
Abstract
Probabilistic and parsimony-based arguments regarding available genetics data are used to propose that Hardy and Higgin’s amyloid cascade hypothesis is valid but is commonly interpreted too narrowly to support, incorrectly, the primacy of the amyloid-β peptide (Aβ) in driving Alzheimer’s disease pathogenesis. Instead, increased activity of the βCTF (C99) fragment of AβPP is the critical pathogenic determinant altered by mutations in the APP gene. This model is consistent with the regulation of APP mRNA translation via its 5’ iron responsive element. Similar arguments support that the pathological effects of familial Alzheimer’s disease mutations in the genes PSEN1 and PSEN2 are not exerted directly via changes in AβPP cleavage to produce different ratios of Aβ length. Rather, these mutations likely act through effects on presenilin holoprotein conformation and function, and possibly the formation and stability of multimers of presenilin holoprotein and/or of the γ-secretase complex. All fAD mutations in APP, PSEN1, and PSEN2 likely find unity of pathological mechanism in their actions on endolysosomal acidification and mitochondrial function, with detrimental effects on iron homeostasis and promotion of “pseudo-hypoxia” being of central importance. Aβ production is enhanced and distorted by oxidative stress and accumulates due to decreased lysosomal function. It may act as a disease-associated molecular pattern enhancing oxidative stress-driven neuroinflammation during the cognitive phase of the disease.
Keywords
30 YEARS OF THE AMYLOID CASCADE HYPOTHESIS
In 1992 Hardy and Higgins published their famous paper, “Alzheimer’s disease: the amyloid cascade hypothesis” [1]. The paper captured the excitement of the Alzheimer’s disease (AD) research field over the emerging genetic evidence implicating the Amyloid Beta A4 Precursor Protein (APP) gene in the inherited, early onset form of the disease (familial AD, fAD). In the following decades, this paper has been regarded as the keystone supporting the idea that AD pathogenesis is driven by accumulation of the amyloid-β (Aβ) peptide, which is derived from AβPP after this protein is cleaved by β- and γ-secretases (Fig. 1). Any theory of AD pathogenesis seeking to challenge the central role of Aβ in AD pathogenesis must provide a more parsimonious explanation of the genetic data.
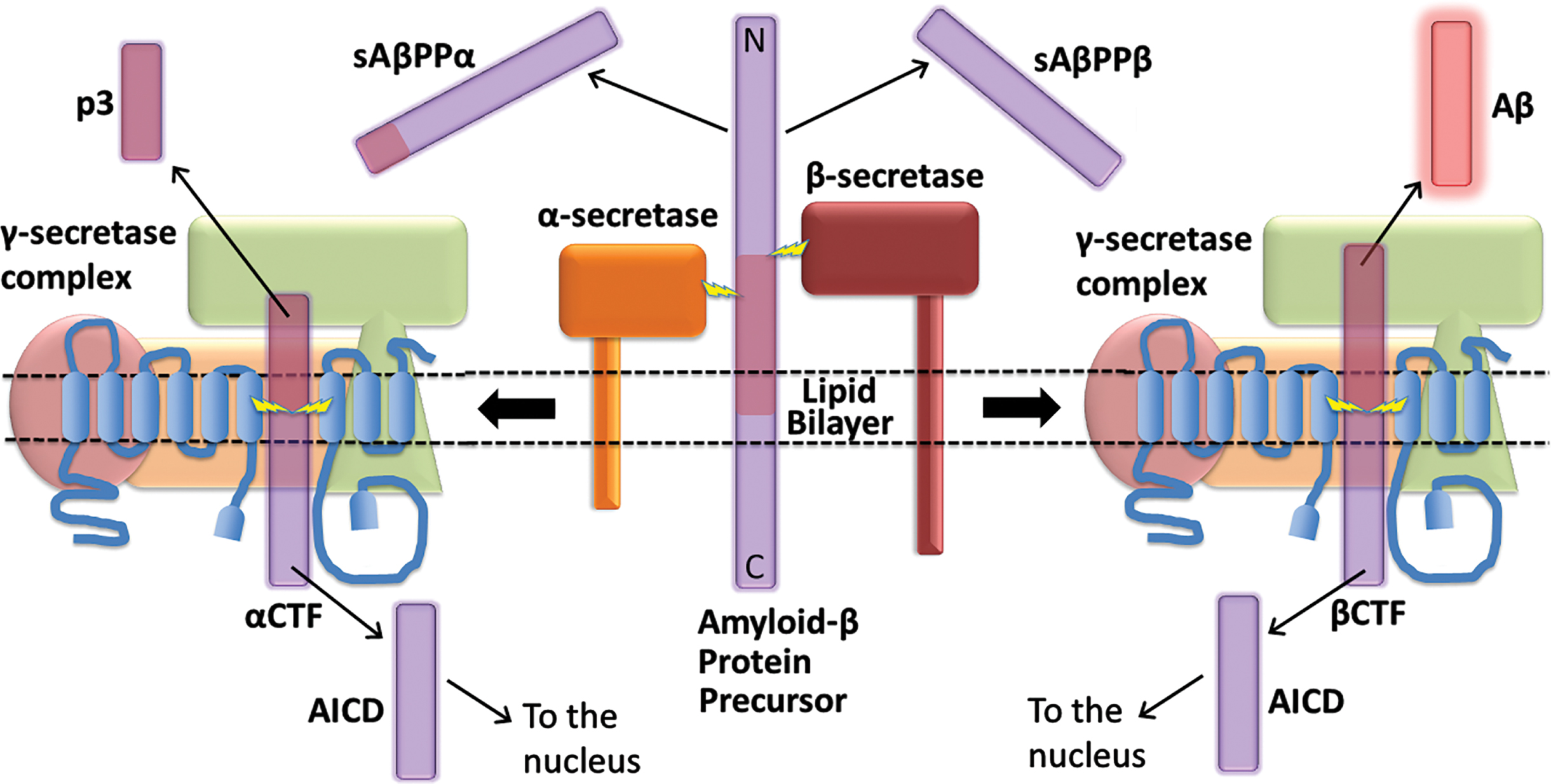
In this paper, we will use arguments based in probability and parsimony to assert that the “Amyloid Cascade Hypothesis” as originally presented by Hardy and Higgins is correct but has subsequently been interpreted too narrowly. Similarly, we will also examine the fAD-promoting mutations in the genes Presenilin 1 (PSEN1) and Presenilin 2 (PSEN2) to argue that their immediate pathogenic effects do not occur via accumulation of Aβ.
MUTATIONS ARE MORE LIKELY TO DESTROY FUNCTION THAN IMPROVE IT
The amazing complexity of cellular structures and of living multicellular organisms today only exists because past changes in genetic information (mutations) have, fortuitously, resulted in improved reproductive competitiveness. However, such beneficial mutations are relatively uncommon (improbable). The great majority of mutations result either in no substantial change in cellular function or are detrimental and inhibit reproductive success. The “screwdriver analogy” illustrates this. If a photocopier (a machine for reproducing images on paper) has its screws tightened or loosened at random, most of these changes will likely have little effect, some changes will degrade photocopy quality, while a very few improbable adjustments may actually improve it.
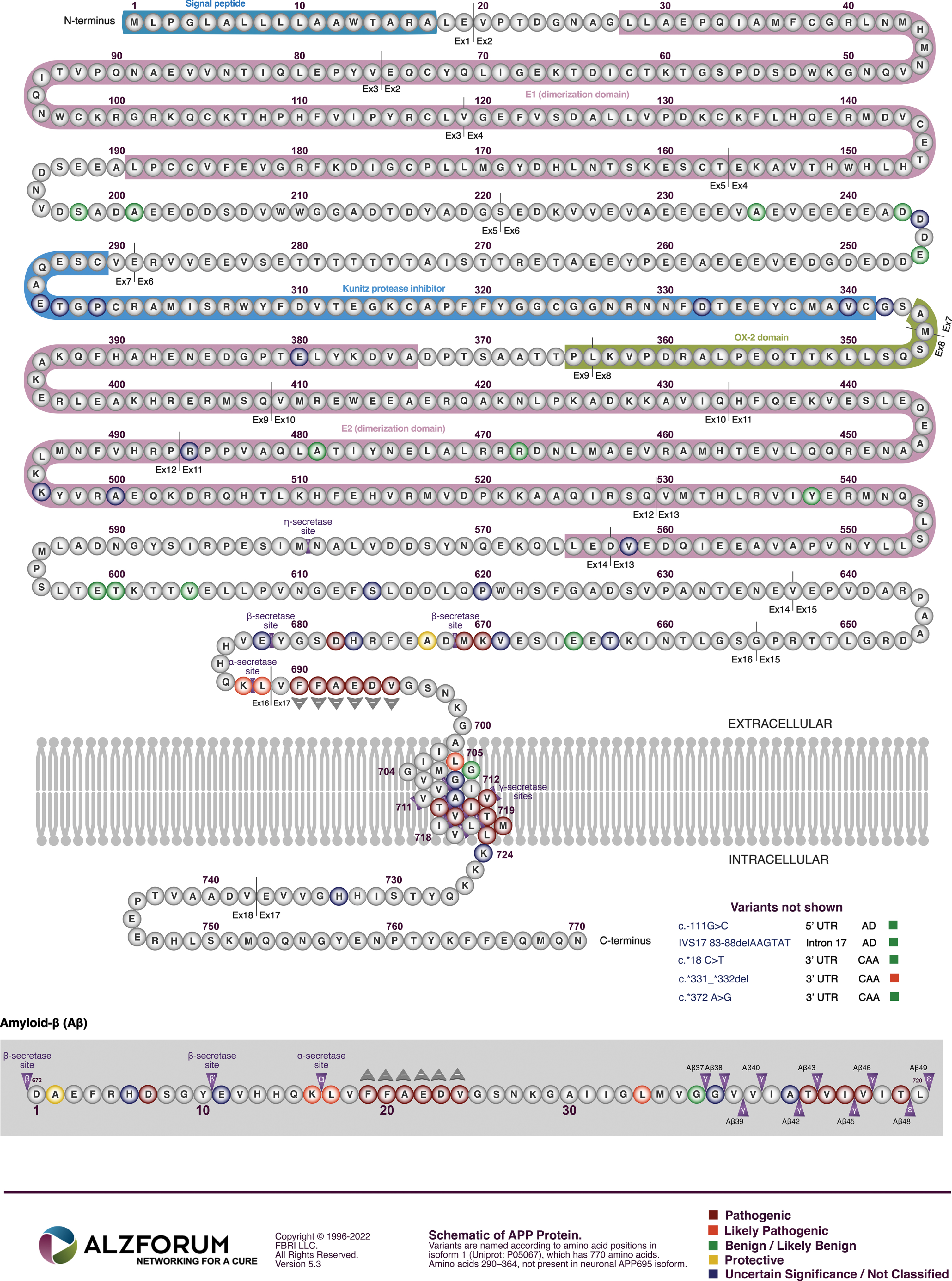
The phenomenon that mutations with no effect are far more likely than mutations with negative effects, that themselves are far more likely than mutations with positive effects, can be referred to as the “mutation effect probability hierarchy” (MEPH). It is well illustrated by examination of mutations in the gene APP (Fig. 2). The vast majority of genetic variations (mutations) in APP in the human population have no known relevance to AD—this is easily seen by inspecting the data collated in the gnomAD database [2]. When Hardy and Higgins published their hypothesis paper in 1992, it was based on identification of three AD-causative mutations in one codon (V717) of the APP open reading frame and one mutation (E693Q) causing a related disease now known as cerebral amyloid angiopathy, APP-related, Dutch variant. Today, we know of at least 27 such deleterious mutations in APP (see Fig. 2 and the mutations collated at Alzforum.org). However, only one, uncommon mutation (A673T) found mainly in the small population of Iceland is known to promote mental health by reducing AD risk [3–8].
THE CURRENT “UNDERSTANDING” OF fAD GENETICS
Of the ∼20,000 protein-coding genes in humans, only three are known definitively to host mutations causing AD: APP, PSEN1, and PSEN2 [9, 10]. A fourth gene, Sortilin-Related Receptor 1 (SORL1) may also cause fAD before the age of 65 years but its relationship to the other fAD genes is relatively poorly understood [10–13].
APP
Decisive for our understanding of how mutation of APP drives AD is the fact that simple increased expression of the APP coding sequence is sufficient to cause disease. Three doses of APP, whether through trisomy of chromosome 21 [14], or duplication of APP within chromosome 21 [15], causes early onset of AD. This means that increased expression of the AβPP protein or of one of its fragments is highly likely to be driving the pathogenic changes of AD.
Within the APP gene, mutations causing AD are clustered within, and flanking, the sequence coding for Aβ (Fig. 2). This fact is seen as support for the prime role of Aβ in driving AD pathology. According to the MEPH, we would expect most of these mutations to degrade/inhibit particular AβPP functions/abilities. Consistent with the MEPH, we find evidence that many of the mutations in the region coding for the γ-secretase cleavage site decrease the ability of the γ-secretase enzyme to cleave AβPP [16]. Other mutations occur in codons for the α- and β’-cleavage sites of AβPP respectively and decrease the proclivity of these sites for cleavage by their relevant enzymes [17, 18], thereby increasing the likelihood of cleavage at the β-site. Interestingly, one unique mutation, the “Swedish mutation” of APP (KM670/671NL) drives development of AD by improving the proclivity of the β-site for cleavage by the β-secretase enzyme BACE1 [19, 20] (while the aforementioned A673T mutation near this site uniquely improves survival by reducing the ability of β-secretase to cleave at the β-site [3]). The experimental data regarding whether fAD mutations in AβPP increase or decrease Aβ production is remarkably variable. The comprehensive analysis published by Xu et al. [16], that saw nearly all fAD mutations of AβPP decrease γ-secretase cleavage, monitored the γ-secretase cleavage needed to produce Aβ using cells transfected to express a modified βCTF. (This is also known as “C99” and is the transmembrane fragment of AβPP produced when AβPP is cleaved by β-secretase but before cleavage by γ-secretase.) The APP intracellular domain (AICD) of βCTF was fused to a transcription factor to allow detection of cleavage at the ɛ-sites (Fig. 3) through activation of luciferase expression. By reporting on γ-secretase at the ɛ-sites rather than after the subsequent, variable “product line” γ-secretase cleavages (see Fig. 3), the rates of γ-secretase cleavage should be consistently comparable between different fAD mutant forms of βCTF. When, instead, levels of the Aβ product are used to compare γ-secretase cleavage of βCTF mutants in transfected cells, the differing levels of βCTF could have differential effects on lysosomal acidification [21], thereby affecting Aβ degradation and accumulation. This might partially explain, for example, why Xu et al. observed decreased γ-secretase cleavage of AβPP due to the fAD mutation D678 H whereas Chen et al. [22] saw a relative increase in Aβ levels. However, Chen et al. also observed that the D678 H mutation shifted the initial cleavage of AβPP from the α-site to the β-site which would increase βCTF levels and also promote Aβ production. Numerous mutations in the Aβ region of AβPP also affect its tendency to aggregate and form fibrils. However, in many cases these mutations are associated with accumulation of Aβ in brain vasculature in the disease cerebral amyloid angiopathy that is distinct from AD, e.g., APP mutations E693K [23, 24] and L705 V [25].
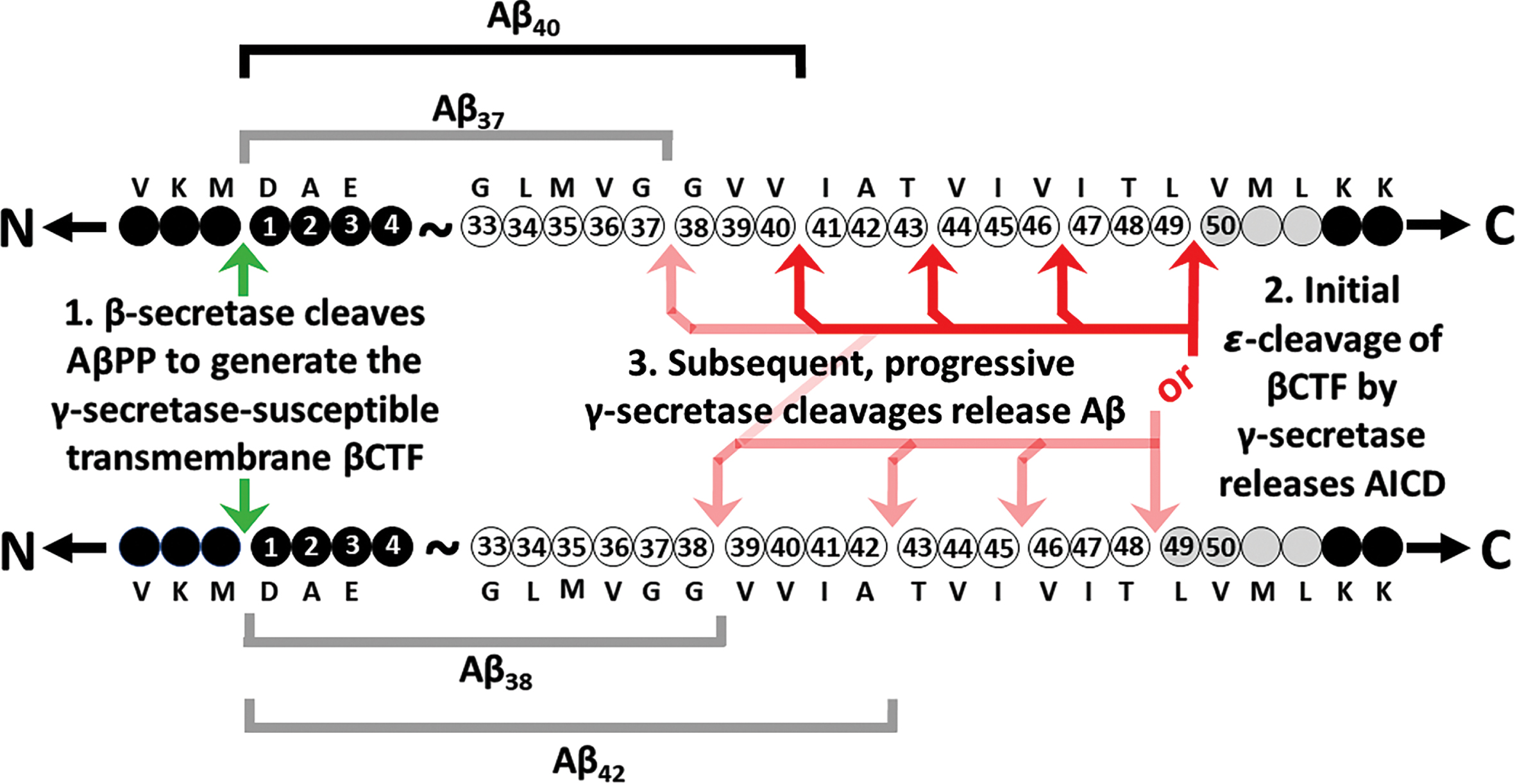
More than 100 transmembrane proteins are subject to cleavage by γ-secretase [26, 27]. However, there is no clear consensus sequence determining proclivity for cleavage by γ-secretase [28] and the mechanism of operation of this enzyme causes variability in the cleavage products [29–31] (Fig. 3). For this reason, Aβ is not one peptide but a family of related sequences of 37-49 amino acid residues (aa) in length depending on the positions of the final γ-secretase cleavages that create their carboxy termini (Fig. 3). The inhibition of γ-secretase cleavage by mutations in/near the region coding for the γ-cleavage site of AβPP [16, 32] is not obviously consistent with a primary role for Aβ in AD as some of these mutations drastically inhibit the production of Aβ [16, 33]. However, rather than excluding an AD-causative role for Aβ, an Aβ-centric hypothesis has evolved positing that the ratio of different Aβ fragment lengths, and the greater tendency to aggregate of longer Aβ fragments, are critical to development of AD, e.g., [34–37]. This idea is inspired by the observation of differences in the ratios of the aggregation-prone 42 aa fragment (Aβ42) relative to the most common, 40 aa Aβ fragment (Aβ40) dependent on brain disease state [35, 38]. Changes in Aβ42/Aβ40 ratios have also been observed in in vitro analyses of fAD mutations of PSEN1 [35–37] and PSEN2 [39–41] (discussed later herein). However, other general explanations for increased Aβ42/Aβ40 ratios in AD brains are that these are caused by the influence of oxidative stress on γ-secretase [42] (since increased levels of oxidative stress are a consistent phenomenon in AD brains [43, 44]) and/or that changes in the composition of the membranes in which AβPP is embedded also affect Aβ42/Aβ40 ratios [45, 46] because AβPP’s β-C-terminal fragment, βCTF, regulates the distribution of cholesterol between the plasma and endoplasmic reticulum (ER) membranes [47] while γ-secretase cleaves βCTF preferentially in cholesterol-rich lipid rafts [48–50].
A difficulty with much of the research analyzing fAD mutations of APP, including their cleavage by γ-secretase, is its conduct using transfected cells that significantly over-express APP. Also, the fAD genes are very responsive to stress (e.g., [51–53]) and cells in tissue culture (such as the transfected cells used in much fAD gene analysis) are frequently stressed by their unusual environmental conditions [54]. In contrast, fAD mutation-carrying humans are almost always heterozygous, raising the possibility of interactions between mutant and non-mutant forms of AβPP or its fragments. Indeed, dimerization of AβPP is known to affect its γ-secretase cleavage to form Aβ [55–57]. Studies using editing of endogenous APP genes in iPSCs to compare heterozygous mutant and non-mutant cells (e.g., [58] below) should produce information that is interpretable with greater certainty (fewer possible variables involved).
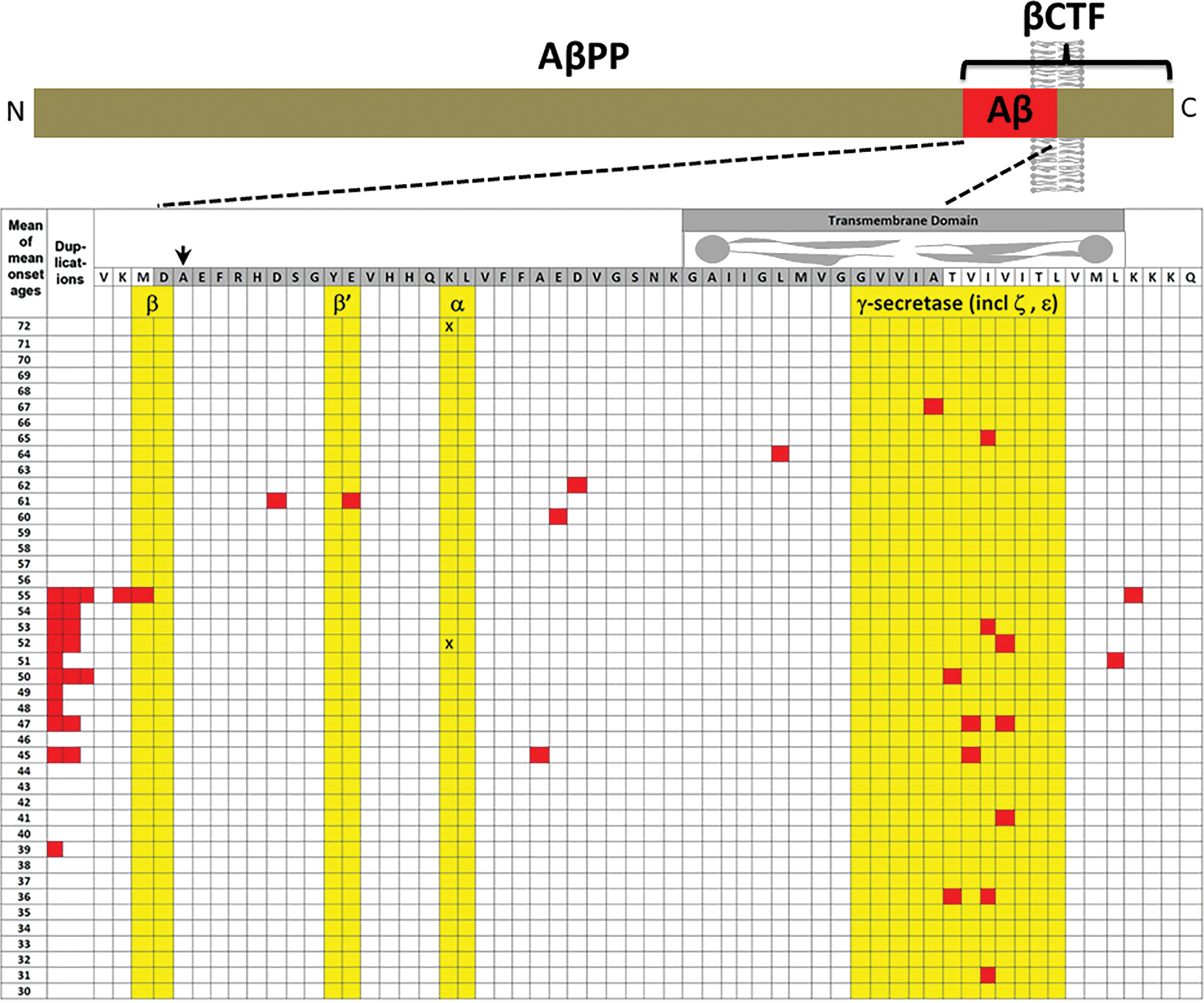
In accordance with the “Law of Parsimony” [59], it is highly likely that all the various mutations in APP cause AD through a single mechanism in common. Based on the information above, this is likely to involve an increase in production of the fragment of AβPP produced by cleavage at the β-site, i.e., βCTF. Increased expression of the APP gene [14, 21], or increased proclivity for cleavage of AβPP by β-secretase [19, 20], or decreased cleavage at AβPP’s α- or β’-cleavage sites that shifts cleavage towards the β-cleavage site [17, 18], or decreased cleavage by γ-secretase [16], should all increase the levels of βCTF. Consistent with this, the most pathogenic mutations in APP causing the earliest onset of AD tend to be those affecting the site of γ-secretase cleavage (Fig. 4) since inhibition of this cleavage would likely be very effective in increasing βCTF levels (as this inhibits the destructive cleavage of βCTF that forms the Aβ and AICD fragments).
THE EXPLANATORY POWER OF INCREASED βCTF LEVELS AS THE AD-CAUSATIVE EFFECT OF MUTATIONS IN APP
Three decades of research focused on Aβ has not yet established convincingly the toxic nature of this peptide under normal, physiological conditions, e.g., [60–64]. The apparent recent successes of anti-Aβ antibody therapies [65] supports a pathological role for this peptide in the clinical phase of the disease, possibly as a disease-associated molecular pattern contributing to the inflammation that is characteristic of AD brains [66, 67]. It is also notable that βCTF includes the Aβ sequence, raising the possibility that these antibody therapies may affect cellular function by altering βCTF levels (since antibodies have been seen to act intracellularly [68]). However, the anti-Aβ antibody therapies are apparently unable to prevent onset of the dominantly inherited disease [69] and only slow the progression of the sporadic, late onset disease rather than stopping it [65]. This supports that something else drives the pre-clinical phase of AD. In interpreting the results of much Aβ-based AD research, it is remarkable how seldom the presence and levels of βCTF are assessed. And yet βCTF levels are likely very significantly altered in transgenic mouse models of AD that include APP transgenes (including the models 3xTg, 5xFAD, A7, APP23, APPswe/PSEN1dE9, PDAPP, J20, PDAPP, PS2APP, Tg2576, Tg-ArcSwe, TgCRND8, [70]), while βCTF has been seen to accumulate in AD brains [71–73]. A critical role for βCTF levels in AD pathogenesis is also consistent with the superior correlation to cognitive decline of levels of soluble Aβ (the immediate product of βCTF cleavage by γ-secretase) in contrast to levels of accumulated insoluble Aβ deposits, e.g., in amyloid plaques [74, 75]. Much of the evidence supporting a role for βCTF in both fAD and sporadic AD was reviewed in a paper by Checler and colleagues [76].
A number of papers below give very strong support for increased βCTF levels as the pathological effect of fAD mutations in APP. Note that the first two papers, and our later discussion of the effects of fAD mutations in PSEN1 and PSEN2, link these genes to normal functioning of the cellular endolysosomal system. Many of the late onset AD risk loci discovered by genome-wide association studies are also important for endolysosomal system function [77, 78]. Indeed, the strongest genetic risk factor for sporadic (non-familial) AD, the ɛ4 allele of APOE (APOE4), reduces lysosomal acidification relative to the ɛ3 allele [79] (in common with fAD mutations in APP [21] and the presenilins [80]).
Kwart et al. [58] examined the subcellular effects of a variety of fAD mutations in APP and PSEN1 after these were edited into a single iPSC cell line (for consistency of genetic background) that was then differentiated into neurons. The mutations uniformly caused accumulation of, specifically, AβPP’s βCTF but had inconsistent effects on Aβ levels. Rab5 + early endosome size was correlated with βCTF levels and could also be increased by chemical inhibition of γ-secretase activity (which reduced Aβ to negligible levels).
Jiang et al. [21] examined fibroblasts from trisomy 21 individuals (Down syndrome) who suffer early onset AD. They showed that the increased dosage of the APP gene (present on chromosome 21) led to increased levels of βCTF. This decreased the acidification of lysosomes (and, thereby, the activity of a marker of lysosomal function, the enzyme cathepsin D). These phenomena could be modulated by various treatments altering the levels of βCTF in normal and Down syndrome fibroblasts. For example, chemical inhibition of β-secretase cleavage of AβPP (to decrease βCTF levels) decreased lysosomal pH in trisomy 21 syndrome fibroblasts but, interestingly, did not lower lysosomal pH in euploid (normal diploid) fibroblasts.
Montesinos et al. [47] found that AβPP’s βCTF regulates the trafficking of cholesterol from the plasma membrane to the ER. Increased levels of βCTF inhibits a cell’s own cholesterol synthesis and increase the rate of cholesterol trafficking from the plasma membrane. The increased ER cholesterol increases formation of the mitochondrial associated membranes (MAM) of the ER through which cells regulate mitochondrial energy production via Ca2+ ion release [81]. Increased MAM formation is observed in fibroblasts from both fAD and late onset AD patients compared to those from cognitively healthy controls [82].
In previous papers some of these same researchers showed that AβPP’s βCTF and the PSEN1 and PSEN2 proteins are all present in the MAM [50, 83] where they regulate sphingolipid and ceramide homeostasis [50].
THE ORIGINAL AMYLOID CASCADE HYPOTHESIS INCLUDED βCTF AS A CANDIDATE PATHOGENIC AGENT
The possibility that βCTF is a pathological agent driving AD is actually suggested by Hardy and Higgins in their 1992 paper. (In the following quotation, the authors refer to Aβ as “AβP” and I have bolded the text referring to βCTF.)
“Our cascade hypothesis states that AβP itself,
“ . . . The evidence we have described supports the hypothesis that the AβP molecule initiates the pathological cascade of Alzheimer’s disease.
Interestingly, in a 1996 paper examining transgenic mice expressing only the βCTF portion of AβPP [84], Neve and colleagues (mentioned above) observed “ . . . oddly shaped secondary lysosomes . . . These inclusions strongly resembled those that we had identified earlier in affected regions of AD brains . . . ”. Recently, Nixon and colleagues have shown that dysmorphic lysosomes in transgenic AD model mice accumulate Aβ and are precursors of amyloid plaques [85].
The likelihood that βCTF represents the toxic agent in APP mutation-driven fAD means that experiments looking at Aβ accumulation in human brains or animal models should also examine βCTF levels to avoid the possibility of inappropriately attributing pathological agency to Aβ.
PSEN1 AND PSEN2
The majority of mutations causing fAD occur in the gene PSEN1 both in terms of their relative frequency among fAD mutations in the human population (∼63%, [10]) and relative to the number of fAD mutations known in APP and PSEN2 (∼80% of the known mutations, [86], Fig. 5). Collectively, fAD mutations in PSEN1 also have the earliest median onset age [87]. Relatively far fewer mutations occur in the gene PSEN2 and these also have a later median age of onset than even mutations in APP [87] (Fig. 6). Importantly, both PSEN1 and PSEN2 encode proteins with similar structures and functions, implying that disruption of a function they share in common causes AD. Intriguingly, while the large number and wide distribution of fAD-causative mutations in the coding sequences of PSEN1 and PSEN2 make it extremely likely that these mutations cause loss of a function(s), two characteristics of these mutations are consistent with action via a gain-of-function mechanism: The great majority of fAD mutations in PSEN1 and PSEN2 are missense mutations altering the protein coding sequence or mutations inserting or deleting entire codons (which, for small indel mutations in coding sequences, is less probable than creation of frameshifts). Even those mutations causing changes in transcript splicing (e.g., PSEN1 L113_I114insT [88]) always produce at least one transcript isoform that preserves the open reading frame to code for a “full-length” protein. We addressed this issue in a previous review [9], describing it as the reading-frame preservation rule. Consistent with this (with the exception of one unique and important gain-of-function mutation discussed later [89]), no mutations or genetic variants affecting PSEN1 or PSEN2 transcript regulation (without changing protein-coding sequences) have been discovered that increase the risk of AD [90]. Loss-of-function mutations that would be expected simply to reduce γ-secretase activity, without otherwise distorting it, do not cause fAD. This is dramatically illustrated by the existence of frameshift mutations in PSEN1 that truncate the open reading frame and do not cause fAD while causing a completely unrelated disease of the skin, Acne Inversa, familial 3, (ACNINV3, also known as hidradenitis suppurativa) [91–93]. Unlike fAD, familial Acne Inversa can also be caused by mutations in genes encoding two other components of the γ-secretase enzyme complex, Nicastrin (NCSTN), and Presenilin Enhancer, Gamma-Secretase Subunit (PSENEN, formerly known as PEN2) [91] (see also Fig. 7). All these mutations affecting different components of the γ-secretase enzyme complex almost certainly act through reduction of cellular γ-secretase activity but mutations causing fAD only occur in PSEN1 and PSEN2 and not in genes encoding other γ-secretase complex components. Therefore, fAD cannot be due to a simple loss of γ-secretase activity. The reading frame preservation rule implies that a gain-of-function mechanism is involved.
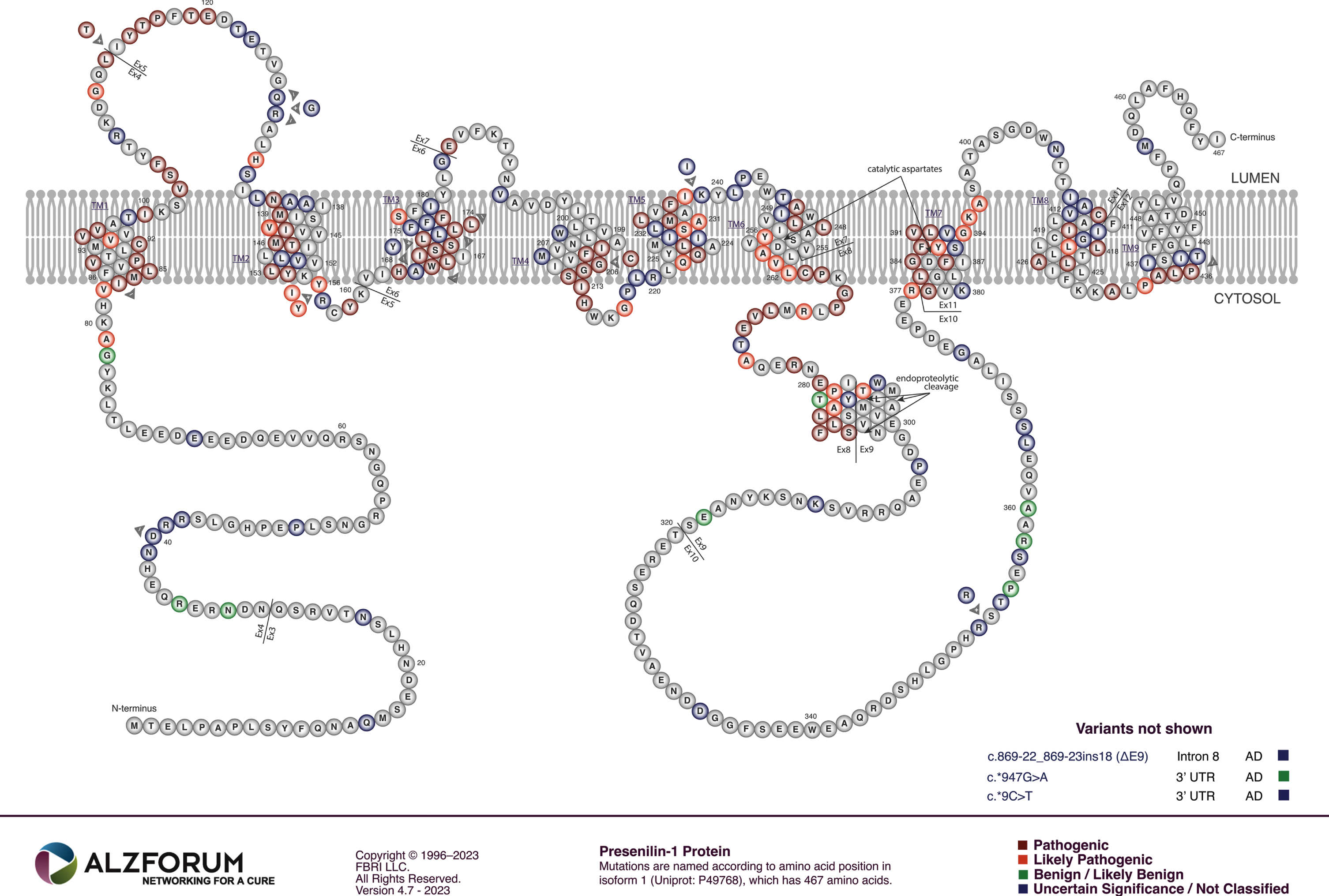

The role of both PSEN1 and PSEN2 in γ-secretase activity, and the fixation of the AD research community on Aβ that requires γ-secretase activity for its production from βCTF, has meant that nearly all research on PSEN1 and PSEN2 has focused on γ-secretase. Since fAD mutations in APP that affect the protein’s γ-secretase cleavage site reduce Aβ production, while fAD mutations in PSEN1 and PSEN2 do not consistently reduce γ-secretase activity (see below), research has focused on one possible commonality of all these mutations—their effects on Aβ peptide length ratios no matter what the total level of Aβ production may be. Some early studies appeared to find correlations between the ages of AD onset caused by different fAD mutations in PSEN1 and the Aβ42/Aβ40 ratio caused by these same mutations in transfected cell culture-based assays [94, 95]. However, the comprehensive analysis of 138 PSEN1 fAD mutations by Sun et al. (2016) [96] using cell-based and in vitro assay systems found these apparent correlations to be statistical artefacts (see Fig. 5 in [96]). That work also found that, while the majority of the 138 mutations they analyzed decreased γ-secretase activity (as measured by combined Aβ40 and Aβ42 production, a loss of function), ∼10% of the fAD mutations increased γ-secretase cleavage of AβPP (a gain of function, at least in their cell-free assay system that over-expressed mutant PSEN1 proteins in the absence of wild type PSEN1). The increased γ-secretase activity of at least one of these mutations, S365A of PSEN1, has been confirmed independently by others [97]. Sun et al. saw that some fAD mutations of PSEN1 lost all apparent γ-secretase activity while others showed decreased Aβ42/Aβ40 ratios rather than increased ratios. In summary, Sun et al. saw no consistent changes in γ-secretase activity (when mutant proteins were assayed in homogeneous states) that would support a role for γ-secretase activity, or for Aβ, as pathogenic agents promoted by fAD mutations in PSEN1.
The idea that Aβ fragment length is the critical determinant of the AD pathological process was reinvigorated by a study published in 2022 (Petit et al. [98]) analyzing a wider variety of Aβ fragments to define a Aβ(37+38+40)/Aβ(42+43) ratio. (They transduced mutant forms of PSEN1 or PSEN2 and a construct expressing AβPP’s βCTF into mouse embryonic fibroblasts lacking endogenous presenilin genes, and then compared the levels of different Aβ fragment lengths in conditioned medium by ELISA.) This ratio reflects the “processivity” of the γ-secretase enzyme (the rate at which it successively cleaves along the transmembrane domain of AβPP after an initial “ɛ-cleavage”) by comparing the common (Aβ40) plus the “maximally complete” Aβ cleavage products (Aβ37 and Aβ38) of the two alternative γ-secretase cleavage pathways of AβPP’s βCTF with the less common “incomplete” cleavage products of these pathways (Aβ42 and Aβ43, Fig. 3). Presumably, a slower rate of γ-secretase cleavage progression along the transmembrane domain of βCTF allows more time for longer Aβ products to escape from the enzyme complex and the lipid bilayer before further cleavage (Figs. 1 3). For the mutations analyzed by this study, there appears to be a strong correlation between the age of disease onset and the Aβ(37+38+40)/Aβ(42+43) ratio when represented as a percentage of the wild type ratio (see Fig. 2 of the Petit et al. paper). In an earlier, 2017 paper from some of the same authors (Szaruga et al. [99], using in vitro assays involving isolated cell membranes and an antibody-tagged form of βCTF) changes in Aβ product ratios are attributed to the tendency of the mutations to destabilize γ-secretase-substrate interactions (as measured by the change in Aβ fragment formation with temperature) and so the mutations can be viewed as exhibiting loss-of-function. Interestingly, neither the Szaruga et al. or Petit et al. papers report analysis of any of the 12 PSEN1 gain-of-function mutations found by Sun et al. actually to increase total Aβ production (as measured by combined Aβ40 and Aβ42 production).
Supplementary Data File 1, Spreadsheet 1, shows a simplistic re-examination (without statistical corrections) of the data presented by Petit et al. to compare the choice of Aβ “product line” (i.e. Aβ(37+40+43) relative to Aβ(38+42)) with the age at onset (AAO) of disease. For linear correlation it finds, r2 = 0.38 for this compared to the superior value of r2 = 0.75 for the Aβ(37+38+40)/Aβ(42+43) ratio that reflects processivity. This supports that the rate of γ-secretase processivity is more closely associated with disease severity than the choice of product line. This implies that, for presenilin mutations, the severity of fAD is more closely associated with the general degree of distortion and disruption of presenilin structure/function rather than the levels (or ratios) of any particular sizes of Aβ fragments.
A 2023 study (Liu et al. [100]) examined the production of Aβ fragments with lengths of 37, 38, 39, 40, 42, and 43 as produced by HEK293 cells lacking endogenous presenilins but transfected to express wild type, or one of 130 different fAD mutant forms, of PSEN1 (together with AβPP’s βCTF). It found a best correlation with disease AAO of r = 0.5807 (p < 0.0001) for the ratio of Aβ37/Aβ42. The correlation for the Aβ40/Aβ42 ratio was also highly significant. Analysis of each mutation was, apparently, performed only once and, interestingly, almost a third of these showed higher levels of total Aβ production (or of Aβ40 plus Aβ42 production as was reported by Sun et al. [96]) than the wild type controls (see Supplementary Data File 1, Spreadsheet 2, for an analysis of the Aβ production data in Table 1 of Liu et al. [100]). This is not consistent with the idea that presenilin mutations cause fAD by decreasing the γ-secretase cleavage of AβPP’s βCTF.
Later in this paper, we argue that the correlation between presenilin fAD mutation AAO and γ-secretase processivity is consistent with an idea that disease severity is dependent on the ability of conformational changes in presenilin holoproteins and/or γ-secretase complexes to disrupt the ability of these to form stable multimers.
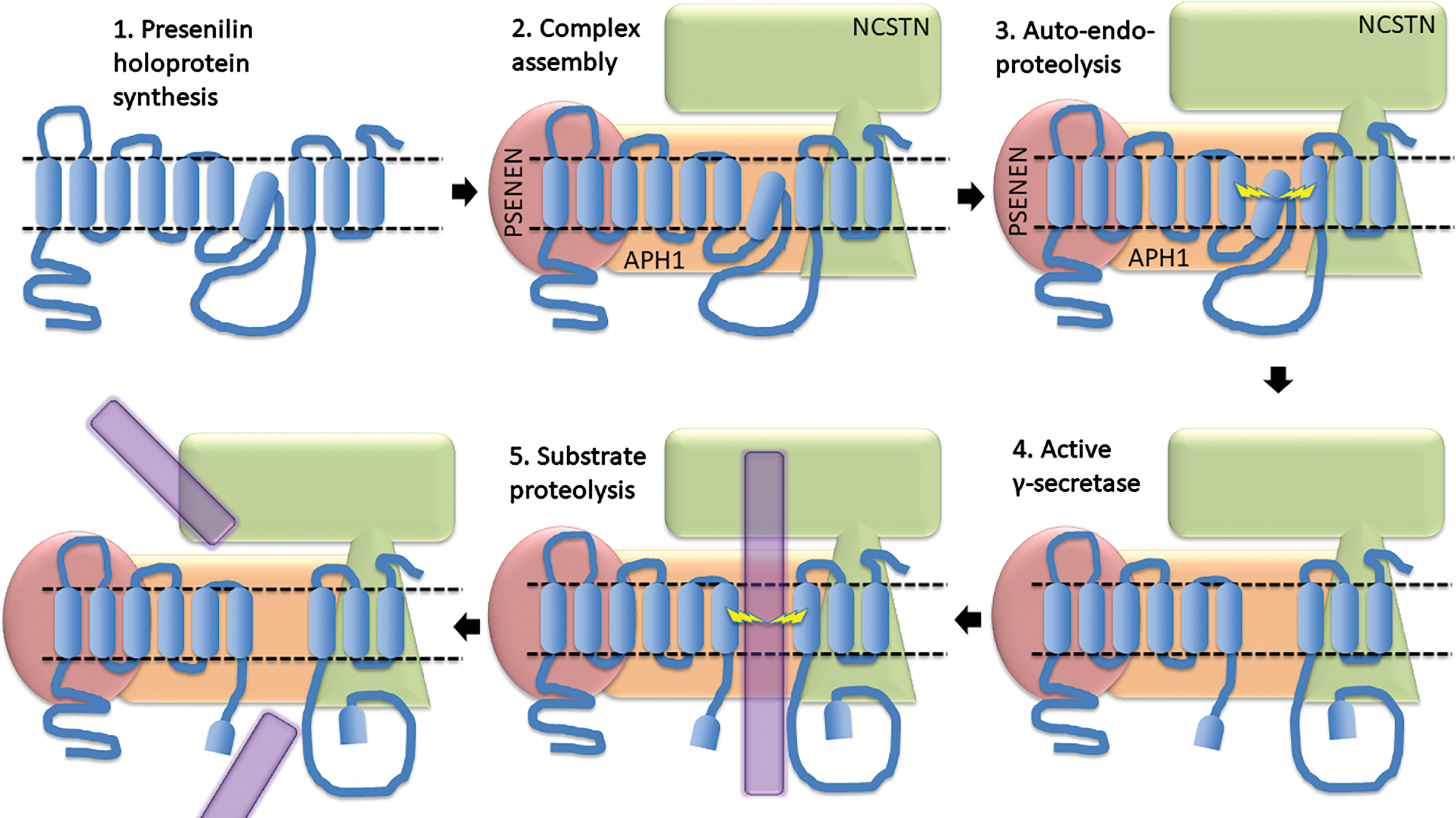
PRESENILIN HOLOPROTEINS
The presenilin proteins at the catalytic core of the γ-secretase enzyme complex may be auto-activating zymogens. Presenilin proteins are first translated as “holoproteins” that possibly cleave themselves endoproteolytically [101] to form catalytically active heterodimers consisting of the N- and C-terminal fragments (NTF & CTF) within the γ-secretase enzyme complex (Fig. 7). Some fAD mutations strongly affect this activating cleavage [102–106]. Petit et al. [98] and Szaruga et al. [99] noted that such mutations (e.g., PSEN1 mutations P433 S, R278I, and L435F) have milder clinical effects (i.e., later ages of onset) than expected from correlations between other mutants. As described below, the later-than-predicted ages of onset of these mutations that strongly inhibit γ-secretase activity are consistent with an alternative presenilin fAD mutation pathogenic mechanism.
Compared to research on γ-secretase, very little research has examined the biological role of the presenilin holoproteins or their possible role(s) in AD pathogenesis. This is likely because the holoprotein states of presenilins are commonly regarded as short-lived and transitory [107]. However, in the early days of characterization of presenilin biology, careful analysis by Dewji et al. [108] demonstrated that, when cells are subjected to gentle lysis, most presenilin protein can subsequently be detected in the holoprotein state. (This suggests the tantalizing possibility that presenilin proteins may be involved in responses to mechanical stress in addition to responding to hypoxia/oxidative stress [52]).
The most important paper demonstrating the functional uniqueness of the PSEN1 holoprotein was published in 2010 by Ralph Nixon and colleagues (Lee et al. [80]). This described an important role for the holoprotein in correct assembly of the vacuolar ATPase (v-ATPase) that pumps H+ ions into the endolysosomal system, and particularly the lysosome, for acidification. Here we see a convergence of the activities of the presenilins and the βCTF of APP, all are modulating endolysosomal pH (Fig. 8). In fact, Nixon and colleagues have commented (without yet presenting experimental data) that βCTF may bind to v-ATPase to modulate v-ATPase activity [21]. These researchers have,

“ . . . identified a direct physical interaction of lysosomal-associated APP-βCTF with the cytosol-exposed domain of the transmembrane V0a1 subunit of the vATPase. Competition by APP-βCTF with specific V1 subunits for binding to the V0a1 subunit impedes association of the V1 and V0 sectors of the complex . . . .Impaired vATPase assembly . . . is rescued by specifically lowering APP-βCTF levels”
Regardless of whether or not βCTF interacts physically with v-ATPase components, the fAD mutant forms of APP, PSEN1, and PSEN2 [109], all affect the acidification of the endolysosomal pathway (as, interestingly, does the late onset AD risk allele APOE4 [79]). Failure to acidify lysosomes sufficiently will inhibit their ability to degrade Aβ [110], causing Aβ to accumulate in lysosomes and leading, ultimately, to the formation of neuritic Aβ plaques [85]. In addition, Kim et al. [111] recently showed evidence that both Aβ and MAPT (Tau) can bind to v-ATPase components to inhibit their function. Furthermore, Mustaly-Kalimi et al. [112] recently observed decreased v-ATPase expression when cytosolic concentrations of Ca2+ ions are raised, while an earlier 2015 paper from Lee et al. [113] found that loss of lysosomal acidification leads to increased levels of cytosolic Ca2 + . These observations imply the existence of pathological positive feedback interactions between altered cytosolic Ca2+ levels and lysosomal acidification. Overall, the function of v-ATPase has emerged as a hub for the pathological influences of many agents identified as important in the development of AD.
The 2010 Lee et al paper [80] showed evidence that, in the endoplasmic reticulum, PSEN1 holoprotein interacts with the oligosaccharyltransferase complex to promote N-glycosylation of the V0a1 subunit of v-ATPase. This is necessary for correct targeting of the V0a1 subunit to the lysosome for assembly of functional v-ATPase. This PSEN1 holoprotein-dependent function in glycosylation is deficient in mouse cells lacking Psen1, but also in human fibroblasts heterozygous for fAD mutations of PSEN1. In conclusion, the numerous (and so, presumably, loss-of-function) fAD mutations of PSEN1 appear to act through a gain-of-function mechanism (see point 2 above) to cause a dominant negative (loss-of-function) effect. How is this possible? We suggest that disruption of a multimeric complex of holoprotein, or of multimers of γ-secretase, provides a parsimonious solution to this quandary.
PRESENILIN HOLOPROTEINS AND/OR γ-SECRETASE COMPLEXES FORM MULTIMERS
Biochemical evidence supports that the presenilin proteins form multimers [104, 115]. In particular, Heilig et al. [104] and Zhou et al. [97] presented strong evidence from immunoprecipitation that fAD mutant PSEN1 holoproteins can bind (directly or indirectly) to wild type PSEN1 holoproteins. Heilig et al. saw association of differentially-tagged holoproteins in cells co-transfected with expression vectors for these proteins. In contrast, Zhou et al. saw association between pre-purified γ-secretase complexes containing different mutant forms of presenilin. This raises the question of whether the multimeric association involves only the holoproteins or occurs between holoproteins in association with some or all of the other components of γ-secretase complexes. Both Heilig et al. and Zhou et al. reported dominant negative effects of the mutant proteins on the γ-secretase activity of the wild type (non-mutant) protein. However, under both experimental regimes, the relative reduction in γ-secretase activity was small (∼20%) for a 1 : 1 ratio of mutant to wild type protein resembling a heterozygous situation. This led Zhou et al to conclude,
“To gain a better understanding of the moderate dominant negative effect, the expression level of the WT and mutant PSEN1 alleles, as well as the oligomerization state of γ-secretases, should be carefully examined under in vivo circumstances, especially in patients’ brains.”
Importantly, Zhou et al. also noted,
“We wish to stress that our experimental data provide no supporting evidence for a potential role of γ-secretase in the development of AD. In fact, a number of the γ-secretase variants with pathogenic PS1 mutations, exemplified by S365A, have WT-level proteolytic activity in terms of Aβ42 and Aβ40 production (18 [i.e. see [96]]). The development of AD in the patients with these PS1 mutations cannot be explained by the WT-level proteolytic activity of these γ-secretase variants in vitro. Nonetheless, the dominant negative effect of these PS1 mutations in patients with AD still applies, suggesting such an effect may not be recapitulated by the catalytic function of PS1 in γ-secretase. We speculate that, for the vast majority of patients with AD, the dominant negative effect of PS1 is perhaps effected through other mechanisms that are independent of γ-secretase.”
It is also worthwhile noting that a super-resolution microscopy study of presenilin distribution on the surface of cells saw mainly monomers with some possible dimer formation [116]. However, this may not be true of all cellular membranes as the amounts of presenilins/γ-secretase complexes vary within cells, with particularly high levels being located in the MAM of the endoplasmic reticulum [83].
The importance of PSEN1 and AβPP’s βCTF for the acidification of the endolysosomal pathway, and the involvement of many late onset AD risk loci in endolysosomal function [77], supports that defects in endolysosomal pathway function may be a key, unifying, driver of AD pathology (Fig. 8). If all fAD mutations of the presenilins caused loss of γ-secretase activity, then this would also promote accumulation of AβPP’s βCTF to reduce lysosomal acidification and would be consistent with this unifying endolysosomal hypothesis for AD pathogenesis. However, as described above, some fAD mutations appear to increase γ-secretase cleavage of βCTF. There is evidence that correct endolysosomal pH may be important for normal γ-secretase activity [117–120] (although, as discussed by Maesako et al. [120], this conflicts with the supposed pH optimum of 6.3–7.0 for γ-secretase activity established in biochemistry-based assays). Therefore, it is conceivable that the problems with endolysosomal acidification, caused by the effect of any fAD mutation on presenilin holoprotein function, may be sufficient to decrease overall γ-secretase activity in the endolysosomal system, even for those fAD mutations that show increased γ-secretase activity in homogeneous assay systems (i.e., no presence of wild type presenilin). However, a recently discovered mutation in PSEN2 is inconsistent with this idea. Pang et al. [89] found that a mutation in the 3’ untranslated region (UTR) of PSEN2 transcripts increases the expression of PSEN2, apparently by destroying the binding site of the miR-183-5p microRNA. This mutation was strongly linked to fAD in a Chinese family lacking identifiable fAD mutations in other genes. Since no missense mutations were identifiable in PSEN2, PSEN1 or APP, an implication is that increased γ-secretase activity alone can drive AD pathogenesis.
A PARSIMONIOUS MODEL TO ACCOUNT FOR SUPERFICIALLY INCONSISTENT/CONFLICTING PRESENILIN MUTATION DATA
Of the ∼20,000 protein-coding genes in the human genome, the overwhelming majority of fAD mutations occur in only three: APP, PSEN1, and PSEN2. This restricted number of genes, and the known enzyme/substrate relationship between their protein products, supports that a single mechanism underlies fAD pathogenesis, consistent with a role for γ-secretase activity in fAD. Alternatively, protein substrates of γ-secretase are not uncommon—they are encoded by more than 1/200th of the set of all protein-coding genes [26, 27]—so it is not extremely improbable that the fAD-promoting shared activity of mutations in APP, PSEN1, and PSEN2 may be something other than effects on γ-secretase, such as reduced lysosomal acidification. In other words, while loss of γ-secretase cleavage susceptibility may be important for the action for some fAD mutations in APP, changes in γ-secretase activity in heterozygous fAD presenilin mutants might not play a role in the pathogenic activity of those presenilin mutations.
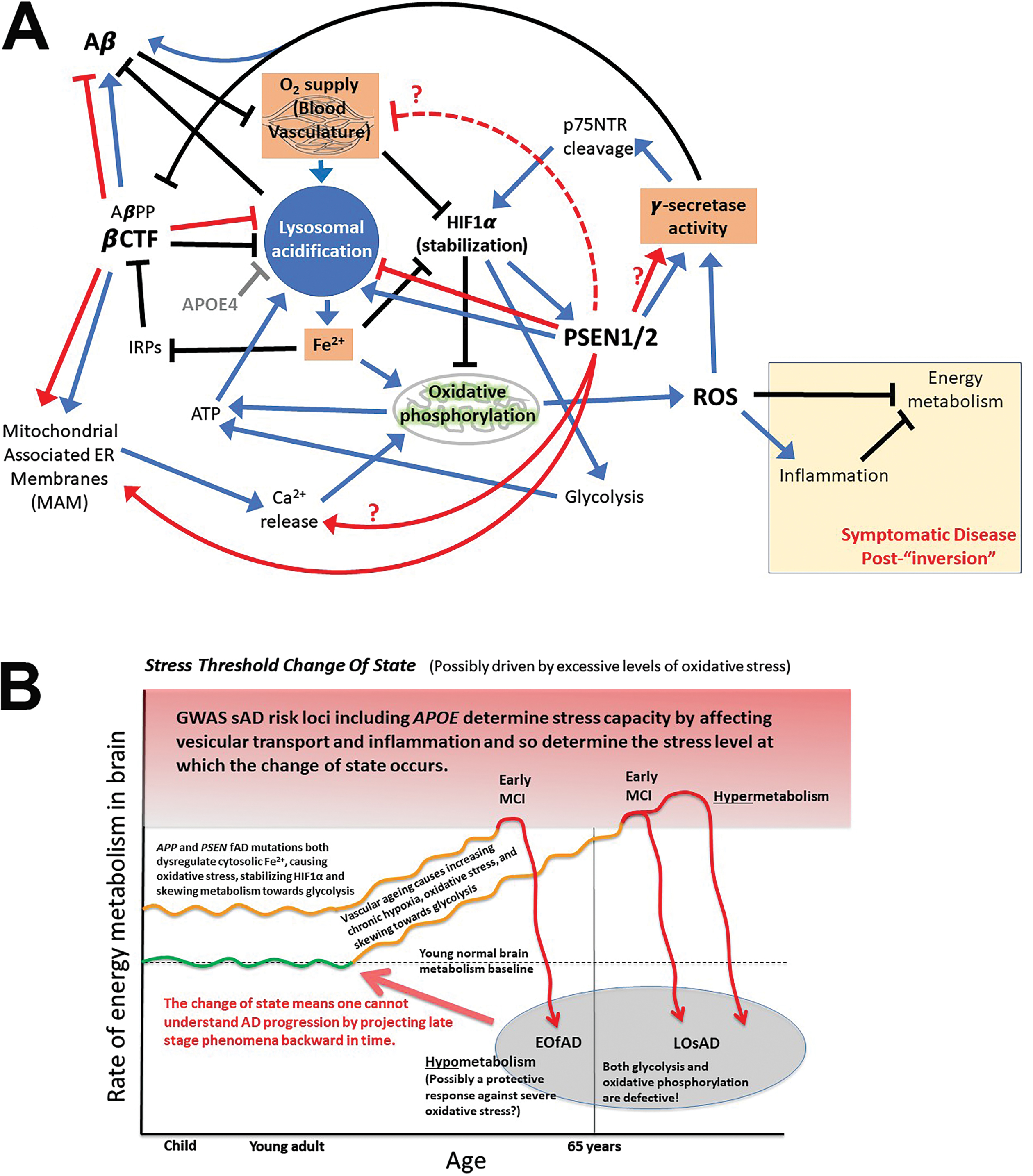
In a 2018 review examining the role of iron in AD [121], we discussed that the disease may have two distinct phases: A preclinical phase of increasing brain hypoxia with increasing reliance on glycolysis, followed by a clinical phase where cognition falters due to “metabolic inversion” of the brain into a hypometabolic state (after breaching a brain metabolic homeostasis capacity threshold - see the “Stress Threshold Change of State” model in Fig. 10B). Interestingly, γ-secretase is intimately involved in hypoxia responses. It cleaves p75NTR in a feed-forward mechanism driving stabilization of the HIF1α subunit of the transcription factor HIF1, a master regulator of cellular responses to hypoxia [122]. Importantly, γ-secretase activity is also activated by HIF1α but not via transcriptional regulation. Instead, HIF1α binds to γ-secretase [123] (possibly directly to presenilin [124]) to increase the pool of active γ-secretase [123]. In experiments using zebrafish heterozygous for a fAD-like mutation of their PSEN1-orthologous gene (psen1Q96 _ K97del), under normoxia we saw increased basal expression of genes known to be responsive to hypoxia in the brains of young adult fish that were, nevertheless, still able to upregulate these genes further under acute hypoxia. However, when the fish became aged, the response to hypoxia of these hypoxia-responsive genes became inverted; they were downregulated by acute hypoxia. This inverted response occurred at an earlier age in fAD-like mutant fish than in non-mutants, supporting that the fAD-like mutation drives accelerated aging in this aspect of brain metabolic control [125]. The observations above would be consistent with an increase in γ-secretase activity in the brains of young fish and a decrease in γ-secretase activity after an age-dependent “state change” in energy metabolism regulation.
Is there any additional evidence that fAD mutations might drive pathogenesis by first increasing γ-secretase activity before it decreases after a metabolic change of state? When we analyzed the transcriptomes of young adult zebrafish brains heterozygous for another fAD-like mutation of the zebrafish PSEN1-orthologous gene (psen1T428del, equivalent to the T440del mutation of human PSEN1), we did see overall increased expression of genes known to be upregulated by Notch signaling that is dependent on γ-secretase activity [126].
The wide distribution and missense nature of fAD mutations in the PSEN1 and PSEN2 protein-coding sequences is not consistent with the mutations affecting any particular sub-functional domain in these proteins. However, all such mutations might be expected to affect, to some degree, the conformation of the holoprotein, and that may destabilize holoprotein multimeric complexes. If multimer dissociation is required for conversion of the holoprotein into active γ-secretase then, in heterozygous cells, a fAD-mutant form of holoprotein with decreased intrinsic γ-secretase activity might, nevertheless, promote an overall increase in γ-secretase activity by causing increased activation of non-mutant holoproteins, in addition to displaying its own γ-secretase activity (see Fig. 9). Cells heterozygous for fAD mutations encoding proteins with very little intrinsic γ-secretase activity (such as the PSEN1 P433 S, R278I, and L435F mutations mentioned above) should show comparatively less total cellular γ-secretase activity than other fAD mutations, and would drive AD pathogenesis more slowly, giving their relatively later ages of disease onset as observed [98]. We suggest, therefore, that a consistent, unifying pathogenic activity of fAD mutations in PSEN1 and PSEN2 may be that these all increase γ-secretase activity in heterologous systems co-expressing both fAD mutant and wild type forms of these proteins.
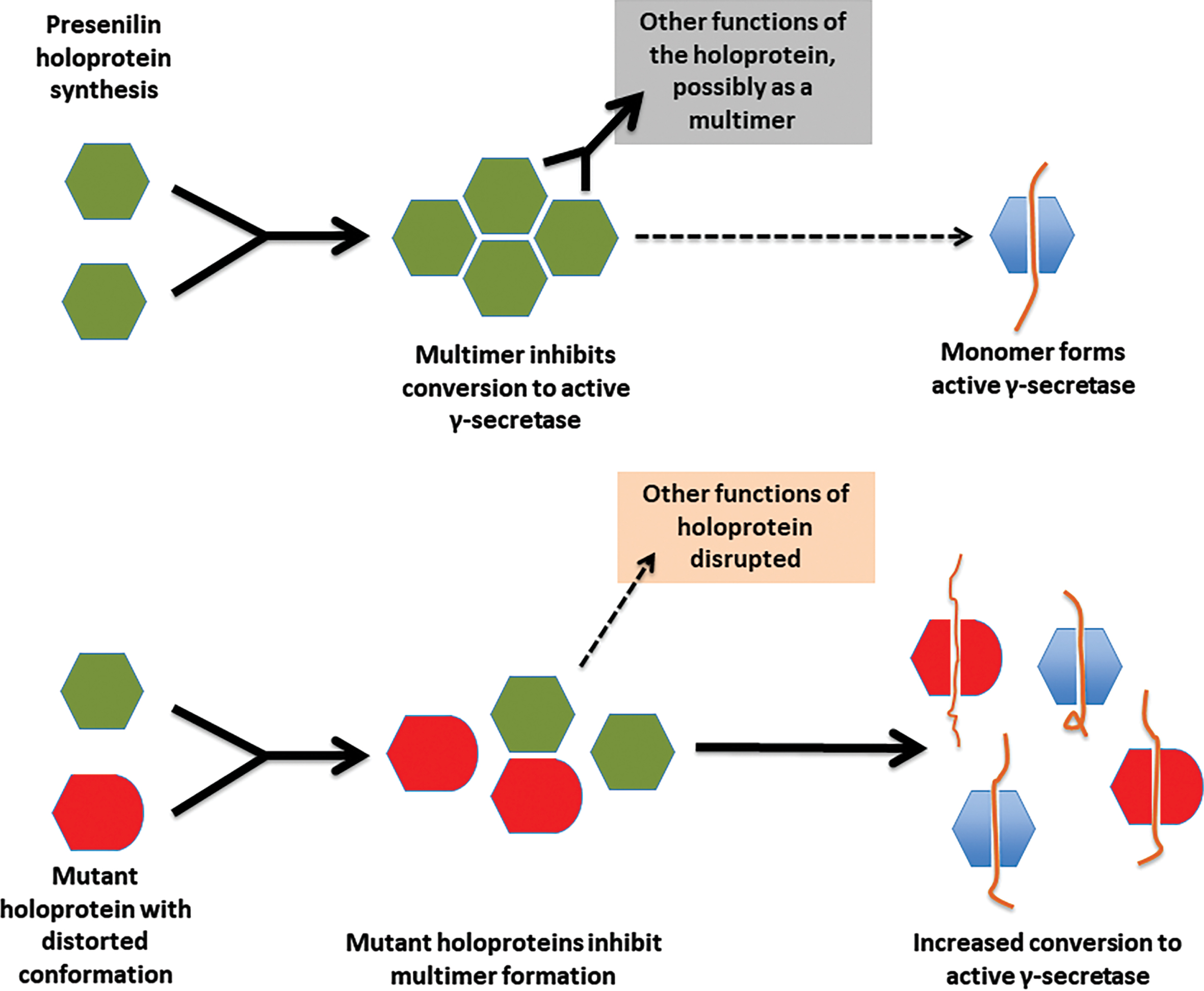
This hypothesis should be consistent with the mutation fAD onset age/Aβ fragment ratio correlation of Petit et al. [98] described previously since, for those mutations that were examined, the Aβ fragment ratio apparently reflects the degree of destabilization of the presenilin-substrate interaction (Szaruga et al. [99]) which likely corresponds to the degree of distortion of the presenilin protein within the γ-secretase complex. Any such fAD mutation might be expected to distort the conformation of the holoprotein (or an assembled γ-secretase complex) to a similar relative extent, and this should be reflected in the degree of destabilization of holoprotein multimeric complexes. We note that, for this holoprotein multimer destabilization model to function, the mutant and non-mutant forms of presenilin holoprotein must retain some ability to interact/bind each other. The N-terminal sequences of PSEN1 are known to be important for multimer formation [114, 128] and, interestingly, there is no strong evidence for fAD mutations occurring in the PSEN1 coding sequence upstream of codon 79 in the N-terminal region or downstream of codon T440 at the C-terminal end (i.e., conservation of structure in these areas may be critical for the protein interactions needed for fAD mutations elsewhere in the coding sequence to exert a pathogenic effect, see Fig. 5).
ADDITIONAL EVIDENCE CONSISTENT WITH INCREASED γ-SECRETASE ACTIVITY DUE TO fAD MUTATIONS
There are a number of curious observations that appear consistent with the idea that fAD mutations of the presenilin genes may increase γ-secretase levels. These involve the gene PSEN2.
A naturally occurring transcriptional isoform of PSEN2 exists named PS2 V [129]. This name is derived from the fact that PS2 V transcripts exclude sequences from what was once denoted as exon 5 (i.e., “V”) of PSEN2 (but is now denoted as exon 6). PS2 V levels are increased in AD brains due to an oxidative stress-sensing and -regulatory mechanism involving binding of HMGA1 protein to PSEN2 transcripts during splicing [129, 130]. Interestingly, the importance of this regulatory mechanism is demonstrated by its conservation during approximately half a billion years of vertebrate evolution, except, oddly, in mice and rats [131]. PS2 V mRNAs encode a truncated form of PSEN2 protein that cannot have any γ-secretase activity itself, but that nevertheless increases γ-secretase activity in cells [132]. This increase appears to be caused by suppression of the unfolded protein response, presumably in the ER [132–134]. (This illustrates once again the involvement of the presenilins in cellular stress responses). The existence of PS2 V is relevant to early onset AD due to the existence of the fAD mutation K115Efs of PSEN2 [41]. This deletion of two nucleotides truncates the PSEN2 open reading frame so that it would encode a protein similar to that of the truncated PS2 V isoform. Like PS2 V, the K115Efs mutant protein can also increase γ-secretase activity [53]. Note, however, that, while the K115Efs mutation was initially thought to be a possible exception to the reading frame preservation rule of presenilin fAD mutations, Braggin et al. [135] reported that this mutation also induces forms of alternative splicing that generate mRNAs with restored open reading frames but with significant coding changes. Therefore, currently, we cannot resolve whether the K115Efs mutation causes fAD through increased γ-secretase activity due to constitutive PS2V-like activity, or by encoding distorted full-length holoproteins, or both.
Another curious experimental result suggests that the most intensively investigated fAD mutation of PSEN2, N141I, shows greater than normal γ-secretase activity. Mastrangelo et al. [136] attempted to rescue embryo development in mice lacking Psen1 activity by inserting transgenes expressing either wild type human PSEN2 or N141I mutant PSEN2. Only the N141I mutant form could compensate for the absence of Psen1 activity and so allow embryo survival. Whether this superior ability of the N141I mutant is due to overall increased γ-secretase activity, or increased γ-secretase activity in a particular area of cells (e.g., due to fAD mutations changing presenilin proteins’ intracellular distributions as described by [137]), or some other mechanism is not known.
Finally, most invertebrates have only one presenilin gene but a duplication event early in the evolution of the vertebrates gave rise to PSEN1 and PSEN2. As these genes have similar structures (and, therefore, similar functions) it is unsurprising that they are both loci for fAD mutations. However, PSEN2 hosts only ∼10% as many fAD mutations as PSEN1, despite that these two genes represent almost identically sized “targets” for mutagenesis. Also, fAD mutations of PSEN2 tend to show less severity (later age of onset) than those of PSEN1 [87]. Keeping in mind that ∼10% of the fAD mutations in PSEN1 apparently increase the intrinsic γ-secretase activity of the mutant protein [96], one possibility is that only mutations in PSEN2 that increase the intrinsic γ-secretase activity of its encoded protein are sufficiently severe to cause disease. (Alternatively, since only PSEN2 protein appears to bind directly to MFN2 protein to promote MAM formation, and since the N141I fAD mutant form of PSEN2 protein was shown to bind MFN2 more avidly than wild type PSEN2 [138], it may be that, in contrast to fAD mutations of PSEN1, mutations of PSEN2 must affect both endolysosomal acidification and MAM formation if they are to cause AD—possibly a less likely event than an fAD mutation of PSEN1 affecting endolysosomal acidification alone.)
IRON AND HYPOXIA MAY BE IMPORTANT FACTORS THAT INTERACT WITH fAD MUTATIONS
Our transcriptome analyses of young adult brains from zebrafish heterozygous for fAD-like mutations in the psen1, psen2, or sorl1 genes identified oxidative phosphorylation in mitochondria as the cellular process disrupted in common between these brains [139]. Some fAD mutations may alter mitochondrial activity by affecting MAM formation or function [138]. However, Yambire et al. [140] demonstrated that disturbance of lysosomal acidification has profound effects on mitochondrial activity by affecting the availability of ferrous iron ions (Fe2+) within cells that are required by mitochondria to form the iron-sulphur clusters in mitochondrial electron transport chains (ETC) performing oxidative phosphorylation. When lysosomal acidification is reduced (i.e., pH is increased), lysosomes may accumulate poorly soluble ferric iron (Fe3+) since the conversion of Fe3+ to the more soluble Fe2+ form (that can be exported for uptake by mitochondria) is inhibited. Interestingly, neuritic Aβ plaques are complexed with iron [141] and, since Lee et al. [85] have explained how the plaques arise through accumulation of Aβ in distorted lysosomes, it is now clear how this association between Aβ and iron may occur.
Most ATP is produced through oxidative phosphorylation. Production of ATP is the most essential cellular function since most other cellular activities are dependent on the energy it provides. For example, ATP is consumed by v-ATPase to drive lysosomal acidification. However, an additional, underappreciated, but essential role of ATP is as a “hydrotrope” inhibiting aggregation of the proteins in cells that are, collectively, close to solution saturation concentration in the cytosol [142]. The generalized aggregation of proteins observed as cells age [143] probably reflects falling ATP levels together with molecular crosslinking due to rising oxidative stress (particularly if the proteins are associated with iron [141]).
Disturbance of mitochondria’s ETC, whether through deficiency of either oxygen or Fe2+, commonly generates reactive oxygen species causing oxidative stress [144]. The co-dependence of cells on oxygen and Fe2+ for ETC function is so tight that transcriptional responses to deficiencies of oxygen and Fe2+ are both regulated by the HIF1 transcription factor [145]. In fact, the effects of Fe2+ deficiency are sometimes described as “pseudo-hypoxia”. Hypoxia is thought to play a major role in the development of AD [146, 147]. The oxidative stress caused by hypoxia greatly increases the activity of the molecular machinery producing Aβ [52, 148–150] that may then be protective through its activity as an antioxidant [151–155].
Since fAD mutations in APP, PSEN1, and PSEN2 likely all inhibit correct lysosomal acidification (see Fig. 8 and previous discussion), they likely also generate pseudo-hypoxic effects by limiting cellular Fe2+ maintenance and disturbing oxidative phosphorylation (Fig. 10A). In this sense, the fAD mutations accelerate the increase in hypoxia and oxidative stress that brains normally experience during aging [156] due to the gradual degradation of vascular function [157–159] limiting the supply of oxygen and causing mitochondria to generate increased levels of reactive oxygen species [160]. Increased oxidative stress increases Aβ production at the same time as Aβ degradation is inhibited due to insufficient lysosomal acidification. Therefore, one can see rising levels of brain Aβ accumulation as a biomarker of incipient AD pathological processes [161]. At some point, the brain “shifts” into a hypometabolic state, possibly as a final effort to avoid the rising ravages of oxidative stress that also drive neuroinflammation [162] (Fig. 10B). (Notably, the increasing accumulation of Aβ plateaus as AD becomes overt [161], and the shift into hypometabolism of AD brains appears consistent with the Cell Danger Response as described by Naviaux [163, 164].)
There are many observations consistent with an overarching role for hypoxia in AD, such as the increased risk of AD due to sleep apnea [165], decreased blood flow in AD brains [166], lower levels of HIF1α in postmortem AD brains [167] and data from model organisms studied by us [125] and others (e.g., Qian et al.’s [168] demonstration that cholinergic basal forebrain neurons, that express p75NTR [169] and are selectively lost in AD [170], are particularly sensitive to degeneration in their mouse model of sleep apnea, and that this is mediated by Hif1α). An additional intriguing connection between AD and hypoxia that we have not seen emphasized elsewhere relates to the “Two-Hit” hypothesis of Alzheimer’s disease described in 2004 by Mark Smith and colleagues [43, 171]. They postulated that, “ . . . although either oxidative stress or abnormalities in mitotic signalling [aka cell cycle events, CCEs] can independently serve as initiators, both processes are necessary to propagate disease pathogenesis . . . and [are] invariant features of disease.” CCEs are seen when neurons inappropriately enter the cell cycle but do not undergo cell death. Both oxidative stress and CCEs result from hypoxia [172] with sufficient levels of HIF1α being essential for preventing inappropriate entry of cells into the cell cycle during hypoxia [173].
WHAT IS AβPP’s ROLE IN IRON HOMEOSTASIS?
The iron regulatory proteins, IRP1 and IRP2, act to maintain cellular iron homeostasis by binding to iron responsive elements (IREs) in the transcripts of genes to regulate their translation and/or stability (reviewed by [174]). The importance of iron homeostasis to cell function and survival is indicated by the fact that more than 10% of protein-coding genes may include IREs in their transcripts [175]. Considerable research has focused on the existence of an IRE in the 5’ untranslated region (UTR) of APP mRNA that inhibits translation of AβPP when cells are deficient in Fe2+ [176, 177]. A number of papers have suggested that AβPP acts to stabilize the cellular iron exporting protein ferroportin while others show data in conflict with this (discussed by us previously in [175]). The ability of β-secretase inhibitors to enhance ferroportin stability is regarded as evidence of a role for APP in stabilization of ferroportin [178]. However, this stabilization by β-secretase inhibition would also be expected if the common partner of ferroportin, the ferroxidatase hephaestin, was a β-secretase substrate. There is evidence that this is the case [179].
The fact that excessive expression of APP can cause fAD supports that iron-dependent regulation of APP mRNA translation may be a significant influence in fAD pathogenesis. We note that the IRE-dependent regulation of APP mRNA translation is consistent with a role for AβPP’s βCTF in modulating pH in the endolysosomal system. When cells are replete in Fe2+, βCTF levels would be higher since AβPP translation would not be repressed by the binding of IRP1 to the 5’ IRE in APP mRNA [177]. The consequent increase in endolysosomal pH would reduce importation of Fe3+ and its conversion to Fe2 + .
Iron is seen to accumulate in AD brains leading to the common speculation that this excessive iron load drives oxidative stress via the Fenton reaction [180, 181]. However, the mechanism described in this paper, whereby insufficient endolysosomal acidification contributes to fAD, supports an alternative idea—AD brains suffer from a functional ferrous iron deficiency in a background of ferric iron accumulation. This is supported by transcriptome analysis of postmortem AD brains indicating increased expression of iron importation protein TFRC [175], the transcripts for which are stabilized during Fe2+ deficiency through binding of IRPs to multiple IREs in their 3’ UTR [182, 183]. Recently, LeVine et al. found other transcriptome-level evidence of functional iron deficiency in a human olfactory bulb AD data set [184], while Wu et al. found an increased ratio of ferric over ferrous iron in brains of the 5xFAD model mouse, although total ferrous iron was, apparently, increased rather than decreased [185].
PREDICTIONS ARISING FROM THESE PROPOSED MECHANISMS
If fAD mutations in APP exert their effect by increasing βCTF levels, then we expect that ongoing research from the AD research community will continue to find a strong association between these levels and disease pathologies, when researchers take the care to examine βCTF levels in addition to their usual examination of Aβ levels.
If fAD mutations in APP, PSEN1, and PSEN2 all cause AD though their action on endolysosomal system acidification, then a prediction of this model would be existence of an inverse correlation between mutations’ effects on lysosomal pH and age of disease onset. The most severe mutations with the earliest ages of disease onset should show the relatively highest lysosomal pH (when comparing heterozygous mutations under uniform conditions in cellular systems with otherwise identical genetic backgrounds). Unfortunately, it is currently challenging to make precise measurements of lysosomal pH.
Testing of the hypothesis that all fAD mutations in the presenilin genes lead to increased γ-secretase activity could be performed by restoring the wild type sequence in iPSCs derived from an individual heterozygous for a fAD mutation in PSEN1 or PSEN2. Comparison of the γ-secretase activity between the mutant and the wild type cells (possessing otherwise identical genetic backgrounds) could then be performed after their differentiation to neurons or other cell types. The method used to compare γ-secretase activity in wild type versus mutant cells would need to be one that did not disrupt cellular integrity or place them under other exceptional stress.
To test the idea that brains “invert” into a hypometabolic state upon exceeding a particular oxidative stress threshold, a model organism such as the zebrafish could be exposed to varying levels of reactive oxygen species (e.g., by placement under varying degrees of hypoxia) and then metabolic states assessed via metabolomics and transcriptomics strategies.
CONCLUSIONS
The most parsimonious explanation for the many types of fAD mutation affecting APP is that these increase the expression of AβPP’s βCTF. This alters cellular cholesterol distribution and reduces lysosomal acidification (likely affecting Fe2+ ion homeostasis thus driving pseudo-hypoxia) to impact mitochondrial function. The identification of excessive βCTF as the pathogenic agent promoted by fAD mutations in APP is consistent with Hardy and Higgins’ Amyloid Cascade Hypothesis as originally published.
A unifying mechanism for fAD promotion by mutations in PSEN1 and PSEN2 is that these drive pseudo-hypoxia by reducing lysosomal acidification thereby disrupting Fe2+ ion homeostasis. The possibility exists that these mutations also promote a pseudo-hypoxic state by increasing γ-secretase activity. The formation of multimeric complexes by presenilin holoproteins and/or γ-secretase complexes may be fundamental to understanding how mutations in the presenilin genes cause fAD. Nevertheless, this phenomenon remains almost completely uninvestigated.
Hypoxia and pseudo-hypoxia cause increased production of Aβ as brains age. Failure of lysosomal acidification results in accumulation of undegraded Aβ, likely with co-accumulation of Fe3+, in the lysosomes of neurons. Upon neuronal death these are observed as neuritic plaques.
Im E, Jiang Y, Stavrides PH, Darji S, Erdjument-Bromage H, Neubert TA, Choi JY, Wegiel J, Lee JH, Nixon RA. Lysosomal dysfunction in Down syndrome and Alzheimer mouse models is caused by v-ATPase inhibition by Tyr682-phosphorylated APP βCTF. (2023) Sci Adv. 9(30):eadg1925. doi: 10.1126/sciadv.adg1925. Epub 2023 Jul 26. PMID: 37494443; PMCID: PMC10371027.
Footnotes
ACKNOWLEDGMENTS
The author wishes to thank Eric Schon, Greg Sutherland, and Giuseppe Verdile (and the anonymous peer-reviewers of this paper) for their feedback after critical reading of the manuscript. All opinions and discussion in this work are the responsibility of the author alone.
FUNDING
The author is an academic employee of the University of Adelaide.
CONFLICT OF INTEREST
The author declares no conflict of interest.