
Abstract
Background:
Alzheimer’s disease (AD) is a neurodegenerative and progressive disorder with no cure and constant failures in clinical trials. The main AD hallmarks are amyloid-β (Aβ) plaques, neurofibrillary tangles, and neurodegeneration. However, many other events have been implicated in AD pathogenesis. Epilepsy is a common comorbidity of AD and there is important evidence indicating a bidirectional link between these two disorders. Some studies suggest that disturbed insulin signaling might play an important role in this connection.
Objective:
To understand the effects of neuronal insulin resistance in the AD-epilepsy link.
Methods:
We submitted the streptozotocin (STZ) induced rat AD Model (icv-STZ AD) to an acute acoustic stimulus (AS), a known trigger of seizures. We also assessed animals’ performance in the memory test, the Morris water maze and the neuronal activity (c-Fos protein) induced by a single audiogenic seizure in regions that express high levels of insulin receptors.
Results:
We identified significant memory impairment and seizures in 71.43% of all icv-STZ/AS rats, in contrast to 22.22% of the vehicle group. After seizures, icv-STZ/AS rats presented higher number of c-Fos immunopositive cells in hippocampal, cortical, and hypothalamic regions.
Conclusion:
STZ may facilitate seizure generation and propagation by impairment of neuronal function, especially in regions that express high levels of insulin receptors. The data presented here indicate that the icv-STZ AD model might have implications not only for AD, but also for epilepsy. Finally, impaired insulin signaling might be one of the mechanisms by which AD presents a bidirectional connection to epilepsy.
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative and progressive disorder with no cure and constant failures in clinical trials [1]. The main AD hallmarks are amyloid-β (Aβ) plaques, caused by Aβ deposition, neurofibrillary tangles generated by abnormal tau hyperphosphorylation and neurodegeneration [2]. However, many other events and molecular elements have been implicated in AD pathogenesis [3], indicating that AD is actually a multifactorial disorder [4] related to many comorbidities [5]. Although still controversial, the recent FDA approval of two Aβ-targeting therapies, aducanumab and lecanemab, has given hope to society, mostly because these drugs could modify AD progression [6]. On the other hand, the modest reduction in cognitive impairment, high costs and significant rate of adverse events have raised questions regarding their safety and efficacy [6].
Diabetes mellitus is a common comorbidity of AD [7] and disturbed insulin signaling has been frequently reported in both AD brains and experimental AD models (recently reviewed in Alves et al. [8]). Indeed, because alterations suggestive of brain insulin resistance are consistent and extensive in AD, some authors have proposed the term “type 3 diabetes” to refer to AD [9, 10]. Corroborating this hypothesis, intracerebroventricular (icv) injections of streptozotocin (STZ), a glucosamine-nitrosourea compound with selective uptake into pancreatic beta cells used to mimic diabetes in animals, also induce AD features in animal models, including brain insulin resistance [11–14]. Because of that, icv-STZ injections have been widely used to mimic AD in animal models based on the disruption of the insulin signaling pathway in the brain [13].
Another disease that has been strongly associated with AD is epilepsy [15], the third most frequent neurological disorder in people over age 65 [16], characterized mainly by a susceptibility to generate seizures [17]. Although the debate regarding this connection is recent, there is important evidence indicating a bidirectional link between these two chronic neurological disorders [18–20] that date back to fifteen years before the first case report of AD [21], when Aβ plaques were first identified in brains of epileptic patients as reviewed by Buda et al. [22]. Indeed, both Aβ and tau pathologies have been associated to cognitive impairment in epilepsy [23]. Curiously, physiological memory processes might be extremely relevant for seizure development. According to Das and Lucsak [24], in order to establish epileptic circuits, the recruitment of physiological memory mechanisms may be required [24].
On the other hand, other authors have already shown that, although cognitive deficits are frequently reported in patients with chronic drug-resistant temporal lobe epilepsy, Aβ plaques and hyperphosphorylated tau are both uncommon in the resected brain tissue of these patients, which implicates that other elements might be involved in the mechanism that drives memory impairment in epilepsy [25]. In addition, AD patients have augmented risk for developing spontaneous seizures and epilepsy [26]. Between 22% to 54% of AD patients present subclinical epileptiform activity [27]. Curiously, male sex and the presence of diabetes as a comorbidity are both associated with a higher risk of developing epilepsy in AD patients [28].
In the opposite direction, epilepsy, especially late-onset epilepsy, is also a risk factor for AD development [29]. Jones et al. [30] have demonstrated that seizures can be blocked in rodent models by targeting hyperphosphorylated tau with sodium selenate. Furthermore, recently, we have reported a partial AD-like phenotype and central insulin resistance in the Wistar Audiogenic Rat (WAR) strain, a genetic model of epilepsy [31]. Consistent with these findings, Sekar et al. [32] identified alterations in insulin signaling in the brains of genetic absence epilepsy rats from Strasbourg (GAERS), another genetic model of epilepsy. Also, Chuvakova et al. [33] identified downregulation of the insulin-like growth factor-binding protein 5 gene, a protein that modulates the regulation of the IGF signaling pathway [34], in the Krushinsky-Molodkina (KM) audiogenic rat strain. These findings suggest that brain insulin resistance might play important roles in the bidirectional link between AD and epilepsy.
Despite the description of shared pathologic events in these two neurological conditions, the mechanisms that could explain the bidirectional relationship between AD and epilepsy remain unknown. As we have proposed recently [31], disturbed insulin signaling might play an important role in this bidirectional relationship. Thus, in order to understand the effects of neuronal insulin resistance in the AD-epilepsy link, rats received icv-STZ to mimic AD and then were submitted to a single high-intensity sound stimulation (∼120 dB), a protocol known to induce audiogenic seizures in susceptible animals [35, 36] We also investigated the c-Fos-positive immediate neuronal activity after acoustically-induced seizures [37, 38] in regions that express high levels of insulin receptors, and thus, we expected them to be more affected by STZ effects.
METHODS
Animals
Experiments were performed in accordance with the National Institutes of Health (NIH) guidelines for the care and use of laboratory animals (NIH Publications No. 8023, revised 1978) and approved by local government authorities (Commission for Ethics in Animal Experimentation, of the Ribeirão Preto Medical School at the University of São Paulo - protocol numbers 119/2019 and 1114/2022R3). In this study, 29 male Sprague Dawley rats were used (350–400 g). The rats were obtained from the main breeding colony at the Campus of Ribeirão Preto of the University of São Paulo. Animals were housed in a quiet, temperature controlled (23±2°C) and humidity-controlled room with 12:12 h light/dark cycle and access to food and water ad libitum.
Experimental design
Surgical procedure
Before surgery, all rats were anesthetized with Thiopentax, sodium thiopental 3% [30 mg/kg, intraperitoneal injection (ip)] with doses of maintenance (0.1 ml) by the combination of anesthetics Ketamine (60 mg/kg, ip; Agener Union Animal Health, Embu Guaçu, SP, Brazil) and Xylazine (20 mg/kg; ip; Bayer Animal Health, São Paulo, SP, Brazil), as shown by Ahiskalioglu et al. [39]. Also, in the scalp, subcutaneous anesthetic composed of 2% lidocaine hydrochloride containing epinephrine (Astra, Naucalpan, Mexico; 5 mg/kg) was injected before incision.
For the implantation of bilateral cannulas, animals were placed on a stereotaxic frame (Model 962; David Kopf Instruments, Tujunga, CA, USA). The skin overlying the skull was cut to expose the bregma. We selected the following coordinates to access the lateral ventricles: anterior-posterior – 0.8 mm, medio-lateral 1.7 mm, and dorso-ventral – 3.5 mm relative to bregma [40]. After surgery, the animals received intramuscular veterinary pentabiotic (0.05 ml; Fort Dodge Animal Health LTDA, São Paulo, SP, Brazil). The animals remained in a recovery period of 3–5 days. In order to avoid cannula infection, the rats were housed individually for the whole protocol and had access to food and water ad libitum. No environmental enrichment was applied, and the animals were not handled before the protocol.
Intracerebroventricular injection of streptozotocin
The icv-STZ AD model was generated as described previously [41], with slight modifications. 3–5 days after cannula implantation, awake animals were gently restrained for STZ microinjection. STZ (Sigma-Aldrich® - 1.5 mg/kg) dissolved in 0.05 M citrate buffer (pH = 4.5) or vehicle were divided into two equal doses and administered bilaterally through the implanted cannulas. STZ was injected immediately after dilution by means of a 5μl syringe (Hamilton Company, Reno, NV, USA) connected to a microinjection pump (Harvard Apparatus PHD 2000, Holliston, MA, USA). The total injected volume was 4μl (2μL/ventricle) at a speed of 0.5μl/min. The icv-STZ group (n = 20) was injected with STZ, and the control group (vehicle; n = 9; initially 10, but one animal had to be excluded because of cannula infection) was injected with vehicle (0.05 M citrate buffer pH = 4.5). STZ was bilaterally injected twice, with an interval of 48 h between the injections. Therefore, each icv-STZ rat received a cumulative dose of 3 mg/kg of STZ. The unequal sample size can be explained by the inclusion of 10 additional rats in the icv-STZ group to ensure that the seizures were not related to bad drug storage or drug brand. For that purpose, we executed the same protocol testing STZ from two different brands: Acros Organics (n = 5) and Santa Cruz (n = 5). However, there was no significant difference in memory or seizure behaviors when we compared the three brands (Sigma, Acros Organics, and Santa Cruz). Therefore, the animals were pooled as a single group to facilitate data analysis and representation.
Morris Water Maze test (MWM)
Spatial learning memory was assessed by the MWM test [42], performed as described previously [31] at the 14th day after STZ or vehicle icv injections.
Briefly, the MWM test consisted of 5 sessions: 4 sessions of place navigation training with 6 trials per day. A probe trial was performed in the 4th session, immediately after the 24th training trial. One minute after the probe trial, a short-term memory test was executed. One additional trial was performed in the 5th session for evaluation of long-term memory, 24 h after the short-term test, totaling 27 attempts. Figure 1a shows a diagram of the experimental design. On the 25th trial (probe trial), performed in the 4th experimental session, the platform was removed, and the animals were monitored for exploration of the target quadrant (quadrant 1) for 60 s. During the short and long-term tests, the platform was relocated in the target quadrant and the animals had 180 s to find the escape platform. The short-term test was performed immediately after the probe trial, while the long-term test was carried out 24 h after the short-term test. The apparatus consisted of a black circular pool (150 cm in diameter, 39 cm in height) filled with water (temperature at 22±1°C), conceptually divided into four imaginary equal quadrants. A black platform (12 cm in diameter, 25 cm in height) was placed 2 cm below the surface in the center of the target quadrant. The animals were trained to find the hidden platform from different starting points using visual cues fixed in the room walls. The platform remained in quadrant 1 during all trials and tests, except for the probe trial in which the platform was removed. A time of 60 s was established for the animal to find the escape platform, except for the last 2 tests, which had a maximum duration of 180 s. The test ended when the rat found the platform and remained on it for 30 s. The animals were handled and wrapped in a towel for 1 min after each MWM session in order to dry them before returning to their cage. Rats were videotracked and their movement was quantified with Ethovision software (Noldus), which calculated the following variables: latency to the hidden platform, swimming velocity and quadrant occupation.
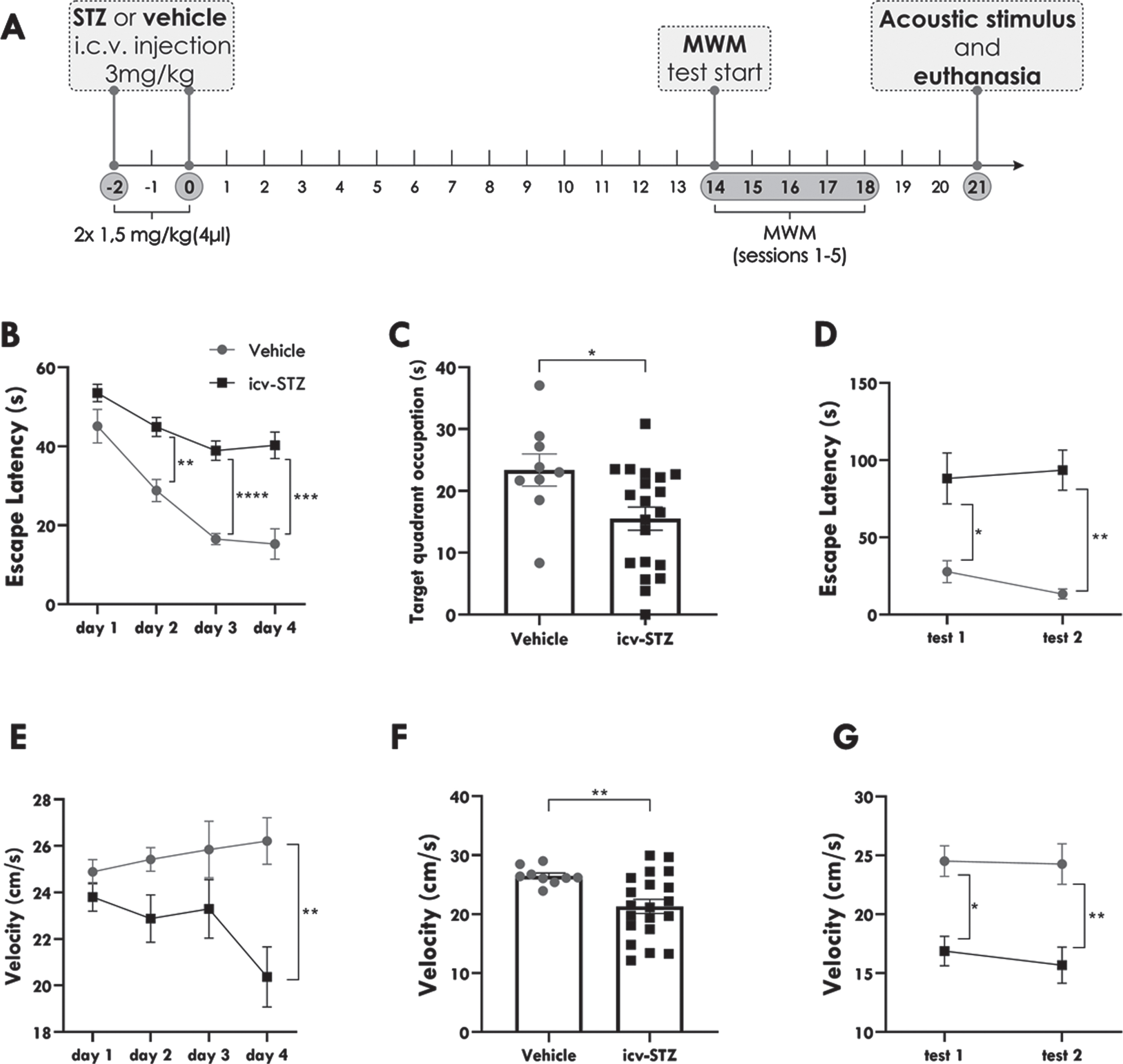
icv-STZ rats present worse performance in the MWM test. Spatial navigation performance of icv-STZ and vehicle-treated rats in the Morris Water Maze test (MWM) during the 24 trials of spatial acquisition (training period), probe trial and tests performed immediately (test 1), and 24 hours after the probe trial (test 2). A) Diagram of the experimental design. B) Latency to locate the escape platform during spatial acquisition represented by day. C) Time of target quadrant occupation during the probe trial. D) Latency to locate the escape platform during the short (test 1) and long-term tests (test 2). E) Swimming velocity to locate the escape platform during spatial acquisition represented by day. F) Swimming velocity during the probe trial. G) Swimming velocity during short and long-term tests. Data are expressed as the means±SEM (vehicle n = 9; icv-STZ n = 20). Statistical analysis was calculated by a two-way repeated-measures analysis of variance (ANOVA) followed by Sidak’s or by unpaired t test. The asterisks indicate significant differences as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Acoustic stimulation
At the 21st day after STZ or vehicle injection, icv-STZ animals (n = 14) and vehicles (n = 9) were placed in an acrylic, acoustically isolated, arena (height: 32 cm, diameter: 30 cm), located inside a soundproof chamber (45×45×40 cm). After 1 min of acclimation, they were submitted to a single high-intensity acoustic stimulus (AS-120 dB broadband sound) for 1 min or until the appearance of a tonic seizure (score 4), see details below. Animal behavior was recorded for 1 min before, during, and 1 min after AS. icv-STZ rats submitted to sound are going to be referred as icv-STZ/AS and vehicles submitted to sound are going to be referred as vehicle/AS. After stimulation, the animals were kept in the cage for one more minute to assess postictal behaviors and then returned to their home cages. Brainstem seizure severity was scored using the Categorized Severity Index (CSI) [43] and the Racine index was used for limbic seizures severity (Table 1) [44]. A control group with no sound (icv-STZ/NS) consisted of icv-STZ rats that were equally manipulated as the rats submitted to the acoustic stimulus, but the sound was not applied.
Tissue processing
For immunohistochemistry, 90 min after AS, or animal exposure to the acoustic chamber only for the icv-STZ/NS group, rats were anesthetized with sodium Thiopental (50 mg/kg; i.p.; Abbott, Brazil) and then submitted to transcardiac perfusion with 0.1 M phosphate buffer saline (PBS; pH 7.4) and paraformaldehyde solution (PFA 4%, diluted in PBS; pH 7.4). Posteriorly, their brains were removed, postfixed in PFA 4% solution for 4 h, and cryoprotected with sucrose solution (30%). Brains were frozen and cut into coronal sections (40μm thickness) using a cryostat (Microm HM-505-E, Microm International, Walldorf, Germany), and stored in PBS pH 7.4 at 4°C. Serial sections were taken using bregma references (–1.32 mm to –4.08 mm) [40] for further c-Fos immunohistochemistry aiming the hippocampus, hypothalamus, and motor and piriform cortices.
Neuroethological analyses
As mentioned previously, and shown in Table 1, brainstem and limbic seizures severity were categorized according to the CSI [43] and Racine scale [44], respectively. In order to compare behavioral seizures among the groups icv-STZ/AS, vehicle/AS, and icv-STZ/NS, a second-by-second neuroethological analysis was performed during the pre-, sound, and post-sound phases, using a glossary of behavioral items (Table 2) proposed by Garcia-Cairasco et al. [45]. This analysis accounts the frequency and duration of seizures along with the sequence and power of interactions between behavior pairs (dyads) [46, 47], allowing the creation of flowcharts with behavioral categories or clusters: wild running, tonic-clonic seizures, limbic seizures, postictal, exploratory, orofacial, grooming, and other behaviors. The software ETHOMATIC [46] was used to analyze interactions between behavior pairs (dyads) and to calculate log χ2 values. Frequency, duration, and interaction between behaviors were represented according to the flowchart calibration pattern (Fig. 3a). Different behaviors were represented by rectangles (height: frequency; width: duration) and clusters are represented by colored ellipses.
Glossary of behavioral items
Immunohistochemistry
c-Fos immunostaining was performed in free-floating sections (40μm). Tissue was washed (5 times, 5 min each) in PBS, and endogenous peroxidase activity was blocked in 2% H2O2 solution for 30 min. After washing in PBS, a PBS-Triton 0.2% solution was applied for 2 h. Then, sections were incubated overnight in a primary rabbit polyclonal antibody (1:500; sc-7202, lot. A208, Santa Cruz Biotechnology) diluted in bovine serum albumin solution (2% BSA, Amresco). On the next day, sections were washed in PBS and incubated for 2 h in a biotinylated secondary antibody anti-rabbit IgG (1:1000; BA-1000, lot. Zb0318, Vector) diluted in BSA solution. Next, tissue was reacted for 1 h with an avidin-biotin-peroxidase conjugate (1:800, VectaStain ABC kit, Vector) and washed in PBS and Tris-HCl (0.05 M; pH 7.6). Immunoreactive antigenic sites were visualized using the 3,30-diaminobenzidine (DAB) peroxidase (HRP) substrate kit with nickel (SK-4100, Vector). Nickel was used to intensifying the DAB reaction, avoiding possible confusion with small immunoreactive neurons. The sections were mounted on glass slides and coverslipped with Permount (Sigma, USA), then they were visualized and photographed in a scanning microscope (Olympus BX61VS). c-Fos immunopositive neurons (c-Fos+) were analyzed in 400× using the software ImageJ (National Institute of Mental Health, Bethesda, MD, USA). The mean number of c-Fos+ cells were calculated for each structure in every animal using the number of c-Fos+ cells of 3 rectangular standardized areas (regions of interest, ROIs, 50×50μm2 in the hippocampus and 100×100μm2 in other areas), randomly measured in each section analyzed, following [48].
Statistical analysis
For latency during spatial acquisition and tests in the MWM, a two-way repeated-measures analysis of variance (ANOVA) was performed followed by Sidak’s multiple comparisons test. For time in target quadrant during the probe trial in the MWM, unpaired t-test was used. For immunohistochemistry data, one-way ANOVA followed by Tukey’s multiple comparison test were used for comparisons among groups. Neuroethological analysis with frequency, duration, sequential behavior occurrence, and pairs of interactions between behaviors (dyads) was calculated by a Chi-Square test (χ2). Significant interactions should present χ2≥3.84, log χ2≥0.25. Comparison of binarized (presence or absence) measures for seizures between the two groups was assessed using the Fisher’s exact test, while latency to first running was analyzed by the Log-rank (Mantel-Cox) test between the groups. Seizure severity (CSI) was analyzed using unpaired t test. Quantitative analysis of the neuroethology and c-Fos expression were calculated using one-way ANOVA followed by Tukey’s multiple comparison test. Data are presented as means±SEM, and p < 0.05 was considered statistically significant. All statistical analyses were performed using Prism (GraphPad Prism 8.0 Software Inc., San Diego, CA, USA).
RESULTS
Effects of icv-STZ injections on spatial learning memory
In order to evaluate the hippocampus-dependent acquisition and short and long-term spatial memory, all animals used in this study were submitted to the MWM test [42, 49] on the 14th day after STZ (or vehicle) icv injection.
During the training trials, for the parameter latency, we observed a main effect of day, which indicates learning throughout the four days of spatial acquisition [F (2.071, 55.92) = 26.82; p < 0.0001; Fig. 1b]. We also found a significant effect of drug injection [F (1, 27) = 36.34; p < 0.0001; Fig. 1b], indicating that icv-STZ rats took significantly longer to find the escape platform than vehicle-treated animals. In the post-hoc analysis, we observed that, although icv-STZ and vehicle-treated animals presented similar latencies in the first day of training (p > 0.05), icv-STZ rats presented higher latencies to find the hidden platform in the second (p = 0.0014), third (p < 0.0001), and fourth (p = 0.0004) day of training. These data indicate that both icv-STZ and vehicle-treated rats learned to locate the escape platform throughout the 4 days of training trials. However, these two groups significantly differed in their latency to reach the platform from the second to the fourth day of spatial acquisition, suggesting worse MWM spatial learning of icv-STZ rats two weeks after STZ icv injection.
Immediately after the 24th trial, we performed a probe trial to evaluate the retrieval of spatial memory. During this trial, the escape platform was removed, and the animals were monitored for exploration of the target quadrant for 60 s. We observed a statistically significant decrease in the time (s) of occupation of the target quadrant in icv-STZ animals relative to vehicles (15.51±1.861 versus 23.38±2.599; p = 0.0236; Fig. 1c), showing that icv-STZ rats explored less the quadrant where the platform was usually located.
Then, we performed two additional trials: immediately after the probe trial (test 1) and 24 h after the first test (test 2). We found a significant effect of drug injection in the latency to locate the hidden platform during the tests [F (1,27) = 17.16; p < 0.001; Fig. 1d), but no effect of the day of testing. As observed in the training trials, icv-STZ animals took significantly longer to find the platform than the vehicles in both short (p = 0.0203) and long-term (p = 0.0017) tests, but their performance was similar in tests 1 and 2. Therefore, both short and long-term memories were impaired in icv-STZ animals.
For swimming velocity, we found a significant effect of drug injection during spatial acquisition [F (1,27) = 5.523; p = 0.0263; Fig. 1e] and tests [F (1,27) = 17.36; p = 0.0003; Fig. 1g]. In the post-hoc analysis, we observed that, during spatial acquisition, there were no significant differences in velocities between icv-STZ and vehicle-treated rats in the 3 first days of training (p > 0.05). However, icv-STZ rats presented significant lower speed in the fourth day of training (p = 0.0055). On the other hand, during the tests, icv-STZ animals swam slower than vehicles in the first (p = 0.0038; Fig. 1g) and second tests (p = 0.0011; Fig.1g). icv-STZ animals also presented decreased swimming velocity in the probe trial when compared to vehicles (21.32±1.210 versus 26.49±0.5053; p = 0.0095; Fig. 1f).
These data altogether indicate that icv-STZ rats present short and long-term memory deficits compared to vehicles. Moreover, although they presented similar swimming velocities in the first trials, icv-STZ animals swam slower than vehicles in the last day of training, probe trial, and tests.
Effects of icv-STZ injections on seizure expression
Twenty-three animals were submitted to a single AS at the 21st day after STZ (icv-STZ/AS n = 14) or vehicle injection (vehicle/AS n = 9). As shown in Table 3, 71.43% (10/14), of all icv-STZ/AS animals presented at least one seizure-related behavior (wild running), in contrast to 22.22% (2/9) of vehicles, suggesting that icv-STZ injections are associated to the manifestation of seizure-related behaviors (p = 0.0291, Fisher’s exact test; Table 3). Moreover, although none of the vehicles/AS presented tonic seizures (0.0%; 0/9), 64.29% (9/14) of icv-STZ/AS animals developed tonic seizures, indicating that icv-STZ is somehow involved in the development of more severe type of seizures (p = 0.0024, Fisher’s exact test; Table 3).
Number and percentage of animals that developed at least one seizure-related behavior (wild running) and tonic seizures
Statistical analysis was done with the Fisher’s exact test. The asterisk indicates significant difference as follows: *p < 0.05; **p < 0.01.
Regarding seizure severity, we observed that seizures were more severe in icv-STZ/AS rats in comparison to vehicles/AS (3.929±0.7447 versus 0.4444±0.3379; p < 0.01; Fig. 2a). Also, the seizures presented by icv-STZ/AS rats were positively associated with lower seizure onset latencies (χ2 = 6.221; p = 0.0126; Fig. 2b).
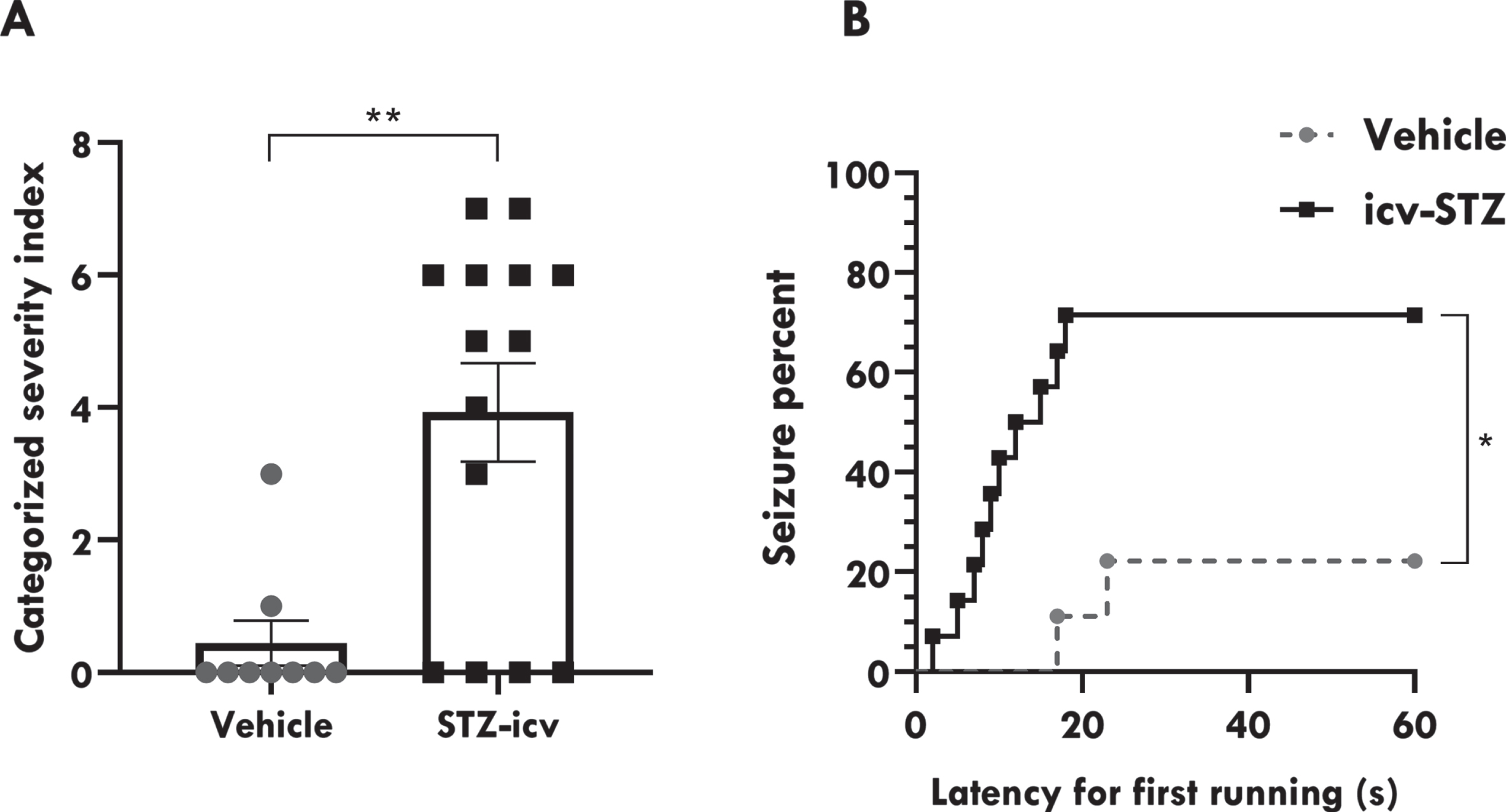
icv-STZ administration facilitates audiogenic seizures in rats. A) Seizure severity categorized according to CSI. B) Seizure percent and latency to first running during acoustic stimulation. Data are represented by mean±SEM (vehicle/AS n = 9; icv-STZ/AS n = 14). Unpaired t test compared vehicle/AS and icv-STZ/AS groups during acoustic stimulation. Latency to first running was analyzed with a Log-rank (Mantel-Cox) test. The asterisks indicate significant differences as follows: *p < 0.05; **p < 0.01.
Although some animals presented limbic seizure behaviors, those were not significant enough to be categorized according to Racine (1972) index. In fact, limbic seizures are usually displayed after chronic audiogenic seizures (audiogenic kindling) in genetically susceptible strains [44]. These behaviors will be presented in detail in the neuroethological analysis.
Neuroethological analysis of icv-STZ effects during acoustic stimulation
Neuroethological analysis (see Garcia-Cairasco et al. [45, 46]) showed complex sequences of animals’ behaviors in response to acute high-intensity sound stimulation (Fig. 3a). As can be observed in Fig. 3b, before the AS (pre-sound phase), vehicle/AS animals explored the chamber, but also presented grooming behaviors and masticatory movements (automatisms). When the sound started, vehicle/AS rats immediately expressed a startle response, significantly followed by withdrawal. Sequentially, vehicles/AS rats explored the chamber. Two vehicle/AS animals presented seizures that began with wild running (yellow symbols), gyrating, or circling behavior, followed by jumping; however, they did not progress to tonic seizures. Therefore, with the absence of tonic seizures (following Garcia-Cairasco et al. [45]), the sound continued until the stimulus completed 1 min. Only one animal from the vehicle/AS showed grooming behavior. When the sound ended, the vehicles presented freezing and grooming behaviors, except for one vehicle/AS animal that presented tachypnea after wild running behavior.
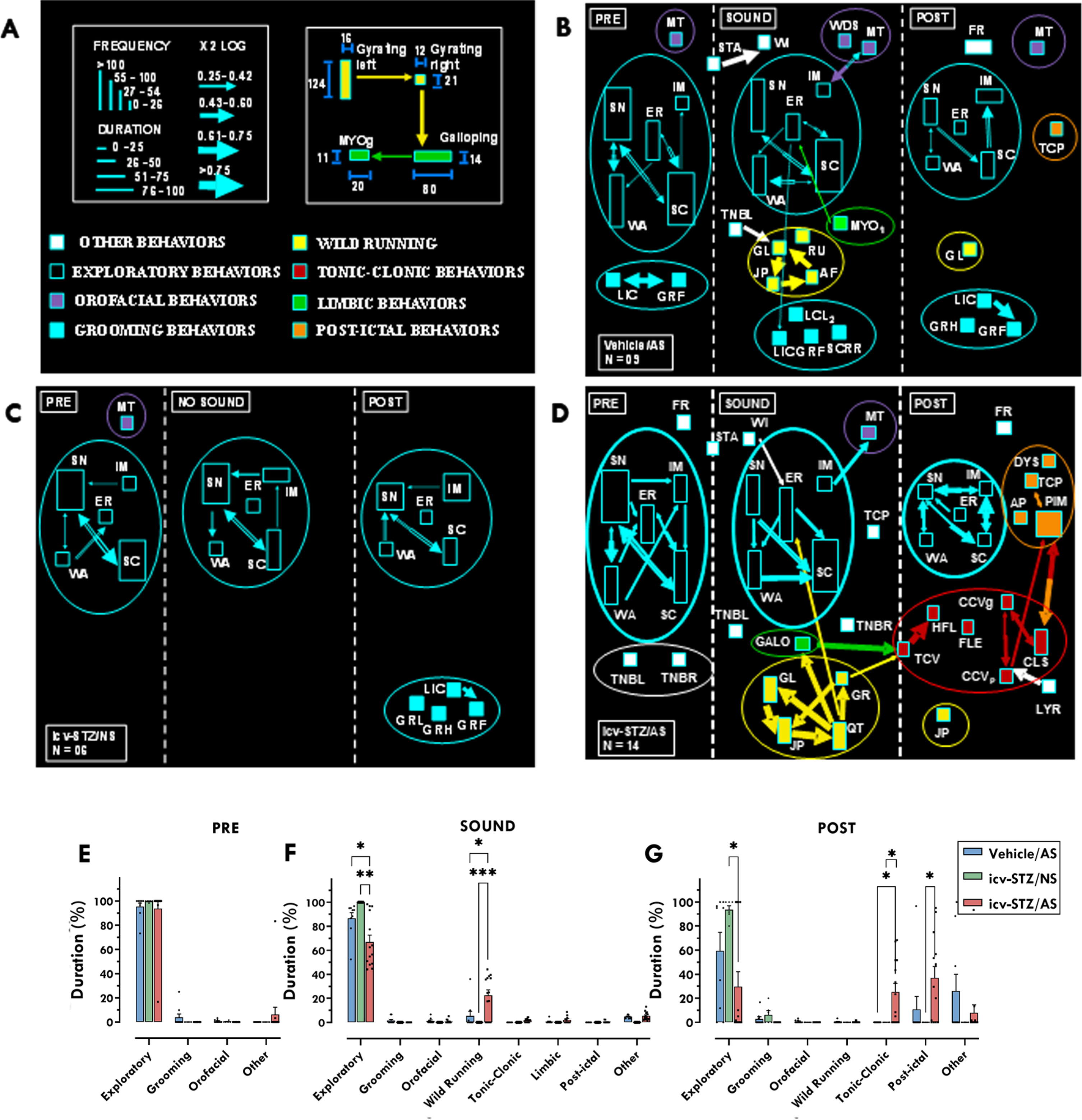
Neuroethological analysis of STZ effects during acoustic stimulus. A) Flowchart calibration pattern for neuroethological analysis, as proposed by Garcia-Cairasco et al. [45] Each group is divided into three different stages, pre-sound (1 min), sound (until tonic seizure or a maximum of 1 min), and post-sound (1 min). All behaviors are displayed as rectangles, their height corresponds to the behavior’s frequency, and their base corresponds to the mean duration over the observation period. Statistical interactions (χ2, see details below) between behavioral pairs (dyads) are proportional to the width of the arrows (χ2 values in log scale) that link them. The colors used in the rectangles are applied to qualitatively identify the clusters (or categories). The starting point of a dyad determines the color of the arrows. Therefore, any rectangle and its corresponding exiting (out) arrow must have the same color to facilitate identifying the direction of the behavioral interactions. Yellow clusters: wild running behaviors; red clusters: tonic-clonic behaviors; green clusters: limbic behaviors; orange clusters: post-ictal behaviors; purple cluster: orofacial behaviors; blue cluster: grooming behaviors; empty symbols: exploratory behaviors; white symbols: other behaviors. Complete information about each acronym and its related behavior can be found in Table 2. B) Detailed vehicle/AS behaviors before, during and after acoustic stimulus. C) Detailed icv-STZ/NS behaviors before, during and after acoustic stimulus. D) Detailed icv-STZ/AS behaviors before, during and after acoustic stimulus. E-G) Quantitative analysis of the neuroethology calculated using one-way ANOVA followed by Tukey’s multiple comparison test, before (E), during (F), and after (G) AS. Data are represented by mean±SEM (vehicle/AS n = 9; icv-STZ/AS n = 14; icv-STZ/NS = 6). Chi-Square test (χ2) was used to calculate behavior pair (dyad) interactions. Significant interactions: χ2≥3.84, log χ2≥0.25. The asterisks indicate significant differences as follows: *p < 0.05; ***p < 0.001.
On the other hand, although icv-STZ/AS rats also presented a startle response, there was no significant interaction between this response and withdrawal (Fig. 3d). Differently from the vehicle group, in the pre-sound phase, the icv-STZ/AS presented only exploratory, but no grooming or masticatory movements. In the icv-STZ/AS group, seizures started with wild running (yellow symbols) and gyri followed by jumping and atonic falling. Wild running was followed by fragmented tonic-clonic seizures behaviors (red symbols), characterized by an initial tonic seizure, followed by head ventral flexion and forelimb hyperextension, which gives rise to partial and generalized clonic seizures and clonic spasms. Six icv-STZ/AS animals presented galloping (green symbol) before tonic seizures. Only two animals presented forelimbs hyperextension, but there were no significant interactions with any other behavior. Hindlimbs hyperextension were not observed. When the audiogenic seizures ended, we noticed a strong interaction between clonic spasms and postictal immobility. Also, other postictal behaviors (orange symbols), such as apnea and tachypnea were observed. As expected, the icv-STZ animals that were not exposed to an acoustic stimulus (icv-STZ/NS) presented only exploratory behaviors during the first two minutes of recording. In the third minute, icv-STZ/NS animals presented exploration and grooming behaviors (Fig. 3c).
The quantitative neuroethological analysis showed that, during the pre-sound stage, the three groups presented similar exploratory behaviors (Fig. 3e). However, during the sound stage, a one-way ANOVA revealed that there was a statistically significant difference in the duration of exploratory behavior among the groups [F (2,26) = 9.467; p = 0.0008; Fig. 3f). Tukey’s multiple comparisons test showed that the duration of exploratory behavior was significantly lower in the icv-STZ/AS group when compared to the icv-STZ group that was not exposed to sound (icv-STZ/NS; p < 0.01; Fig. 3f) and to the vehicle group (p < 0.05; Fig. 3f).
We also observed a statistically significant difference in the duration of wild running behaviors during the sound stage among the groups [F (2,29) = 9.766; p = 0.0006; Fig. 3f]. Animals from the icv-STZ/AS group presented more wild running behaviors when compared to rats from the icv-STZ/NS group (p < 0.001; Fig. 3f) and vehicles (p < 0.05; Fig. 3f).
In the post-sound stage, there was a significant difference in the duration of exploratory behaviors among the groups [F (2,26) = 5.114; p = 0.0134; Fig. 3g]. Animals of the icv-STZ/AS group presented lower duration of exploratory behaviors when compared to icv-STZ/NS (p < 0.05; Fig. 3g), but there were no significant differences relative to vehicles/AS. On the other hand, we observed a significant difference in the duration of tonic-clonic behaviors among the groups in the post-sound stage [F (2,26) = 6.411; p = 0.0055; Fig. 3g]. The icv-STZ/AS animals presented a longer duration of tonic-clonic behaviors when compared to both icv-STZ/NS rats (p < 0.05) and vehicles/AS (p < 0.05). Regarding the post-ictal behaviors, there was a significant difference among groups [F (2,29) = 5.053; p = 0.0131; Fig. 3g). The icv-STZ/AS animals presented a significant longer duration of postictal behaviors relative to icv-STZ/NS (p < 0.05), but not to vehicles/AS.
Effects of icv-STZ injections on neuronal activation after seizures: c-Fos expression
Although the involvement of brainstem structures in acute audiogenic seizures is well described in the literature [37, 50–53], the mechanism by which icv-STZ facilitates the generation of audiogenic seizures has never been described before. In AD, the main mechanism by which icv-STZ exerts its effects on cognition is through the promotion of neuronal insulin resistance, especially in regions that express high levels of insulin receptors, such as the hippocampus, cerebral cortex, and hypothalamus [54, 55]. Therefore, our goal was to investigate the immediate early nuclear transcription factor c-Fos, a marker of neuronal activation, in these regions, which are not usually activated in a single acute audiogenic seizure [38].
One-way ANOVA showed that there is a statistically significant difference between groups in the number of c-Fos+ cells in CA1 of the dorsal hippocampus [F (2,12) = 31.87; p < 0.0001; Fig. 4a]. Tukey’s multiple comparisons test revealed that icv-STZ/AS rats presented higher number of c-Fos+ cells in CA1 when compared to both icv-STZ/NS (p < 0.0001) and vehicles/AS (p < 0.0001) rats. There was no significant difference between icv-STZ/NS and vehicles/AS. Similarly, there was also a significant difference between groups in CA3 [F (2,12) = 33.72; p < 0.0001; Fig. 4b], in which icv-STZ/AS rats presented higher number of c-Fos+ cells when compared to both icv-STZ/NS (p < 0.0001) and vehicles/AS (p < 0.0001) rats, with no significant difference between icv-STZ/NS rats and vehicles/AS. In the granular layer of dentate gyrus (DG), we also observed a significant difference between groups [F (2,12) = 13.45; p = 0.0009; Fig. 4c]. Animals from the icv-STZ/AS group showed increased number of c-Fos+ cells in DG relative to rats from the icv-STZ/NS (p < 0.001) and vehicles/AS (p < 0.05; Fig. 4c) groups.
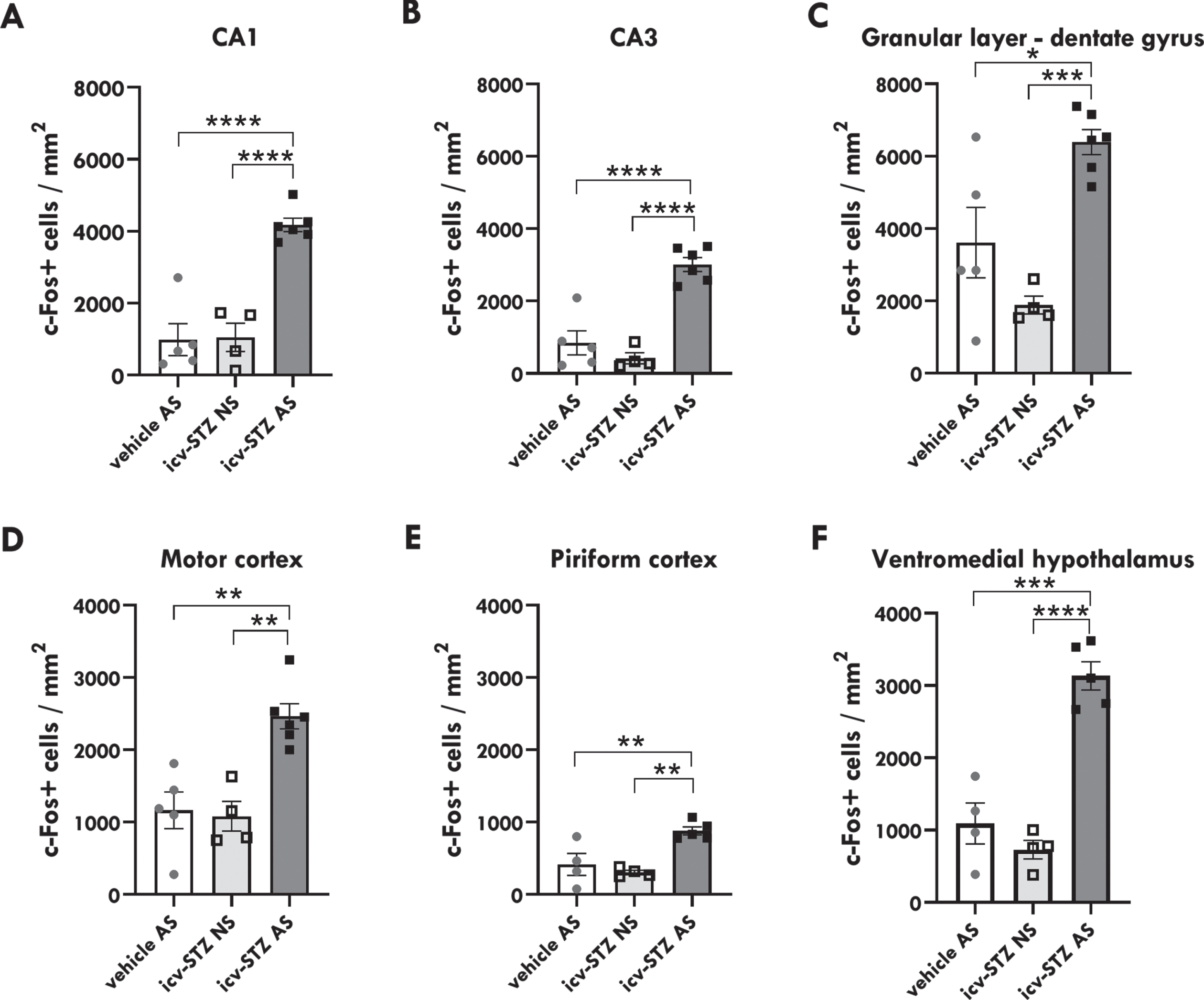
Icv-STZ/AS rats present increased number of c-Fos+ cells. A) Quantitative analysis of c-Fos+ cells per mm2 in the Cornu Ammonis 1 (CA1). B) CA3 and C) granular layer of the dentate gyrus (DG) region of the hippocampus. D) Motor cortex. E) Piriform cortex. F) Ventromedial hypothalamus. Statistical significance was calculated by one-way ANOVA followed by Tukey’s multiple comparison test. Data are represented by mean±SEM (n = 3–6/group). The asterisks indicate significant differences as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Regarding cortical regions, we identified significant differences between groups in the motor [F (2,12) = 14.33; p = 0.0007; Fig. 4d] and piriform cortices [F (2,10) = 12,41; p = 0.0020; Fig. 4e]. In the motor cortex, the icv-STZ/AS group presented an increased number of c-Fos+ cells relative to the icv-STZ/NS group (p < 0.01) and vehicles/AS (p < 0.01). Similarly, in the piriform cortex, icv-STZ/AS rats had a higher number of c-Fos+ cells when compared to both icv-STZ/NS (p < 0.01) and vehicles/AS (p < 0.01) rats.
In the ventromedial hypothalamus, there was also a significant difference between groups [F = (2,10) = 40.24; p < 0.0001; Fig. 4f), in which the icv-STZ/AS group presented higher number of c-Fos+ cells relative to the icv-STZ/NS (p < 0.0001) and vehicle/AS groups (p < 0.001). These data suggest that icv-STZ injections coupled to an acute audiogenic seizure promote increased neuronal activity in hippocampal, cortical, and hypothalamic regions (Fig. 5a). Moreover, although we focused our investigation on structures that express high levels of insulin receptors, we noticed increased neuronal activation in other brain regions (Fig. 5b), especially in brainstem structures intrinsically associated with generalized tonic-clonic seizures in audiogenic models [52] including the periaqueductal gray matter and inferior and superior colliculi [56].
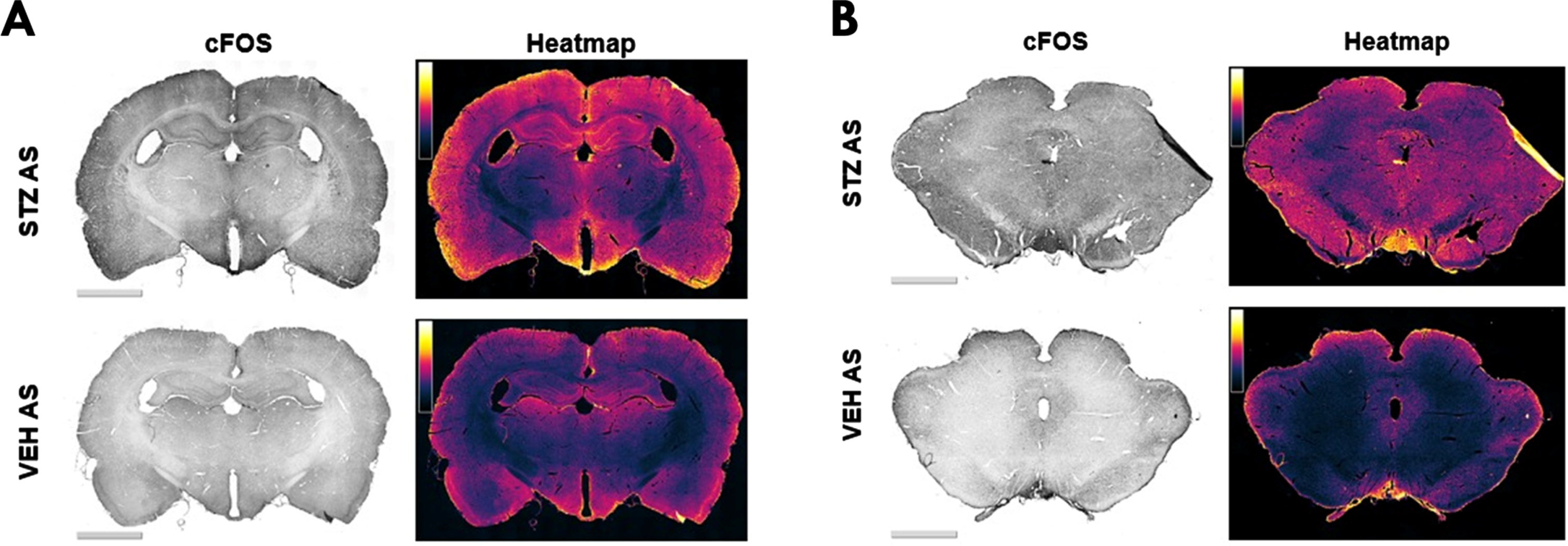
Representative images of c-Fos immunostaining in forebrain (A) and brainstem regions (B) in different experimental groups (left column) and their corresponding heatmap (right column). The color code represents the c-Fos intensity in a range of 0 (black, minimum) to 255 (white, maximum).
DISCUSSION
In this study, we have demonstrated for the first time that icv-STZ injections, usually used to mimic AD in animals [12–14, 57–59], not only promote spatial memory impairment, but also facilitate audiogenic seizures in rats submitted to a single high-intensity acoustic stimulation. Moreover, the generation of seizures in this model seems to be related to increased neuronal activity in regions that express high levels of insulin receptors, which suggests that central STZ effects on insulin signaling might have roles not only in cognitive impairment but also in seizure susceptibility with implications for both AD and epilepsy.
Although memory impairment is considered one of the main features of AD, cognitive deficits have also been frequently described in epilepsy in both clinical [60–62] and experimental studies [63–65]. In fact, recent evidence suggests that epileptogenesis requires physiological memory processes to form epileptic circuits [24]. Regarding the icv-STZ AD model, spatial memory impairment has been frequently reported in these rats [54, 66–68]. Studies have also shown that icv administration of low STZ doses promotes cognitive deficits through alterations in the regulation of the insulin signaling pathway [12, 69], which can be identified two weeks after STZ administration [13, 69].
In the current study, corroborating previous data from the literature, icv-STZ animals presented both short and long-term memory impairment when compared to vehicles two weeks after icv-STZ administration. These alterations were demonstrated by higher latencies to locate the hidden platform and decreased exploration of the target quadrant in the MWM test, which are in line with other studies that also used the MWM to investigate spatial memory in icv-STZ rats [70, 71]. Gumus et al. showed that, approximately one month after icv-STZ administration, icv-STZ rats presented memory deficits in the MWM, which were accompanied by significant alterations in Nogo-A receptor and osteocalcin gene expression. However, zinc supplementation was able to restore these alterations [72].
We also observed that, although icv-STZ animals and vehicles presented no significant differences in swimming velocity in the first three days of spatial acquisition, icv-STZ animals tended to diminish their speed over the test. Recently, we reported a very similar behavior in the WAR strain, a genetic model of epilepsy that presents a partial AD-like phenotype, including brain insulin resistance [31]. Moreover, lower swimming velocity has already been described in a transgenic AD model [73]. In addition, there is evidence that STZ does not cause impairment of motor coordination or balance [58], and usually there are no differences in spontaneous locomotion and exploratory activity when icv-STZ animals are compared to controls [74, 75]. Therefore, this parameter seems to be related to worse navigational precision, rather than to motor deficiency.
Alongside with cognitive impairment, we observed audiogenic seizures in 71.43% of the icv-STZ rats submitted to acoustic stimulation. These results are in line with clinical reports. In fact, seizures are three times more frequent in patients diagnosed with AD in comparison to healthy controls [15]. A recent meta-analysis showed that the incidence of seizures among AD patients ranges from 4.2 to 31.5 per 1000 person-years and the prevalence of seizures is higher among individuals diagnosed with early-onset AD (up to 49.5%) [26]. Although two vehicle-treated animals (22.22%) presented wild running behaviors during the stimulus, none of the animals from the vehicle group presented tonic-clonic seizures. Usually, a small percentage of naïve control Wistar rats present seizures when submitted to acoustic stimulation [76]. In this sense, Cunha et al. [77] reported mesencephalic seizures in 26% of naïve Wistar rats submitted to acoustic stimulation [77]. According to Vinogradova and Grinenkobin [78], in audiosensitive rats of Wistar and Sprague-Dawley outbred strains with low seizure susceptibility, audiogenic seizures start with one or two episodes of mild running behavior [78], which is in line with the observations of the current study. Moreover, in this study, the vehicle-treated rats were previously submitted to a surgical procedure for cannula implantation and received icv injections of citrate buffer. Therefore, both mechanical and chemical factors could influence seizure development. To properly eliminate these questions, naïve animals and rats only submitted to surgery (without vehicle injection) should have also been submitted to AS. Thus, this event may be a limitation of the study.
On the other hand, rats with high genetic susceptibility to audiogenic seizures, such as the WARs [36] and the genetically epilepsy-prone rats [79], usually present wild running followed by severe sound-induced tonic-clonic seizures [78], which are very similar to the seizures presented by icv-STZ/AS rats. However, in the present study, we noticed that icv-STZ/AS rats usually presented galloping before tonic seizures. Galloping is not a common behavior observed in acute audiogenic seizures. In fact, it usually appears as a consequence of audiogenic kindling, with repetitive audiogenic seizures and just prior to the emergence of tonic-clonic seizure behaviors [48, 80]. During the chronic audiogenic seizure protocol, known as audiogenic kindling [81], the epileptic activity spreads from the brainstem to different structures of the forebrain, including the amygdala, cortex and hippocampus of several genetically-selected audiogenic strains [35, 81].
The seizures displayed by icv-STZ/AS rats were also more severe and had lower latencies. This is the first time that audiogenic seizures were described in the icv-STZ AD model. However, other authors have already briefly mentioned sporadic spontaneous seizures in icv-STZ rats during animal experimentation in AD studies [82, 83]. In fact, in our laboratory, we have also observed sporadic spontaneous seizures in icv-STZ AD model not submitted to high-intensity sound stimulation during different protocols not related to epilepsy investigation. Unfortunately, because the spontaneous seizures were unpredictable, there is no record of these seizures and, therefore, it is not possible to properly analyze and describe them (personal communication; unpublished observations). Consistent with these findings, in an AD study, Angelova et al. (2019) mentioned sporadic motor seizures during the first month after icv-STZ administration, but since the seizures were occasional, they were also not able to represent them quantitatively [82]. Moreover, audiogenic seizures have already been investigated in transgenic AD animal models. Westmark et al. (2010) submitted the Tg2576 AD mouse model as well as a Down syndrome mouse (Ts65Dn) to a high-pitched siren (118 dB) from a personal body alarm and observed audiogenic seizures in 40–80% of them [84]. These authors also demonstrated that these seizures can be attenuated by antagonists to metabotropic glutamate receptor 5 (mGluR5) or by passive immunization with anti-Aβ antibody [84]. Interestingly, there is evidence of disturbed insulin signaling in both Tg2576 [85–87] and Ts65Dn [88] mouse models.
In order to assess the brain areas potentially involved with the generation of audiogenic seizures in icv-STZ animals, we investigated the immediate early gene c-Fos, a marker of neuronal activation. According to Simler et al. [37, 38], a single high-intensity sound stimulation induces the expression of c-Fos in the subcortical auditory nuclei, superior colliculus, and periaqueductal gray matter, but only a few labeled neurons can be observed in the forebrain. Curiously, in the present study, after a single acoustic stimulus in icv-STZ/AS animals, we observed a high number of c-Fos+ cells in hippocampal, cortical, and hypothalamic areas, those areas in which are usually detected insulin receptors [54, 69]. Differently from acute audiogenic seizures, after audiogenic kindling, epileptic neuronal activity propagates from the brainstem to forebrain and limbic structures, such as the amygdala and the perirhinal, piriform, and motor cortices [81, 89].
The piriform cortex and hippocampus are the last areas to express c-Fos after audiogenic kindling [38]. Controversially, in the current study, both areas were markedly activated in the icv-STZ rat after a single audiogenic seizure. Neurons from the hippocampus, cerebral cortex and hypothalamus are the most affected by STZ because they express high levels of insulin receptors [54, 55]. Moreover, the most common type of seizures among AD patients are focal limbic seizures [28], those in which the epileptic foci are located in limbic structures. Therefore, the hyperactivation of dysfunctional neurons in limbic areas might be related to the early appearance of galloping behavior (described above) in this model.
Curiously, evidence from the literature indicates that icv-STZ usually decreases the number of c-Fos+ cells in several regions of the brain, including the hippocampus [90], brainstem [91], temporal cortex [92], and GABAergic neurons in the parafacial zone [93] in the first four weeks after icv-STZ administration, after diverse protocols with different types of stimuli inducing c-Fos expression. Contradictorily, in the current study, a single audiogenic seizure coupled to previous icv-STZ administration increased neuronal activation in cortical, hippocampal, and hypothalamic regions, which suggest that an additional trigger might be needed to induce a tonic-clonic seizure in the icv-STZ AD rat model. On the other hand, this hypothesis contradicts the reports of spontaneous sporadic seizures in this model [82, 83]. In line with our findings, Chen et al. (2012) reported markedly increased expression of c-Fos in icv-STZ mice 6 weeks after icv injection [94]. Although there are many differences among the cited protocols, in our study, STZ might have increased the number of c-Fos+ cells, even in the absence of AS. That would contribute to understanding the contradictory fact that c-Fos levels did not differ between the vehicle/AS and icv-STZ/NS. However, to confirm this hypothesis, a group of vehicle-treated rats not submitted to sound stimulation (vehicle/NS) should have been included to differentiate c-Fos expression due to AS and STZ. Therefore, this may be another limitation of this study. Moreover, the data presented reinforces the need to understand better how both spontaneous and reflex seizures are triggered in the icv-STZ AD model, as well as to better characterize the behavioral seizures and brain areas involved with their manifestation. Those groups would also contribute to evaluate additional issues. For example, the fact that STZ is injected into the ventricle, let us expect that nearby structures can be affected and eventually lesioned by neurodegeneration. This naturally adds complexity to the interpretation of acute audiogenic seizures induced by icv-STZ in usually resistant Sprague-Dawley rats, and the consequent regional expression of c-Fos.
In that context, since STZ impairs neuron insulin signaling [12, 95], we believe that STZ somehow facilitates seizure propagation, especially to neurons that express high levels of insulin receptors. Thus, central insulin resistance might have implications not only in AD, but also in epilepsy. Moreover, impaired insulin signaling might be one of the main mechanisms by which AD is linked to epilepsy. Corroborating this hypothesis, recently, disturbed insulin signaling has been implicated in different genetic models of epilepsy: GAERS [32], WARs [31], and in the KM rat strain [33]. Moreover, a recent in vitro study demonstrated that insulin decreases epileptiform activity in rat prefrontal cortex by increasing GABAergic tonic currents [96]. Regarding clinical investigation a study showed that a diagnosis of diabetes mellitus increases the risk of developing epilepsy among AD patients [28].
In line with these findings, an experimental study demonstrated that levetiracetam, an antiepileptic drug, decreases memory impairment and hippocampal cell loss in icv-STZ rats through attenuation of oxidative damage, suppression of neuroinflammatory markers, and inhibition of tau hyperphosphorylation [97]. Regarding clinical (translational) evidence, another recent study reported that the treatment with levetiracetam ameliorated executive function and spatial memory in AD patients who presented seizures or subclinical epileptiform activity [98]. Moreover, metformin, a first-line antidiabetic drug that has been investigated in AD treatment, decreases seizure generation, delays the onset of epilepsy, and restores memory impairment in both acute and chronic models of epilepsy [99].
Therefore, although the findings regarding brain insulin resistance in epilepsy are recent and scarce, we believe that disturbed insulin signaling pathway impairs neuronal function, hence facilitates the generation and propagation of seizures.
Conclusions
Besides the induction of spatial memory impairment, we showed that STZ facilitates audiogenic seizures in usually resistant Sprague-Dawley rats submitted to a single high-intensity sound stimulation. The generation of seizures in this model seems to be related to increased neuronal activity in cortical, hippocampal, and hypothalamic areas, which usually express high levels of insulin receptors. Because STZ impairs insulin signaling, these data altogether suggest that central STZ effects on insulin signaling have implications in both cognitive impairment and seizure susceptibility.
Therefore, although the findings regarding brain insulin resistance in epilepsy are recent and scarce, we believe that STZ might facilitate seizure generation and propagation by impairing neuronal function, especially in regions expressing high insulin receptor levels. The data presented here indicate that the icv-STZ model of AD might have implications not only for AD but also for epilepsy. Impaired insulin signaling might be one of the main mechanisms by which AD presents a bidirectional relationship with epilepsy. Further studies are needed to understand these mechanisms better.
Footnotes
ACKNOWLEDGMENTS
The authors have no acknowledgments to report.
FUNDING
This research was supported by the São Paulo Research Foundation – FAPESP [grant numbers 17/21155-3, 19/02787-4, 19/05957-8, 19/16574-2, 19/00849-2, and 2021/13622-6], the National Institutes for Science and Technology (INCT) - “Translational Medicine”, FAPESP [grant numbers 14/50891-1], National Council for Scientific and Technological Development (CNPq) [grant numbers 2022-2952, 305883/2014-3 and 465458/2014-9], Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001, and Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior – PROEX-CAPES. NGC holds a CNPq 1A Research Fellowship.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The datasets analyzed during the current study are available from the corresponding authors on reasonable request.