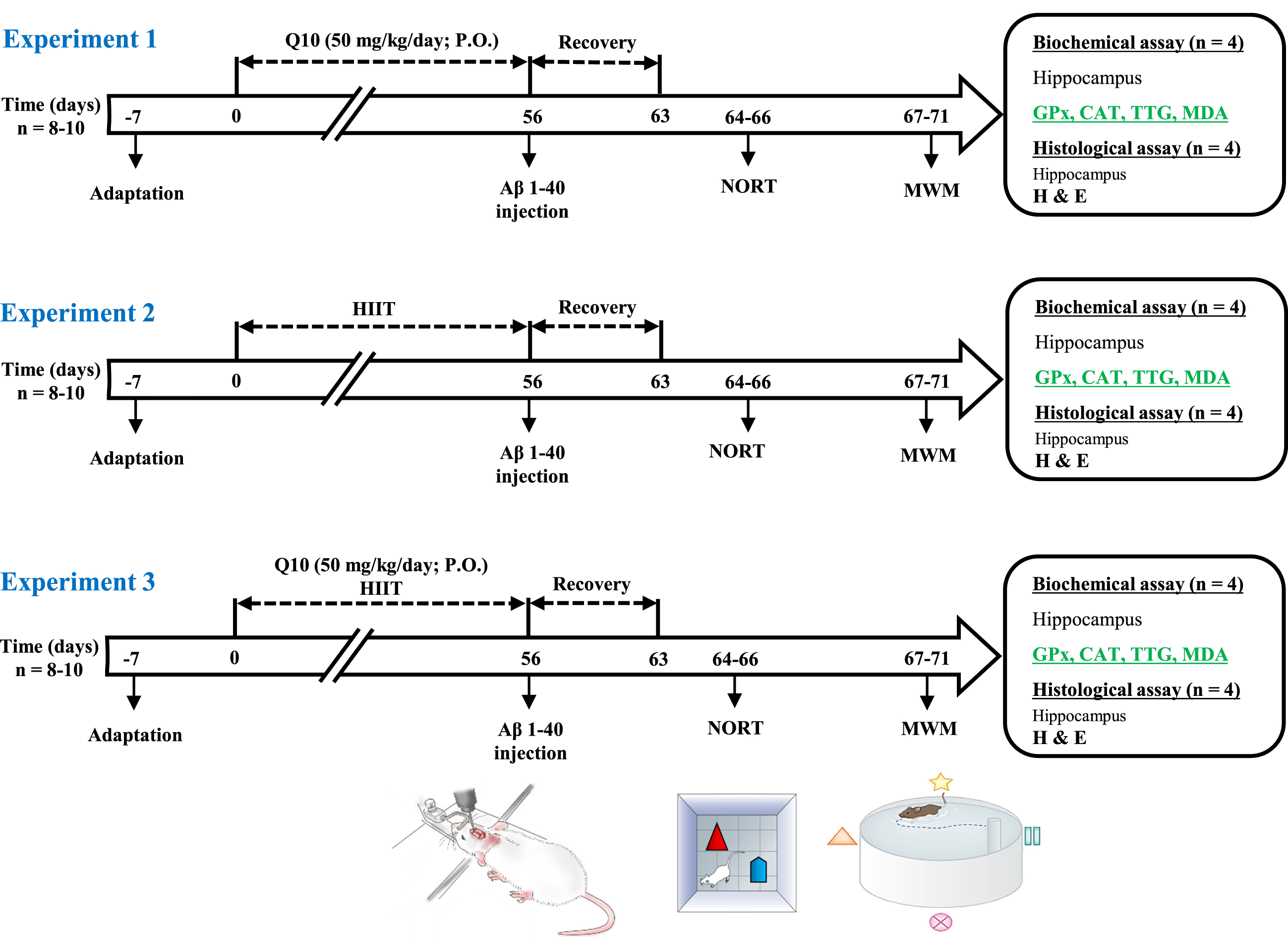
A schematic diagram of the experimental design and treatment schedule. All animals used in this study were divided into three experiments as follows: Experiment 1) After one week of adaptation, pre-treatment of rats with Q10 was initiated, and it continued for eight consecutive weeks. Then, AD was induced by Aβ1 - 40 and after recovery for one week, the novel object recognition capacity (NOR test) and spatial learning and memory (MWM task) of rats (n = 8– 10) were tested on days 64– 71 of the study. Subsequently, biochemical (n = 4) and histological (n = 4) alterations were also examined on the same day. Experiment 2) After one week of adaptation, HIIT was performed for eight consecutive weeks. Then, AD was induced by Aβ1 - 40 and after recovery for one week, the novel object recognition capacity and spatial learning and memory of rats (n = 8– 10) were tested on days 64– 71 of the study. Subsequently, biochemical (n = 4) and histological (n = 4) alterations were also examined on the same day. Experiment 3) After one week of adaptation, HIIT was performed, and also pre-treatment with Q10 was initiated, and continued for eight consecutive weeks. Then, AD was induced by Aβ1 - 40 and after recovery for one week, the novel object recognition capacity and spatial learning and memory of rats (n = 8– 10) were tested on days 64– 71 of the study. Subsequently, biochemical (n = 4) and histological (n = 4) alterations were also examined on the same day. Aβ, amyloid-β; HIIT, high-intensity interval train; NOR test, Novel object recognition test; MWM, Morris water maze; H & E, hematoxylin and eosin; CAT, catalase; GPx, glutathione peroxidase; TTG, total thiol groups; MDA, malondialdehyde; P.O., per oral.
Sham group: This group was given phosphate-buffered saline (PBS) injection (5μl/rat, intracerebroventricular (ICV).
Control group: Normal rats with free access to food and water.
Q10 group: This group received CoQ10 (50 mg/kg/day; P.O.) for eight weeks.
HIIT group: This group performed HIIT on the treadmill for eight weeks.
HIIT + Q10 group: This group performed HIIT on the treadmill and received CoQ10 (50 mg/kg/day; P.O.) for eight weeks.
AD group: This group was treated with Aβ1 - 40 (5μL/rat, ICV).
AD + Q10 group: This group was treated with CoQ10 (50 mg/kg/day; P.O.) for eight weeks, and then received Aβ1 - 40 (5μL/rat, ICV).
AD + HIIT group: This group performed HIIT on the treadmill for eight weeks, and then received Aβ1 - 40 (5μL/rat, ICV).
AD + HIIT+Q10 group: This group performed HIIT on the treadmill and was treated with CoQ10 (50 mg/kg/day; P.O.) for eight weeks, and then received Aβ1 - 40 (5μL/rat, ICV).
Novel object recognition test
NOR test investigates recognition memory according to the animal’s natural tendency to explore new objects. NOR has three sessions of habituation, training, and retention (Fig. 2B). The animals are given 10 min in the square box (50×50×36 cm3) with no object. In the training trial, two similar objects are positioned in the box’s back corner and the time spent exploring (sniffing, touching, or directing the nose toward the objects) the objects within 10 min is recorded. In the retention session, which is performed 24 h following the training session, the animal could explore one similar object and one new object for 10 min (different in color and shape). Then, object discrimination was assessed by comparing the time taken exploring the new object and the old object, and the discrimination index (DI) was determined as follows:
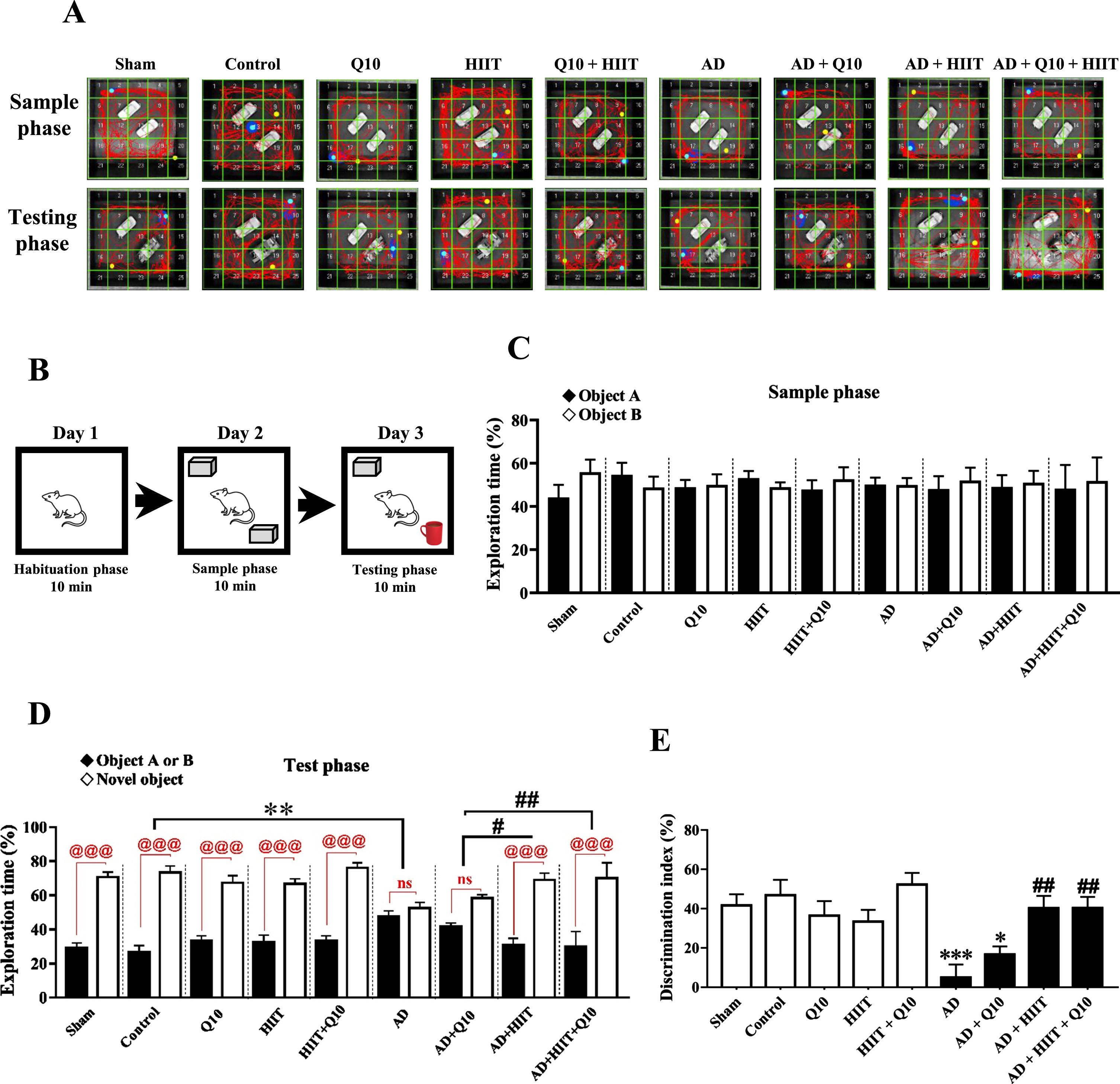
Effects of Q10 (50 mg/kg/day; P.O. for 8 consecutive weeks) and HIIT (for 8 consecutive weeks) on recognition memory by NOR test in AD rats (Aβ1 - 40, 5μL/rat, ICV). Panel A represents sample traces from rats in each group in the NOR test. Panel B shows the NOR test apparatus. Panels C-E represent exploration times and discrimination index of animals. The values were analyzed using a parametric ANOVA followed by Tukey’s post hoc test (mean±SEM, n = 8– 10). Sham group: The sham-operated rats that received PBS; Control group: The normal control rats; Q10 group: The normal control rats that received Q10; HIIT group: The rats that performed HIIT on the treadmill; HIIT+Q10 group: The rats that performed HIIT on the treadmill and received Q10; AD group: The rats that received Aβ1 - 40; AD+Q10 group: The rats that were pretreated with Q10 and then received Aβ1 - 40. AD+HIIT group: The rats that performed HIIT on the treadmill, and then received Aβ1 - 40; AD+Q10 + HIIT group: The rats that performed HIIT on the treadmill and were treated with Q10, and then received Aβ1 - 40. *p < 0.05, **p < 0.01, and ***p < 0.001 versus the control group; #p < 0.05 and ##p < 0.01 versus the AD group; @@@p < 0.001 and ns, no significant differences between groups (p > 0.05). AD, Alzheimer’s disease; HIIT, high-intensity interval training; ICV, intracerebroventricular; NOR test, Novel object recognition test.
Animals showing no exploration of the objects during testing were removed from the statistical analysis.
Assessment of spatial memory
Morris water maze (MWM) tested spatial memory [38]. It has a circular pool (diameter: 180 cm, depth: 60 cm, 22±1°C) (Fig. 3B) and four quadrants with four starting points (north (N), east (E), south (S), and west (W)) at similar distances. There is an invisible platform (diameter: 10 cm) 1 cm below the water in the northern quadrant center. Training consisted of eight trials. Each trial was terminated until the animal could find the hidden platform or after passing a maximum of 60 s. Animals that could not find the hidden platform during the considered time were guided to the platform and could remain there for 30 s. The desired parameters, such as traveled distance, time taken to reach the platform (escape latency), and speed of swimming were recorded by a video camera (Nikon, USA) installed above the pool attached to a tracking system [39]. On day five, in the probe trial, the platform was removed and animals could swim for 60 s. The time spent in the target quadrant was determined as a measure of spatial memory [40, 41].
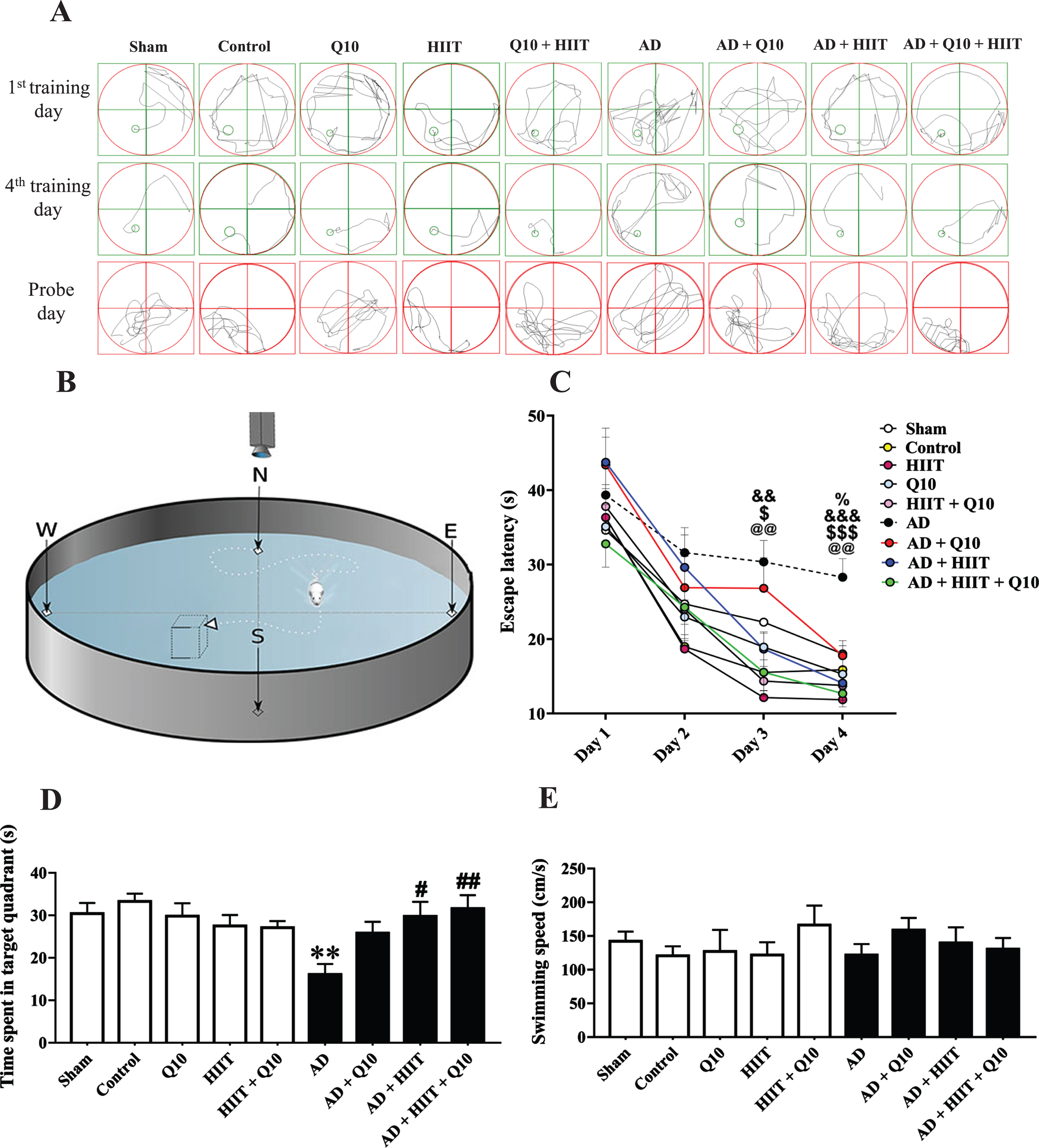
Effects of Q10 (50 mg/kg/day; P.O. for 8 consecutive weeks) and HIIT (for 8 consecutive weeks) on spatial cognitive ability using MWM in AD rats (Aβ1 - 40, 5μL/rat, ICV). Panel A shows swimming traces of rats on the 1st and 4th training days and the probe day. Panel B shows the MWM apparatus. Panel C represents escape latency to reach the hidden platform during training days. Panels D and E show the time spent in the target quadrant and swimming speed on the probe day. The values were analyzed using a two-way repeated-measures ANOVA followed by Tukey’s post hoc test (mean±SEM, n = 10). Sham group: The sham-operated rats that received PBS; Control group: The normal control rats; Q10 group: The normal control rats that received Q10; HIIT group: The rats that performed HIIT on the treadmill; HIIT+Q10 group: The rats that performed HIIT on the treadmill and received Q10; AD group: The rats that received Aβ1 - 40; AD+Q10 group: The rats that were pretreated with Q10 and then received Aβ1 - 40. AD+HIIT group: The rats that performed HIIT on the treadmill, and then received Aβ1 - 40; AD+Q10 + HIIT group: The rats that performed HIIT on the treadmill and were treated Q10, and then received Aβ1 - 40. @@p < 0.01 represents the significant difference between the AD group versus the control group; $p < 0.05 and $$p < 0.01 represent the significant difference between the AD group versus the AD+HIIT group; &&p < 0.01 and &&&p < 0.001 the AD group versus the AD+HIIT+Q10 group; %p < 0.05 the AD group versus the AD+Q10 group; **p < 0.01 versus the control group; #p < 0.05 and ##p < 0.01 versus the AD group. AD, Alzheimer’s disease; HIIT, high-intensity interval training; ICV, intracerebroventricular; MWM, Morris water maze.
Biochemical assay
At the end of the study, four rats from each group were randomly selected and anesthetized by a ketamine-xylazine mixture (100 mg/kg– 10 mg/kg; IP), and their brains were extracted from the skull, followed by removing the hippocampus and freezing at – 80°C after washing in PBS. The hippocampi of the brains were dissected on dry ice. To assess the GPx and CAT of the hippocampi, the samples were homogenized in a cold phosphate buffer (pH 7.4) and centrifuged at 12000×g for 15 min at 4°C. For MDA assessment, they were homogenized in MDA lysis buffer+BHT 100X and centrifuged at 6000×g for 10 min at 4°C and for TTG, the samples were homogenized in lysis buffer and centrifuged at 12000×g for 15 min at 4°C. Finally, based on the manufacturer’s protocols, the supernatant was removed and concentrations of the oxidative stress markers were assessed by commercial assay kits.
Histological study
After the experiments, to assess the number of intact neurons in the hippocampi, four rats were randomly selected from each group and decapitated after anesthetization by an IP injection of the ketamine-xylazine mixture (100 mg/kg– 10 mg/kg, respectively). The brains were fixed in a 10% neutral formalin solution for 72 h. Then, the brain samples were placed in a tissue processor for 21 h and washed with tap water. Ethanol dilution (70%, 80%, 90%, and 99% respectively) was used for dehydration, and then cleared in xylene and embedded in paraffin wax (60°C) until cooling. Next, samples were sectioned with a microtome to obtain sections with a thickness of 5μm and finally collected on glass slides. In the staining protocol, brain sample slides were deparaffinized in xylene (20 min), rehydrated in descending alcohol (96%, 90%, 80%, and 70%, 5 min each solution), washed in tap water, stained with hematoxylin (10 min), and washed with tap water, HCL 1%, and lithium carbonate (30 s each). Then, the slides were stained with eosin (60 s) and washed with tap water, destained in ascending alcohol solutions (70%, 80%, 90%, and 96%), and cleared with xylene. The number of intact neurons was counted in the hippocampal CA1, CA3, and DG areas. An Olympus PX 50 F3 optic microscope (Japan) was employed to observe the slides.
Statistical analysis
GraphPad Prism 5.0 was applied for data analysis and graph construction. The homogeneity of variance and normal distribution were tested by Levene’s test and the quantile-quantile (QQ) plot, respectively. Outliers were indicated considering whether the data were more than two standard deviations away from the mean. The escape latency in the MWM was evaluated using repeated measures (RM) of two-way analysis of variance (ANOVA), and the difference between the groups was checked using one-way ANOVA. A parametric ANOVA was used for other values and then Tukey’s test was performed for multiple comparisons and the values were reported as mean±standard error of the mean. p-values<0.05 were regarded as significant.
RESULTS
NOR test
The exploration time of the two similar objects was similar among the experimental rats in the sample phase (F8,78 = 0.3428; p > 0.05, Fig. 2C). In the test phase, control rats preferentially explored the new object over the familiar object (p < 0.001, Fig. 2D); nonetheless, AD animals showed a marked reduction in time spent exploring the novel object than the control group (F8,78 = 5.201; p < 0.01). Interestingly, HIIT and HIIT+Q10 fully restored the exploration time spent with the new object in the AD+HIIT (p < 0.05) and AD+HIIT+Q10 (p < 0.01) groups compared to the AD group (Fig. 2D).
The DI showed a marked reduction in the AD group than in the control group (F8,78 = 5.72; p < 0.001). In contrast, it significantly enhanced in the AD+HIIT (p < 0.05) and AD+HIIT+Q10 (p < 0.01) groups than in the AD group (Fig. 2E).
MWM test
A two-way repeated-measures ANOVA was used to evaluate the effects of treatment, day, and treatment×day interaction on the escape latency to find the submerged platform. The escape latency of animals showed the significant effects of the treatment (F8,288 = 8.018; p < 0.001), and day (F3,36 = 163.3; p < 0.001), without the significant effect of treatment×day interaction (F24,288 = 1.101; p > 0.05) in training trials.
As expected, the experimental groups could learn the platform position, evidenced by a decrease in the latency to reach the platform in the four training trials. However, according to the post hoc test, the AD group showed longer latency than the control group on the training days three (p < 0.01) and four (p < 0.01), suggesting that Aβ decreased spatial learning ability in the AD group (Fig. 3C). Surprisingly, the escape latency showed a marked reduction following treatment with Q10, HIIT, and HIIT+Q10 in the AD group on the training days three (p > 0.05, p < 0.05, p < 0.01, respectively) and four (p < 0.05, p < 0.001, and p < 0.001, respectively).
In the probe trial, the AD group showed the shortest time spent in the target quadrant than the control group (F8,78 = 3.203; p < 0.01, Fig. 3D), suggesting a reduction in spatial memory after exposure to Aβ. The AD+HIIT (p < 0.05) and AD+HIIT+Q10 (p < 0.01) groups showed a marked increase in the time spent in the target quadrant compared to the AD group.
No significant difference was detected in swimming speed among the experimental groups (F8,78 = 0.6183; p > 0.05, Fig. 3E), reflecting that all alterations obtained on the probe and training days were not due to the alterations in motor activity.
Oxidative-antioxidative status
Exposure to Aβ reduced the activity of free radical scavenging enzymes, such as GPx (F8,27 = 11.78; p < 0.001, Fig. 4A) and CAT (F8,27 = 8.507; p < 0.001, Fig. 4B) in the AD group than in the control group. The GPx activity markedly enhanced in the AD+HIIT (p < 0.05) and AD+HIIT+Q10 (p < 0.001) groups compared to the AD group. Additionally, there was no significant difference in the CAT activity among the AD+Q10, AD+HIIT, and AD+HIIT+Q10 groups compared to the AD group (p > 0.05). However, no significant difference was detected in the CAT activity among the AD+Q10, AD+HIIT, and AD+HIIT+Q10 groups compared to the control group (p > 0.05).

Effects of Q10 (50 mg/kg/day; P.O. for 8 consecutive weeks) and HIIT (for 8 consecutive weeks) on oxidative-antioxidative status assessed by the levels of GPx (A), CAT (B), TTG (C), and MDA (D) in the hippocampal tissue in AD rats (Aβ1 - 40, 5μL/rat, ICV) using assay kits. Data were analyzed by a parametric ANOVA followed by Tukey’s post hoc test (mean±SEM, n = 4). Sham group: The sham-operated rats that received PBS; Control group: The normal control rats; Q10 group: The normal control rats that received Q10; HIIT group: The rats that performed HIIT on the treadmill; HIIT+Q10 group: The rats that performed HIIT on the treadmill and received Q10; AD group: The rats that received Aβ1 - 40; AD+Q10 group: The rats that were pretreated with Q10 and then received Aβ1 - 40. AD+HIIT group: The rats that performed HIIT on the treadmill, and then received Aβ1 - 40; AD+Q10 + HIIT group: The rats that performed HIIT on the treadmill and were treated with Q10, and then received Aβ1 - 40. *p < 0.05, **p < 0.01, and ***p < 0.001 versus the control group; #p < 0.05, ##p < 0.01, and ###p < 0.001 versus the AD group. AD, Alzheimer’s disease; HIIT, high-intensity interval training; CAT, catalase; GPx, glutathione peroxidase; TTG, total thiol groups; MDA, malondialdehyde; ICV, intracerebroventricular.
TTG as a marker of non-enzymatic antioxidant status significantly decreased in the AD group compared to the control group (F8,27 = 6.553; p < 0.001, Fig. 4 C). Interestingly, TTG levels markedly enhanced in the HIIT and HIIT+Q10 groups in comparison to the AD rats (p < 0.05 for all).
The free radical damage was assessed following Aβ exposure using LPO determined by MDA levels. Tukey’s post-hoc test showed a significant elevation in MDA levels in the hippocampal area of the AD group compared to the control group (F8,27 = 5.003; p < 0.01, Fig. 4D). Also, MDA levels significantly decreased in the AD+HIIT and AD+HIIT+Q10 groups compared to the AD group (p < 0.01 and p < 0.05, respectively).
Histological results
There was no significant difference in the number of intact neurons of the CA1, CA3, and DG areas of the hippocampus among the control groups (Sham, control, Q10, HIIT, and HIIT+Q10) (p > 0.05) (Fig. 5A, B). However, it showed a significant decrease in the hippocampal CA1 (F8,27 = 14.73; p < 0.001, Fig. 5C), CA3 (F8,27 = 8.008; p < 0.001, Fig. 5D), and DG (F8,27 = 13.36; p < 0.001, Fig. 5E) areas of the AD group than in the control rats, reflecting the neuronal loss in the Aβ-exposed rats. In similar experimental conditions, Q10, HIIT, and HIIT+Q10 markedly protected the hippocampal CA1 (p < 0.01, p < 0.001, and p < 0.001, respectively), CA3 (p > 0.05, p < 0.01, and p < 0.01, respectively), and DG (p < 0.001 for all) neurons compared to the AD group.

Effects of Q10 (50 mg/kg/day; P.O. for 8 consecutive weeks) and HIIT (for 8 consecutive weeks) on histological changes in the hippocampal CA1, CA3, and DG areas (H & E staining, scale bar 20μm) in AD rats (Aβ1 - 40, 5μL/rat, ICV). Panel A represents a photomicrograph of the hippocampal CA1, CA3, and DG regions of the rats. Panel B represents the revealing photographs of intact neurons (clear cells with distinct round nuclei, identified by black arrows) and dark neurons (shrinkage of cells with pyknotic nuclei, identified by yellow arrows). The quantitative results of the number of intact neurons are shown in the hippocampal CA1 (C), CA3 (D), and DG (E) regions. Values were obtained at a magnification of 100×from three or four sections per rat. Data were analyzed by a parametric ANOVA followed by Tukey’s post hoc test (mean±SEM, n = 4). Sham group: The sham-operated rats that received PBS; Control group: The normal control rats; Q10 group: The normal control rats that received Q10; HIIT group: The rats that performed HIIT on the treadmill; HIIT+Q10 group: The rats that performed HIIT on the treadmill and received Q10; AD group: The rats that received Aβ1 - 40; AD+Q10 group: The rats that were pretreated with Q10 and then received Aβ1 - 40. AD+HIIT group: The rats that performed HIIT on the treadmill, and then received Aβ1 - 40; AD+Q10 + HIIT group: The rats that performed HIIT on the treadmill and were treated Q10, and then received Aβ1 - 40. ***p < 0.001 versus the control group; ##p < 0.01 and ###p < 0.001 versus the AD group. AD, Alzheimer’s disease; CA1, Cornu ammonis 1; CA3, Cornu ammonis 3; DG, dentate gyrus; HIIT, high-intensity interval training; H & E, hematoxylin and eosin.
DISCUSSION
The main objective of our study was the assessment of the protective effects of CoQ10 and HIIT on spatial memory, recognition memory, oxidative status, and neuronal loss in the hippocampus. We found that an ICV injection of Aβ led to impaired spatial and recognition memory in rats, which was associated with hippocampal apoptosis and oxidative stress, and neuronal loss of the hippocampal CA1, CA3, and DG areas. Also, pretreatment with CoQ10 and HIIT for eight consecutive weeks could ameliorate spatial and cognitive memory impairment, improve the hippocampal oxidative status, and prevent the apoptosis and neuronal loss of the hippocampal CA1, CA3, and DG areas in the Aβ-injected rats.
Aβ peptide is considered a crucial factor in the production and progress of AD [42, 43]. In our study, ICV injection of Aβ was correlated with cognitive decline in the MWM task because there was an increase in the escape latency to reach the hidden platform in the AD rats during four continuous training days. Moreover, in the probe phase of the MWM test, the time spent in the target quadrant by the AD rats showed a significant reduction, indicating impaired spatial memory. These results confirm previous studies [44–46]. Interestingly, the positive effects of pretreatment on MWM parameters in the Q10, HIIT, and Q10 + HIIT groups were confirmed by a decrease in the escape latency to reach the platform and an increase in the time elapsed in the target quadrant in the AD animals, which is in line with previous reports [9, 31, 32, 46]. In line with these results, Gholipour et al. [32] revealed a decrease in the latency and distance in MWM in the Aβ-injected rats by treadmill exercise, suggesting that exercise improves Aβ-related memory impairment. Singh et al. [46] also reported that Aβ1 - 42 administration caused a significant impairment in cognitive performance in the MWM test, and treatment with CoQ10 and minocycline alone for 21 days could significantly improve spatial memory evidenced by decreased transfer latency and increased time elapsed in the target quadrant.
Because of the limitations of the MWM (the stress due to cold water can affect the findings, mice are less suited for this task than rats, assessing only spatial memory, etc.), the NOR test was applied as a second measure of cognitive performance. Our findings showed that the ICV injection of Aβ decreased DI in the NOR test. Conversely, pretreatment with Q10, HIIT, and Q10 + HIIT improved recognition memory by increased DI in AD rats. Consistent with our findings, Gholipour et al. [29] showed that treatment of Aβ-induced AD rats with both oral ecdysterone and performing HIIT for eight weeks increased DI in the NOR test. Also, Monsef et al. [47] assessed the effect of CoQ10 on diabetic rats. They indicated that supplementation of diabetic rats with CoQ10 increased DI in the NOR task.
The mechanism underlying the effects of CoQ10 and exercise training on memory improvement is still unknown, but direct and indirect mechanisms at molecular and supramolecular levels are as follows: 1) Exercise-related changes in gray matter integrity in the brain and hippocampal volume [48], 2) Improving brain circulation [48]; for example, in several studies, exercise training improved brain circulation, especially in the hippocampus, an area important for learning and memory, 3) An increase in circulation in the brain leads to adequate pumping and oxygenation of the blood and clearance of the brain waste products, like Aβ [48, 49], 4) Increased neurogenesis; intensive exercise training produces lactate [50, 51], and then, lactate stimulates the proliferation of neurons and glial cells, particularly in the hippocampus [48, 52], 5) Increased mitochondrial biogenesis after exercise training [48], 6) Increased neurotrophic factors, like brain-derived neurotrophic factor (BDNF), irisin, and insulin-like growth factor (IGF-1) [53], which play a role in hippocampal plasticity and long-term memory [48, 54], 7) Antioxidant effects of CoQ10 and exercise and improved oxidative status [8, 29].
Oxidative stress is an important risk factor for the pathology of AD [32]. Excessive ROS production results in the LPO of cell membranes, and generates cytotoxic products, like MDA, which accordingly results in the disruption of the neuronal membrane [55, 56]. In our study, TTG, CAT, and GPx (as endogenous antioxidants) activities decreased, while MDA levels increased in the hippocampus of the Aβ-induced AD rats, which is in accordance with other studies [29, 56]. Nonetheless, pretreatment of Aβ-induced AD rats with a combination of HIIT and CoQ10 improved the hippocampal oxidative status through an increase in the TTG, CAT, and GPx activities and inhibition of the increase in MDA levels. According to our findings, Gholipour et al. [29] illustrated that HIIT exercise significantly decreased MDA levels and increased GPx activities in Aβ-induced AD rats. Also, Komaki et al. [8] showed that Aβ injection could significantly increase serum MDA levels, while Q10 supplementation could significantly reverse these parameters. Thus, Q10 supplementation and HIIT may have a preventive effect on AD-type dementia.
A decrease in the number of neurons in different areas of the brain is an important factor in the pathogenesis of AD [57]. This reduction arises from impairment of postnatal neurogenesis, inflammation, naturally occurring neuronal death, oxidative stress, and mitochondrial and synaptic dysfunction [57–59]. In our research, the histological assessment showed that ICV injection of Aβ caused morphological alterations and neuronal loss in hippocampal CA1, CA3, and DG areas, which is in line with earlier reports [29, 57, 60]. In contrast, pretreatment with Q10, HIIT, and Q10 + HIIT in Aβ-induced AD rats can effectively reduce neuronal loss and morphological changes in hippocampal CA1, CA3, and DG regions, which is in agreement with other studies [48]. Although our results showed that Q10 supplementation and HIIT exercise caused a decrease in neural loss in the hippocampus of Aβ rats, the mechanism underlying remains unclear. Possible mechanisms underlying the effects of CoQ10 and HIIT on a decrease in the neural loss in Aβ rats are as follows: 1) Exercise-related increase in neurogenesis [61]; 2) A decrease in inflammatory cytokine contents [27]; and 3) Reduced oxidative stress and increased antioxidant capacity [29]. However, molecular experiments should be done to clarify the detailed mechanisms.
To investigate the causes and mechanisms involved in various diseases, especially AD which is more common in women, and for generalizability of research findings to the whole humans, it is better to study both sexes. In addition to these findings, which were related to males, exploring these approaches in females will also be necessary and needed in the future.
Conclusion
In summary, our results showed that pretreatment with CoQ10 and HIIT through a neuroprotective effect can prevent learning and memory deficits and neuronal loss, possibly through the amelioration of oxidative status (increased TTG, CAT, and GPx levels and decreased MDA levels). Hence, a combination of CoQ10 and HIIT may be a potential factor in preventing neuronal loss and cognitive deficits.
Footnotes
ACKNOWLEDGMENTS
The authors are grateful to the staff of the Neurophysiology Research Center, Hamadan University of Medical Sciences for supporting this study.
FUNDING
This research was carried out at the Hamadan University of Medical Sciences (Grant No.: IR.UMSHA.REC.1400.459).
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
DATA AVAILABILITY
The datasets generated and/or analyzed during this study are available from the corresponding author on reasonable request.