
Abstract
Introduction:
Cardiometabolic risk factors and epigenetic patterns, increased in physically inactive individuals, are associated with an accelerated brain aging process.
Objective:
To determine whether cardiometabolic risk factors and epigenetic patterns mediate the association of physical inactivity with unfavorable brain morphology.
Methods:
We included dementia and stroke free participants from the Framingham Heart Study Third Generation and Offspring cohorts who had accelerometery and brain MRI data (n = 2,507, 53.9% women, mean age 53.9 years). We examined mediation by the 2017-revised Framingham Stroke Risk Profile (FSRP, using weights for age, cardiovascular disease, atrial fibrillation, diabetes and smoking status, antihypertension medications, and systolic blood pressure) and the homeostatic model of insulin resistance (HOMA-IR) in models of the association of physical inactivity with brain aging, adjusting for age, age-squared, sex, accelerometer wear time, cohort, time from exam-to-MRI, and season. We similarly assessed mediation by an epigenetic age-prediction algorithm, GrimAge, in a smaller sample of participants who had DNA methylation data (n = 1,418).
Results:
FSRP and HOMA-IR explained 8.3–20.5% of associations of higher moderate-to-vigorous physical activity (MVPA), higher steps, and lower sedentary time with higher brain volume. Additionally, FSRP and GrimAge explained 10.3–22.0% of associations of physical inactivity with lower white matter diffusivity and FSRP explained 19.7% of the association of MVPA with lower free water accumulation.
Conclusion:
Our results suggest that cardiometabolic risk factors and epigenetic patterns partially mediate the associations of physical inactivity with lower brain volume, higher white matter diffusivity, and aggregation of free water in the extracellular compartments of the brain.
INTRODUCTION
Individuals who participate in more physical activity have a lower risk of developing dementia and Alzheimer’s disease [1]. Physical activity has many beneficial effects that may lead to maintenance of brain structure and function into older age. Previous epidemiological studies have reported consistent associations of physical activity with total brain volume and white matter (WM) microstructural integrity [2–4]. There is even limited evidence from exercise interventions demonstrating improved WM integrity [5].
The brain’s WM microstructure, assessed using brain MRI imaging measures such as peak width of skeletonized mean diffusivity (PSMD), WM hyperintensities volume (WMHV), and free water, has been shown to degenerate in older age, and these changes are related to impairments in cognitive function [6, 7]. Cardiometabolic risk factors have also been associated with WM microstructural changes [8]. The effect of physical activity on lowering cardiometabolic risk factors is, of course, well known [9]. Participation in regular physical activity is one of the key components of the American Heart Association’s Life’s Simple 8 to support heart health [10] and is endorsed by the American Diabetes Association for the prevention and management of type 2 diabetes mellitus (DM) [11]. Recently, we also reported that physical inactivity was associated with higher GrimAge [12], a DNA methylation pattern (or “epigenetic clock”) that tends to increase in older age and predicts mortality [13]. Many of these epigenetic age-prediction algorithms have also been associated with markers of structural brain aging and cognitive aging in other observational studies [13–16]. In summary, there is much supporting evidence to suggest that epigenetic factors and cardiometabolic risk factors, among many other potential factors [17], could mediate an association between physical inactivity and brain aging.
In this investigation, we examined cardiometabolic risk factors in the Framingham Heart Study (FHS), including the homeostatic model of insulin resistance (HOMA-IR, the ratio of fasting glucose to insulin) and the Framingham Stroke Risk Profile (FSRP), a score, recently updated in 2017 [18], using weights for age, prevalent cardiovascular disease (CVD), atrial fibrillation (AF), DM, smoking status, antihypertension medications and systolic blood pressure. We further explored the epigenetic age-prediction algorithm, GrimAge. We hypothesized that we would observe mediation by cardiometabolic and epigenetic factors in the association of physical inactivity with lower brain volume and with higher age-related markers of WM microstructure.
METHODS
The Original FHS cohort were recruited from residents of the town of Framingham, Massachusetts in 1948 as an epidemiological study of CVD. The FHS Offspring (Second Generation) cohort began enrollment in 1971 from the children (and their spouses) of the Original FHS, longitudinal cohort study [19]. The Third Generation cohort began enrollment in 2002, recruiting adult children of the Offspring cohort. The New Offspring Spouses cohort also began in 2002, enrolling spouses of the Offspring cohort who were not already enrolled in FHS [20]. Finally, the Omni 2 cohort began enrollment in 2003, expanding the FHS cohort to represent an ethnically diverse group at least 10% of the size of the Third Generation cohort. The Original cohort was re-examined every 2 years, while the Offspring, Third Generation, and Omni FHS cohorts were typically re-examined every 4–6 years. Each study examination included a detailed medical history, a cardiovascular-focused physical examination, and assessment of cardiovascular risk factors, among many ancillary studies that have explored other phenotypes and organ-systems. Participants were included in the current investigation if they provided a brain MRI at a call-back to Offspring exam 9 (n = 1,085, mean time between exam 9 and brain MRI was 1.7±1.0 years), Third Generation exam 2 (n = 2,013), New Offspring Spouse cohort at exam 2 (n = 19), or Omni 2 cohort at exam 2 (n = 120), but excluded if they were diagnosed with dementia, had history of stroke, or other factors that could affect the brain MRI results (n = 159). Participants were also excluded if they did not provide valid accelerometry data (n = 549) or if they were missing any of the cardiometabolic risk factors for the mediation analysis (n = 22 were missing risk factors for FSRP and HOMA-IR mediation [n = 18 missing AF, n = 3 missing DM, n = 1 missing fasting glucose]). Our final sample included 2,507 participants for both the FSRP mediation analysis and the HOMA-IR mediation analysis (Fig. 1). We also explored mediation by epigenetic markers of aging in a subset of our participants with DNA methylation data (n = 1,418). All participants provided written informed consent. The institutional review board at Boston University Medical Center approved the study protocols.
Flow diagram of our study sample. FSRP, Framingham Stroke Risk Profile; HOMA-IR, Homeostatic Model of Insulin Resistance.
Physical activity assessment
All participants were asked to wear an omnidirectional accelerometer (Actical model no. 198-0200-00; Philips Respironics) on the hip for 8 days for 24 h/day for Third Generation/New Offspring Spouses/Omni 2 and only for all waking hours for Offspring, except when bathing. The wearing instructions were altered for the Offspring cohort due to difficulty distinguishing between waking and sleeping hours in Third Generation data. Wear time differences between cohorts were addressed in data processing, described below. The Actical accelerometer records signals within 0.5–3 Hz and accelerations/decelerations within 0.05–2 g. Recorded signals are grouped into “counts” or “steps” at 30-s intervals and averaged over 1-min intervals. Data was analyzed at the FHS using customized software (Kinesoft, version 3.3.63) and a predefined protocol for quality control [21]. Data were processed using a SAS program developed by Colley et al. [22] Non-wear time was removed from data processing using the Choi algorithm [23]. Next, we removed a sliding window of 6 h in each 24-h period (starting at 10 pm on the first day and each subsequent day), during which the lowest total number of counts were accumulated. These windows were removed to account for potential sleep time and could include non-wear time already “removed” by the Choi algorithm. An adherent (valid) day of Actical wear was defined as at least 10 h of wear time (out of a maximum of 18 h) on at least 3 days.
Each minute of wear time was classified using the following intensity cutpoints: moderate to vigorous physical activity (MVPA) was defined at a threshold of ≥1,535 counts/min [22]; sedentary time was defined as any minute with≤100 counts/min [24]. Steps accumulated each day were averaged over all valid days. In order to normalize our skewed steps/day data, we Winsorized at 20,000 steps/day to address 33 outliers. Because sedentary time can vary depending on the amount of time the accelerometer was worn each day (ranging from 10–18 h), we standardized sedentary time to wear time each day by dividing sedentary time/wear time and multiplying by 18 h. The number of steps taken or the time spent in MVPA or sedentary time were accumulated each day and then averaged over all valid days to give average steps/day and time/day spent in sedentary or MVPA, which were used as predictor variables in analysis.
Structural brain MRI
Participants’ brain structures were evaluated with a 1.5-T Siemens Avanto scanner. Three sequences were used: 3D T1-weighted coronal spoiled gradient recalled echo acquisition, fluid-attenuated inversion recovery (FLAIR) sequence, and diffusion tensor imaging (DTI). Methods for segmentation and quantification of total cerebral brain volume (TCBV) and WMHV have been described previously [25]. Briefly, TCBV and WMHV were corrected for differences in head size by computing these variables as ratios with respect to total cranial volume. The interrater reliabilities ranged between 0.90 and 0.94 for total cranial volume, TCBV, and WMHV. All measurements were performed centrally by raters blind to clinical characteristics (QUANTA 6.2, Sun Microsystems Ultra 5 workstation, Santa Clara, CA). WM microstructure was assessed using PSMD and free water calculated from DTI [8, 26]. Briefly, PSMD is a measure of the diffusivity of fluid along white matter tracts in the brain, that is designed to eliminate the contaminating signal of cerebrospinal fluid [26]. When WM tracts are injured, fluid can more easily diffuse out of axons through damaged myelin sheaths, increasing PSMD. Free water refers to the fraction of extracellular water content in the brain interstitial space [8].
Cardiometabolic risk factors and covariates
Participants underwent measurement of blood glucose and insulin after an overnight fast. Insulin was measured using an immunoassay (Roche Diagnostics, Mannheim, Germany), with an intra-assay coefficient of variation (CV) of 2.0% and interassay CV of 4.5% in the Third Generation cohort and an intra-assay CV of 1.3% and interassay CV of 2.1% in the Offspring cohort. HOMA-IR, which was calculated as glucose (mg dL–1)×insulin (μU mL–1)/405 [27]. We also calculated the 2017 revised version of the FSRP [18], using weights for age, prevalent CVD, AF, DM, smoking status, antihypertension medications, and systolic blood pressure. CVD was defined as prevalent coronary artery disease, congestive heart failure, or peripheral vascular disease; AF was defined as arrhythmia noted by a FHS cardiologist on any ECG before or at the time of the FHS examination. We defined DM as a fasting plasma glucose level ≥7 mmol/L, a casual plasma glucose level≥11.1 mmol/L, or the use of insulin or an oral hypoglycemic drug.
DNA methylation
To assess DNA methylation, first, peripheral blood samples were collected and DNA was extracted from the buffy coat. DNA samples were bisulphite converted and hybridized to Human-Methylation450 BeadChips (Illumina Inc, San Diego, CA). Quantification was performed by the Biconductor minfi package with background correction at each laboratory to generate methylation beta values, as described in the Supplementary Methods. The GrimAge methylation equation incorporates age, sex, methylation markers of a series of biomarkers associated with morbidity and mortality, and methylation markers of smoking pack-years [28]. GrimAge was calculated based on 1030 CpG probes, using the sum of the beta values multiplied by the respective regression coefficients reported previously [28–31]. We chose this epigenetic clock because we previously reported associations of physical activity and sedentary time with GrimAge in our FHS study sample [12]. The calculation for GrimAge methylation age was performed online at https://dnamage.genetics.ucla.edu/home [30, 32]. Participants’ methylation age (GrimAge) was then regressed on their chronologic age at the time of blood collection to create age acceleration residuals, “GrimAge-predicted age residuals.” These age prediction models were fit separately for each of three cohort-laboratory combinations (because samples were processed by two different laboratories and one laboratory processed some samples from the Offspring cohort and some from the Third Generation cohort, as described in detail in the Supplementary Methods), adjusting each model for laboratory-specific technical variables.
Statistical analysis
We first performed multivariable linear regression analysis to examine the total effect (c + [a*b]) for the association of MVPA, steps, and sedentary time with brain MRI markers, adjusting for age, age-squared, sex, accelerometer wear time (only for steps and MVPA), cohort, time from exam to MRI, and season of exam. Next, we tested these associations for mediation by FSRP, HOMA-IR, and GrimAge residuals. The indirect (mediation) effect was calculated as (a*b), which is the product of the association of the predictor variable with the mediator (a) and the mediator with the outcome (b). Because the sampling distribution of the indirect effect product term is usually not normal, we used a bootstrapping method that does not make strong assumptions about the distribution of the coefficient of interest to calculate the standard error of the indirect effect. 5000 bootstrap draws were performed for each model. These associations were determined as statistically significant at the p < 0.05 level. In all analyses, MVPA, FSRP, HOMA-IR, WMHV, free water, and PSMD were log transformed to normalize their distributions and all variables except for sex and season were analyzed as continuous variables.
For analyses that had a statistically significant total effect (c + [a*b]) and indirect effect (a*b), we calculated the percent of the association of physical activity or sedentary time with structural brain MRI measures explained by mediation factors (FSRP, HOMA-IR, and GrimAge residuals). Percentages were calculated by dividing the indirect effect by the total effect. Interaction by sex was tested in the primary analysis model (the total association of physical activity or sedentary time with TCBV) by adding a product term in the models and was considered statistically significant if p < 0.1. We also stratified our FSRP results by sex for these primary analyses because FSRP uses different weights by sex. All analyses were performed using the lavaan package in R.
RESULTS
Participants had a mean age of 53.9 years, with a relatively low prevalence of DM and CVD (6.5 and 7.2%), with 26.8% taking anti-hypertensive medications (Table 1). More than 86% of participants attended at least some college and only 0.5% did not graduate from high school. On average, participants walked 7,413 steps/day and spent 19.3 min/day in MVPA and 13.8 h sedentary during a standard 18 h of awake time per day.
Characteristics of the study sample
The associations of higher MVPA, higher steps, and lower sedentary time with higher brain volume (TCBV) were partially mediated through FSRP and HOMA-IR, but not through GrimAge (Fig. 2). FSRP accounted for 8.3–20.5% and HOMA-IR accounted for 13.9–20.5% (p < 0.05, Table 2) of the total association of physical inactivity with brain volume. FSRP, but not HOMA-IR, also partially explained associations of accelerometry variables with lower WM diffusivity (PSMD), accounting for roughly 10.3–22.0% of the total associations, p < 0.05. Differences in FSRP also accounted for 19.7% of the association of higher MVPA with higher free water. Furthermore, GrimAge explained 11.9% of the association of higher step count with lower PSMD.

Associations of physical activity and sedentary time with TCBV, partially mediated by FSRP and HOMA-IR (n = 2,507) and GrimAge (epigenetic age predicted) residuals (n = 1,418). Total = c+(a*b). *p < 0.05 and **p < 0.01, respectively. Grey shading represents unfavorable factors. The arrows indicate the direction of the association with a favorable activity profile (more physical activity or less sedentary time). MVPA, Moderate-to-vigorous physical activity; FSRP, Framingham stroke risk profile; HOMA-IR, homeostatic model of insulin resistance; TCBV, total cerebral brain volume. The following variables were log-transformed: MVPA, FSRP, HOMA-IR, WMHV, free water, and PSMD. Adjustment model: age, age-squared, sex, accelerometer wear time (for steps and MVPA), cohort, time from exam to MRI, and season of exam. For models assessing mediation by GrimAge residuals, we additionally adjusted for the laboratory indicator.
Percent of the association of physical activity
MVPA, Moderate-to-vigorous physical activity; FSRP, Framingham stroke risk profile; HOMA-IR, homeostatic model of insulin resistance; TCBV, total cerebral brain volume; PSMD, peak width of skeletonized mean diffusivity; WMHV, white matter hyperintensities volume. Adjustment model: age, age-squared, sex, accelerometer wear time (for steps and MVPA), cohort, time from exam to MRI, and season of exam. For models assessing mediation by GrimAge residuals, we additionally adjusted for the laboratory indicator. Percentages calculated by dividing the indirect effect by the total effect, but only shown when the indirect effect and total effect were statistically significant (p < 0.05) and in the same direction. – Indirect effect was not statistically significant. †Indirect effects were statistically significant, but total associations were not statistically significant. FSRP explains 46.7% of the association of MVPA with WMHV; FSRP explains 32.1% and 31.6%, respectively, of the associations of steps and sedentary time with free water; GrimAge residuals explain 27.3% of the association of steps with free water. But because the total associations were not statistically significant, we did not include those mediation percentages in this table (see Fig. 2). ‡Indirect effects were statistically significant, but total associations were in the opposite direction, so percentage could not be calculated (see Fig. 2).
Of note, we also observed an unexpected association of lower sedentary time with higher WMHV (beta = –0.036, p = 0.008, Fig. 3), but the mediation (indirect) effect of FSRP was in the opposite direction (beta = 0.005, p = 0.003), and therefore, not a contributor to this unexpected association. We had a similar observation of a negative indirect effect of GrimAge (p < 0.05), acting on an unexpected positive association of higher step count with higher WMHV (p = 0.127).
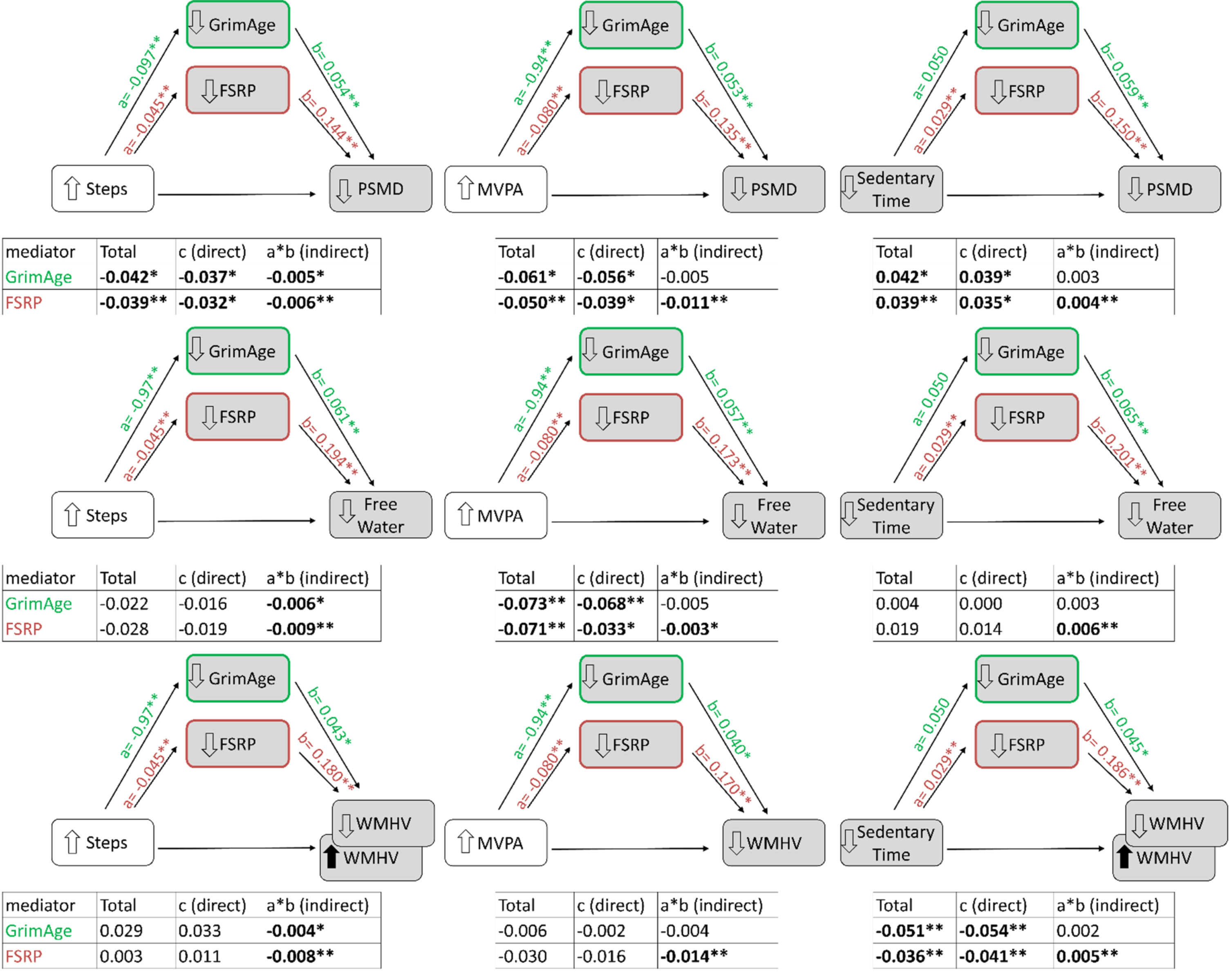
Associations of physical activity and sedentary time with white matter microstructure, partially mediated by FSRP (n = 2,507) and GrimAge (epigenetic age predicted) residuals (n = 1,418). Total = c+(a*b). *p < 0.05 and **p < 0.01, respectively. Grey shading represents unfavorable factors. The up/down arrows in front of each factor indicate the direction of the association with a favorable activity profile (more physical activity or less sedentary time). The association of steps and sedentary time with WMHV were in the unexpected direction (shown with a black arrows) and the indirect “mediation” by FSRP is in the opposite direction of the total association (shown with opposing signs in front of the indirect versus total association beta estimate and by inclusion of a second WMHV box with the opposite arrow direction). Mediation models not shown for HOMA-IR because HOMA-IR was not associated with PSMD, WMHV, or free water (see Supplementary Table 1). MVPA, Moderate-to-vigorous physical activity; FSRP, Framingham stroke risk profile; TCBV, total cerebral brain volume; PSMD, peak width of skeletonized mean diffusivity; WMHV, white matter hyperintensities volume. The following variables were log-transformed: MVPA, FSRP, WMHV, free water, and PSMD. Adjustment model: age, age-squared, sex, accelerometer wear time (for steps and MVPA), cohort, time from exam to MRI, and season of exam. For models assessing mediation by GrimAge residuals, we additionally adjusted for the laboratory indicator.
Stratification by sex
Because our FSRP score is calculated using different weights for sex, we stratified our primary analyses by sex (Supplementary Table 2). We observed statistically significant interaction by sex for associations of accelerometry variables with TCBV, when FSRP was included as a mediator in the model, which was set a priori at p < 0.1 (but not when HOMA-IR or GrimAge residuals were included as mediators, p > 0.1). We observed that the total associations of accelerometry variables (steps, MVPA, and sedentary time) with TCBV were 2-3 times stronger and only statistically significant for women. The associations of FSRP with TCBV were similarly stronger for women. In contrast, the association of accelerometry variables with FSRP were almost twice as strong for men. Despite these differences, the indirect associations (mediation effect) of FSRP for the association of accelerometry variables with TCBV were very similar. For this reason, we chose to report the associations unstratified in Fig. 2 because that was our primary interest in this investigation.
DISCUSSION
In this mediation study, we observed that both cardiovascular (FSRP) and metabolic (HOMA-IR) risk factors mediated the association of physical inactivity and sedentary time with smaller brain volume (TCBV). But, for WM microstructural integrity, cardiovascular risk factors and differences in epigenetic patterns (rather than metabolic factors) appeared to play a more substantial role in mediating the association of physical inactivity with these brain imaging measures. Our investigation supports the notion that cardiometabolic risk factors and certain epigenetic patterns, associated with physical inactivity, may be involved in mechanisms contributing to brain aging.
Our observation that HOMA-IR mediated the association of physical activity with TCBV is in agreement with a previous (much smaller, n = 134) study by Felisatti et al. [33] that reported mediation by insulin in the association of self-reported physical activity with MRI markers of brain aging. However, in contrast to our findings, Felisatti et al. did not observe associations of physical activity with white matter integrity or WMH, and instead they reported associations of physical activity with gray matter and hippocampal volume. Although other studies have also observed associations of physical activity with gray matter and hippocampal volume [34, 35], we have previously reported lack of this association in FHS [4]. Differences between our studies that may explain these different results include our much larger study size (n = 2,507) differences in age ranges and educational status, and use of an objective measure of physical activity and sedentary time. We built onto this previous study with assessment of mediation by HOMA-IR (a variable incorporating fasting glucose and insulin), and FSRP (which predicts risk of stroke, by incorporating risk factors including CVD, AF, DM, smoking, and hypertension). We observed that FSRP not only mediated associations of physical activity with total brain volume, but also with additional brain imaging outcomes (PSMD and free water). PSMD is a measure of low WM integrity, calculated using diffusion tensor imaging to assess small vessel disease-related injury in the brain through quantification of fluid motion along neuronal axons [36] Dawe et al. conducted a mediation analysis in the Rush Memory and Aging Project, observing that there are specific WM microstructural tracts in the brain that explain about 12–17% of the association of physical activity with cognition (episodic memory and perceptual speed), suggesting that the integrity of the brain WM at these specific regions may be important to explore in future research as playing a role in the relationship of physical activity in prevention of dementia [37]. WM integrity has been previously related to lower arterial stiffness in FHS [8]. In addition to WM integrity, a measure called “free water” was associated with higher arterial stiffness. Free water reflects the extracellular water molecules in the brain, which may be another biomarker of subtle injury [8].
One mechanistic explanation for our findings could be that elevated factors making up HOMA-IR and FSRP (e.g., elevated glucose and blood pressure), and their accompanying inflammatory environment, contribute to microvascular injuries to the brain, leading to lower WM integrity (i.e., higher diffusivity, PSMD) and higher free water in extracellular spaces. The role of physical activity in preventing this brain injury is speculated to involve a favorable inflammatory environment, stimulation of growth and neurotrophic factors (such as brain derived neurotrophic factor [BDNF]), and may also involve epigenetic changes [13, 17]. Although bouts of physical activity stimulate BDNF and other factors, BDNF stimulation appears to be only transient [38]. Therefore, chronic circulating BDNF levels may not reflect habitual physical activity levels for individuals. We previously demonstrated a lack of association of accelerometer-measured habitual physical activity with BDNF in FHS [39]. Consequently, we chose not to explore BDNF as a mediator for our investigation, but acknowledge the substantial evidence suggesting its role in the protective effect of physical activity on brain aging [17]. Similar transient effects of physical activity may also make it challenging to assess mediation by inflammatory cytokines and epigenetic patterns. However, we did explore mediation by a newer epigenetic pattern that has been associated with brain aging and mortality, called GrimAge [13, 28,40]. Epigenetic patterns, such as GrimAge, are a reflection of the cumulative biological changes representing the aging process, including inflammatory and oxidative stress pathways that may affect neurogenesis and synaptic plasticity [29]. We recently reported associations of lower physical activity and higher sedentary time with GrimAge in the FHS sample [12]. In the current study, GrimAge mediated the association of the number of steps walked per day with WM diffusivity (PSMD). Investigators have previously reported associations of epigenetic age-prediction algorithms, including GrimAge, with WMHV, volumetric measures of the brain, and cognitive function [13–16]. Although we observed similar associations of GrimAge with the brain imaging markers we measured (TCBV, PSMD, free water, and WMHV), GrimAge did not mediate other associations of physical inactivity with brain aging.
One set of nuanced results from our investigation that deserve elaboration are those involving WMHV. WMHV is a MRI indicator of cerebrovascular injury and has been associated with DM, fasting glucose levels, and other cardiometabolic markers in FHS and other studies [41, 42]. In our sample, we did observe a positive association of FSRP and GrimAge (but not HOMA-IR) with WMHV. On the other hand, as we have previously reported in similar analyses [4], we also observed an unexpected association of lower sedentary time with higher WMHV. It is important to note that we also observed statistically significant mediation by FSRP in the association of sedentary time with WMHV in the expected (positive) direction. It was the negative total association of sedentary time with WMHV that was unexpected, and therefore, is likely due to factors other than cardiometabolic risk factors. In agreement with our unexpected findings, we have also previously observed an unexpected association of higher sedentary time with favorable cognitive outcomes [43]. We speculate that there may be certain sedentary tasks that are cognitively complex and protective of the brain, such as reading and computer use [44–46]. In addition to possible confounding by cognitively complex sedentary tasks, it is also possible that being more physically active leads to other changes in behaviors that affect brain health, such as stress, mood, and sleep, or confounding by unmeasured sociodemographic factors [34].
A major strength of our investigation is the large sample size, with objective measures of physical activity, sedentary time, structural brain aging, as well as cardiometabolic factors. However, we limited the number of brain MRI measures we explored to focus on TCBV and WM integrity. We also limited our stratified analyses to focus on sex, but acknowledge that other factors, such as genetic variation may also modify the associations we observed. The cross-sectional nature of our study design limits our ability to determine causality and increases the likelihood that the associations we observed are bidirectional. Another limitation of our investigation is that participants in the FHS have a largely homogeneous racial and ethnic profile, as well as a lack of diversity in other demographic aspects, having mostly originated from families living in Framingham, Massachusetts in 1948. Therefore, we hope that the approaches used in our proposal are replicated in future studies with more diverse populations.
Conclusions
Physical activity has many beneficial effects on cardiometabolic and brain health. Our results suggest that the associations of physical inactivity with lower brain volume, poorer WM microstructure, and aggregation of free water in the extracellular compartments of the brain are partially mediated by cardiometabolic risk factors and also by certain epigenetic patterns (GrimAge). Achieving a better understanding of the mechanisms by which physical activity may prevent brain aging can help us to realize the modifiable nature of our risk for dementia. It may also help us design interventions that target the most vulnerable risk groups. Our results warrant future research on the impact of physical inactivity on cardiometabolic risk factors, which may have implications for accelerating brain aging.
Footnotes
ACKNOWLEDGMENTS
The authors acknowledge the commitment of the Framingham Heart Study participants as well as research support staff, who are essential to our team.
FUNDING
Framingham Heart Study contract: NHLBI-N01-HC25195, HHSN268201500001I, 75N92019D00031; with additional support from the following grants from the NIH (R01-AG047645; R01-HL131029; R01-HL136266; R01-AG054076; R01-AG049607; R01-AG066524; RF1-AG059421; U01-AG052409; P30AG066546; R01-AG059421; R01-NS017950); American Heart Association (15GPSGC24800006 and 16MCPRP30310001); Alzheimer’s Association (2018-AARG-591645).
CONFLICT OF INTEREST
Dr. Spartano received funding from Novo Nordisk for an investigator-initiated research grant unrelated to the current project. Dr. Murabito was a guest lecturer for Merck unrelated to this work.
DATA AVAILABILITY
The Framingham Heart Study makes phenotypic and genetic data available through the online repositories BioLINCC and dbGap.