
Abstract
Background:
Neuroinflammation and oxidative stress can aggravate the progression of Alzheimer’s disease (AD). Centella asiatica has been traditionally consumed for memory and cognition. The triterpenes (asiaticoside, madecassoside, asiatic acid, madecassic acid) have been standardized in the ethanolic extract of Centella asiatica (SECA). The bioactivity of the triterpenes in different solvent polarities of SECA is still unknown.
Objective:
In this study, the antioxidative and anti-neuroinflammatory effects of SECA and its fractions were explored on lipopolysaccharides (LPS)-induced microglial cells.
Methods:
HPLC measured the four triterpenes in SECA and its fractions. SECA and its fractions were tested for cytotoxicity on microglial cells using MTT assay. NO, pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), ROS, and MDA (lipid peroxidation) produced by LPS-induced microglial cells were measured by colorimetric assays and ELISA. Nrf2 and HO-1 protein expressions were measured using western blotting.
Results:
The SECA and its fractions were non-toxic to BV2 microglial cells at tested concentrations. The levels of NO, TNF-α, IL-6, ROS, and lipid peroxidation in LPS-induced BV2 microglial cells were significantly reduced (p < 0.001) by SECA and its fractions. SECA and some of its fractions can activate the Nrf2/HO-1 signaling pathway by significantly enhancing (p < 0.05) the Nrf2 and HO-1 protein expressions.
Conclusions:
This study suggests that the inhibitory activity of SECA and its fractions on pro-inflammatory and oxidative stress events may be the result of the activation of antioxidant defense systems. The potential of SECA and its fractions in reducing neuroinflammation and oxidative stress can be further studied as a potential therapeutic strategy for AD.
INTRODUCTION
Neurodegenerative diseases are one of the biggest public health concerns affecting millions worldwide. Alzheimer’s disease (AD) is the primary cause of dementia, one of the most prominent symptoms among neurodegenerative diseases. AD, the most common age-related progressive neurodegenerative disorder, has affected more than 50 million individuals worldwide. This number is anticipated to double every five years and reach 152 million by 2050 [1]. The well-known hallmarks of AD are the formation of neurofibrillary tangles and amyloid plaques, which can cause neurodegeneration, neuroinflammation, oxidative stress, and injury of cholinergic neurons [2].
Neuroinflammation refers to a brain injury that induces an inflammatory response, leading to the activation and build-up of microglial cells and astrocytes at the injury sites. Therefore, microglial activation is one of the fundamental pathology hallmarks of AD development. There are two phenotypes of microglial activation: M1 pro-inflammatory and M2 anti-inflammatory phenotypes, which will produce both pro-inflammatory and anti-inflammatory mediators, respectively. When an injury happens, the M1-activated microglia may predominate at injury sites, whereas M2-activated microglia may appear later as they are associated with anti-inflammatory tissue repair processes [3, 4]. The M2 microglia uses coordinated modulation of anti-inflammatory mediators to reduce the M1 pro-inflammatory responses, producing immunosuppressive and neuroprotective benefits. Microglia can move from one phenotype to another in regulating the neuroinflammatory state to avoid or delay the onset of AD.
Lipopolysaccharide (LPS)-induced microglial cells are often used in vitro studies on neuroinflammation-related diseases [5–7]. When activated, microglial cells increase the expression of nuclear factor-κB (NF-κB), which causes pro-inflammatory proteases such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX2) to become active. Pro-inflammatory mediators like nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin (IL)-6, and IL-1β are eventually released. Overproduction of these pro-inflammatory mediators causes neurodegeneration or even mortality, hastening the start and progression of neurodegenerative disorders [8]. Regulating activated microglial cells may thus be a promising therapeutic target for neuroinflammation-related neurodegenerative disorders.
The neurotoxic amyloid-β (Aβ) peptides and the intracellular accumulation of neurofibrillary tangles of hyperphosphorylated tau protein, which are the neuropathological diagnostic criteria for AD, are among the primary causes of oxidative stress [9, 10]. Any injury or insult that results in the activation of microglial cells leads to the events of oxidative stress. Oxidative stress is a biological state caused by an imbalance in the generation of reactive oxygen species (ROS) and the cellular antioxidant defense response [11]. The ROS can react with lipids, proteins, nucleic acids, and other molecules and alter their structures and activities [12–14]. Besides, a previous study has demonstrated that lipid peroxidation products, including malondialdehyde (MDA), 4-hydroxynonenal, and F2-isoprostanes had been observed to be elevated in several brain areas and cerebrospinal fluid of patients with AD or mild cognitive impairment [14, 15].
Multiple antioxidative systems fight oxidative stress and inflammation in the normal state. Nuclear factor erythroid 2-related factor 2 (Nrf2) is a crucial transcription factor that mitigates oxidative stress. Under normal conditions, Nrf-2 binds to Kelch-like ECH-associated protein 1 (Keap1) and is continually destroyed. Under stressful conditions, however, it dissociates from Keap-1 and translocates to the nucleus, boosting the gene expression of anti-inflammatory mediators [16]. In addition, Nrf-2 promotes the production of heme-oxygenase 1 (HO-1) [17]. HO-1, a heat shock protein, possesses antioxidative and anti-inflammatory properties. It has been demonstrated that raising HO-1 activity improves inflammatory response while suppressing it worsens inflammatory injury [18]. As a result, the Nrf-2/HO-1 signaling pathway is an essential therapeutic target for efficiently treating neurodegenerative disorders caused by inflammation.
At present, only symptomatic relief treatments are being prescribed to AD patients, which primarily enhance neuronal transmission and restore adequate levels of acetylcholine in the brain. Based on the WHO’s recommendation, developing countries are encouraged to utilize traditional herbal medicine as a part of the treatment. The WHO estimates that nearly 80% of the global population uses traditional medicines as their primary health treatment [19, 20]. Thus, the focus of researchers has shifted to the utilization of traditional herbal medicine.
Centella asiatica (L.) Urban belongs to the Apiacaea family, known as pegaga in Malay or Indian pennywort in English, and is a perennial creeping herbaceous plant [21–23]. It is traditionally used as a medicinal herb to treat wounds, various skin conditions, diarrhea, fever, amenorrhea, and diseases of the female genitourinary tract, as well as for anxiety relief and improving cognition [24]. Traditional medicines, particularly Centella asiatica (CA) as total plant extracts or its active compounds, have shown various beneficial pharmacological activities in both in vitro and in vivo models. For instance, madecassoside, asiaticoside, and asiatic acid have been derived from CA, as well as whole plant extract have demonstrated the ability to decrease the cell apoptosis, oxidative stress, and pro-inflammatory markers in in vitro AD models, including LPS-induced BV2 microglial cells [25–27], SH-SY5Y neuroblastoma cells [28], Aβ-induced brain microvascular endothelial cells [29] and glutamate-induced HT22 hippocampal cells [30]. Furthermore, CA whole extract and its derived active compounds, such as asiaticoside and asiatic acid, have shown neuroprotective effects against various in vivo AD models, which include chronic stress-induced and cognitively impaired rats’ models [28, 30–33]. Although the efficacy and usefulness are still unknown, medicinal herbs are preferred since they have fewer adverse effects. However, the information on the comprehensive mechanism of action of these medicinal herbs on the human body is still lacking, thus limiting its clinical application [34].
The major group of bioactive compounds available in CA is pentacyclic triterpenoids, which are abundantly found as saponins, asiaticoside, and madecassoside, and their aglycones, asiatic acid and madecassic acid [23]. These four major triterpenes are believed to be active bioactive compounds that play important roles in exhibiting all the potential biological activities. Previous studies demonstrated that these triterpenes, alone or combined, exhibited various biological activities, including neuroactive and neuroprotective properties. For instance, both asiatic acid and asiaticoside exhibited an anxiolytic effect on rat models with no sedative effects [35–37]. In addition, asiaticoside, madecassoside, and asiatic acid have neuroprotective effects in models of stroke, including lower levels of pro-inflammatory cytokine and microglial activation [38–40]. Hence, the triterpenes of CA are among the bioactive compounds responsible for exhibiting the neuroprotective effects.
MATERIALS AND METHODS
Reagents and chemicals
Analytical grade reagents and chemicals were used in the in vitro assays. The reagents and chemicals were purchased from Sigma-Aldrich, Thermo Fisher Scientific, and Nacalai Tesque. The BV2 microglial cell line was purchased from the American Type Culture Collection (ATCC, USA).
Preparation of standardized extract of Centella asiatica (SECA) and its four different fractions
CA extract was prepared by Jusril’s group [41], where the CA plants were collected from Herbagus Trading, Pulau Pinang, Malaysia. The sample was deposited in the Faculty of Applied Sciences, UiTM Shah Alam (Selangor, Malaysia) under the voucher number CA-K017. The whole plant was washed, cleaned, and oven-dried at 40°C. The powdered CA leaves were extracted with 95% denatured ethanol at room temperature for 72 h. A concentrated liquid extract was obtained using a rotary evaporator and subsequently freeze-dried to be designated as SECA [28]. The SECA was then fractionated by the solvent partitioning method. SECA was reconstituted in a ratio of 9 : 1 mixture of 95% denatured ethanol and deionized water. Three different polarities of solvents were selected—namely hexane, dichloromethane, and ethyl acetate—to partition SECA into individual fractions, while the remaining organic layer was considered a water fraction. All fractions formed from the organic layer were collected and followed by evaporation by a rotary evaporator. The ethyl acetate and water fractions underwent an additional freeze-drying process using a freeze dryer. A flow chart in Supplementary Figure 1 details the extraction and fractionation process.
High-performance liquid chromatography (HPLC)
Waters HPLC system was used as an HPLC instrument. Analysis was accomplished over a 2696 separation module and 2996 PDA detection with a reverse phase mode Thermo Hypersil GOLD C18 column (4.6 mm150 mm; 5μm). The elution conditions were as follows: flow rate, 1.0 mL/min; column temperature, 30°C; injection volume, 15 mL; and diode-array UV-vis detection at a wavelength of 206 nm. The solvent system used was water and acetonitrile using a gradient elution program for 55 min, as described in [41] with a slight modification (Table 1).
Gradient condition of HPLC
Cell culture
BV2 microglial cell line was maintained in Dulbecco’s modified essential medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin/streptomycin in a humidified incubator of 5% CO2 at 37°C. The spent media were changed with fresh media every other day. Once the cells achieved at least 70% confluency, they were harvested through trypsinization to detach the cells from the flask. The cells were subcultured once every three days.
Cell viability assay
Following cell culture, a cell density of 1×104 cells/well, in 180μL of complete media, of BV2 microglial cells was plated into a sterile 96-well microplate with a lid. After 24 h of incubation, the cells were treated with 20μL of fresh media alone, 0.1% dimethyl sulfoxide (DMSO), SECA, and its four fractions at four different concentrations (0.1μg/mL, 1μg/mL, 10μg/mL, 100μg/mL). The cytotoxicity test was tested in two different time points: 24 and 48 h incubation. After 24 or 48 h of incubation, the cells’ viability was measured by using the 3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide (MTT) reagent. A volume of 50μL of 2 mg/mL MTT reagent was added into each well containing the cells prior to 4 h incubation at 37°C. After the incubation, the supernatant in each well was discarded. A volume of 100μL of DMSO was added into each well to dissolve the formazan crystals produced by metabolically active cells. The intensity of color formed corresponds to the total viable cells in the well. The absorbance was measured at a wavelength of 550 nm. The percentage viability of the cells was calculated by comparing them with the negative control.
Measurement of nitric oxide level
To measure the NO level, a total volume of 180μL of media containing BV2 microglial cells was plated into a sterile 96-well microplate at a cell density of 1×104 cells/well. After 24 h incubation, the cells were pre-treated with a total volume of 20μL fresh media alone, 0.1% DMSO, 250μM of L-NG-Nitro arginine methyl ester (L-NAME) or SECA and its four fractions at different concentrations (0.1μg/mL, 1μg/mL, 10μg/mL, 100μg/mL). After 24 h incubation, 22.2μL of LPS at a final concentration of 1μg/mL was added into each well containing pre-treated cells. The Griess reagent, comprising 0.01% N-1-napthylthylenediamine dihydrochloride (NED) and 1% sulphanilamide in 5% phosphoric acid, was prepared and mixed thoroughly. After another 24 h incubation with LPS, a total volume of 50μL of the supernatant from each well was transferred into another 96-well plate and incubated with 50μL of Griess reagent for 5 min at room temperature. Absorbance was measured at a wavelength of 540 nm by using a microplate reader. The color intensity produced corresponds to the total NO level in the supernatant formed by the cells and is expressed in fold change against the negative control.
Measurement of pro-inflammatory cytokines levels
The BV2 microglial cells were plated into sterile 24-well tissue culture plates at 1×105 cells/well in 900μL of complete media and incubated for 24 h. Then, the cells were pre-treated with 100μL fresh media alone, 1μM of Dexamethasone, or SECA and its four fractions at four different concentrations (0.1μg/mL, 1μg/mL, 10μg/mL, 100μg/mL). Then, after 24 h, the cells were induced with 111.1μL of LPS at a final concentration of 1μg/mL. After 24 h incubation, the spent media in each well were collected and centrifuged at 1000 rpm for 5 min. The supernatants were then collected. The levels of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) concentrations in the culture supernatants were measured by using ELISA kits (TNF-α and IL-1β: R&D systems; IL-6: Elabscience), according to the manufacturer’s instructions. The color intensity of each experimental well of the experiment was determined at 450 nm wavelength. The unknown concentration of the inflammatory cytokines was calculated by comparing with the standard curve obtained for each experiment.
Measurement of intracellular ROS level
To measure the intracellular ROS level, a total volume of 180μL of media containing BV2 microglial cells was plated in a sterile black 96-well plate with a clear flat-bottom at a cell density of 1×104 cells/well. After 24 h incubation, the cells were pre-treated with a total volume of 20μL fresh media alone, 25μM of quercetin or SECA and its four fractions at four different concentrations (0.1μg/mL, 1μg/mL, 10μg/mL, 100μg/mL). After 24 h incubation, a total volume of 22.2μL LPS at a final concentration of 1μg/mL was added into each well containing pre-treated cells. The spent media were discarded after another 24 h incubation of LPS. Each cell-containing well was rinsed twice using PBS. A total volume of 100μl of DCFH2-DA with a concentration of 20μM [42] was added to each well prior to 1 h incubation at 37°C. Then, the DCF intensity was measured using a fluorescence microplate reader at wavelength excitation of 485 nm and emission of 535 nm. The color intensity produced corresponds to the amount of ROS production by the cells in each well and is expressed in fold change against the negative control.
Lipid peroxidation assay
MDA, an oxidative stress marker, is one of the final products of lipid peroxidation in the cells. An increase in oxidative stress results in overproduction of MDA levels. For the determination of MDA level in the BV2 microglial cells, the MDA assay kit (Abcam, Cambridge, UK) was used. A total volume of 1800μL media containing BV2 microglial cells was plated in a sterile 6-well plate with a lid at a cell density of 5×105 cells/well. After 24 h incubation, the cells were pre-treated with a total volume of 200μL fresh media alone, 25μM of quercetin or SECA and its four fractions at different concentrations (10μg/mL and 100μg/mL). After 24 h incubation, 222.2μL of LPS at a final concentration of 1μg/mL was added into each well containing pre-treated cells. The spent media were discarded after 24 h incubation of LPS. The cells were then harvested, washed with PBS, and homogenized using the provided lysis buffer in the kit. The resultant cell supernatant was used for measurement of MDA with the Lipid Peroxidation (MDA) Assay Kit (Colorimetric/Fluorometric) (Abcam, USA) in accordance with the manufacturer’s instructions.
Measurement of Nrf2 and HO-1 protein expression
Western blot is used to identify the antibody specificity in a complex biological sample by the detection of a single band at a distinct molecular weight [43]. The conditioned medium produced by microglial cell lysates was collected. The lysate was collected as follows: After administering LPS on the pre-treated cells for 24 h, the cells were washed twice with ice-cold PBS and harvested with a cell lysis buffer containing protease inhibitor. Then, the total protein concentration was quantified using the bicinchoninic (BCA) assay. Two sets of gels were prepared: resolving and stacking gels for the sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). Upon completion of the transfer of the proteins to a piece of PVDF membrane, the membrane was blocked in 5% bovine serum albumin (BSA) in TBS-T (Tris-Buffered Saline containing 0.05% of Tween 20) at room temperature for 1 h on an orbital shaker at 30 rpm. The membrane was incubated overnight in 10 mL of diluted primary antibody (Nrf2 and HO-1) at 4° C on an orbital shaker at 30 rpm. The monoclonal Nrf2 (Cell Signalling Technology; D1Z9C, 1 : 1000 dilution), HO-1 (Cell Signaling Technology; E9H3A, 1 : 1000 dilution), and GAPDH (Cell Signalling Technology; D4C6R, 1 : 5000 dilution) were used. Prior to immunodetection, the membrane was rinsed with wash buffer (TBS containing 0.05% of Tween 20, TBS-T) on an orbital shaker at 50 rpm for 5 min. The membrane was then incubated in 10 mL of diluted secondary antibody (Cell Signalling Technology; #7074, 1 : 5000 dilution) at room temperature for 1 h on an orbital shaker at 30 rpm. The bands of interest were analyzed using the NIH Image J software. The pixel values of the digitized bands were measured to determine the protein expression of GAPDH, Nrf2 and HO-1. All proteins of interest were normalized with the loading control protein, and the basal level was adjusted to 1.0.
Statistical analysis
All results obtained were expressed in mean±S.D of three or five replicates. The groups were compared and analyzed using one-way ANOVA using GraphPad Prism (version 8) before Dunnett’s post-hoc test for the statistical significance determination. p values were considered significant if < 0.05.
RESULTS
Quantification of madecassoside, asiaticoside, madecassic acid, and asiatic acid in SECA and its fractions
The HPLC identification and quantification of four different triterpenes compounds in SECA and its four different fractions were done by assessing their retention time (RT) and ultraviolet-visible (UV-VIS) spectra at 206 nm compared with the standards reference. Figure 1 represents the chemical profiles of SECA. The chemical profiles of the fractions and reference standards of madecassoside, asiaticoside, madecassic acid, and asiatic acid are shown in Supplementary Figure 2. In SECA, the content of madecassosides was found to be the highest (336.89±3.22 mg/g) compared to the other triterpenes compound, which is similar to the results obtained in the previous study done by Jusril’s group (2021). For the fractions of SECA, there were different chemical profiles between each fraction. Only 0.16±0.02 mg/g of asiatic acid can be found in the hexane fraction, whereas this compound was more abundant in the DCM fraction, 77.80±1.80 mg/g. Other than SECA, all four triterpenes’ compounds were found in both DCM and EA fractions. Meanwhile, in water fraction, only madecassosides and asiaticoside were found at 37.33±0.45 mg/g and 17.78±0.25 mg/g, respectively. The results are presented in Table 2.

Chromatogram of SECA with four detected reference standards: (1) madecassoside (MD); (2) asiaticoside (AS); (3) madecassic acid (MDA); (4) asiatic acid (ASA).
Triterpenes compounds in SECA and its fractions; hexane, DCM, EA, and water fractions. Concentration of triterpenes contents are expressed in milligram/gram extract/fractions
SECA, standardized extract of Centella asiatica; DCM, dichloromethane; EA, ethyl acetate.
Cytotoxicity profiles of SECA and its fractions on BV2 microglial cells
Cytotoxicity profiles of SECA and its fractions were evaluated at two different time points: 24 and 48 h. After 24 and 48 h of treatment of SECA and its fractions at various concentrations (0.1–100μg/mL), the cell viability of the BV2 microglial cells was evaluated by MTT assay. This assay was carried out to assess the cytotoxicity activity of SECA and its fractions against the BV2 microglial cells. Mitochondria in metabolically active cells reduce MTT to form water-insoluble dark purple formazan; thus, the total amount of formazan produced is directly proportional to the number of viable cells [44]. The results are shown in Table 3, Columns 1 and 2, and Supplementary Figures 3 and 4. Based on the results, all treatments, regardless of the concentration given to cells, showed insignificant changes compared to the negative control (C + M) at both 24- and 48-h incubation. This experiment suggests that SECA and its fraction have a non-toxic effect on the BV2 microglial cells at concentrations up to 100μg/mL. Hence, in the following experiments, the BV2 microglial cells were pre-treated for 24 h, and the treatment will last up to 48 h with the additional 24 h LPS induction prior to the continuation of respective functional assays.
A series of in vitro functional assays to determine the effects of SECA and its fractions—hexane (HEX), dichloromethane (DCM), ethyl acetate (EA), and water—with a concentration of 0.1, 1, 10, and 100μg/ml, in BV2 microglial cells. Positive control was differently used in each assay: Cell Viability assay (without LPS stimulation) – untreated cells, NO assay and Cell Viability assay (with LPS stimulation) – L-NAME (250μM), ELISA – dexamethasone (1μM) and ROS assay – quercetin (25μM)
ap < 0.05 compared to negative control, bp < 0.05 compared to LPS, cp < 0.01 compared to LPS, dp < 0.001 compared to LPS.
Suppression of nitric oxide level by SECA and its fractions on LPS-induced BV2 microglial cells
NO is one of the pro-inflammatory mediators, which can be tested its activity in vitro by performing a Griess assay. Hence, this assay was done to evaluate the suppressive effect of SECA and its fractions against the release of NO in LPS-induced BV2 microglial cells. The results are shown in Table 3, Column 4 and Supplementary Figure 5. LPS significantly increased the production of NO as compared to the negative control (C + M). L-NAME, a non-selective NOS inhibitor, acts as a positive control for this assay and significantly suppresses the production of nitric oxide induced by LPS on BV2 microglial cells. All SECA and its fractions at the highest concentration (100μg/mL) showed a suppressive effect on NO production induced by LPS on BV2 microglial cells compared to the LPS control results. Each extract/fraction showed a dose-dependent suppression of NO production. Interestingly, at the lower concentration of 10μg/mL, SECA and its hexane and dichloromethane fractions significantly suppressed NO production induced by LPS on BV2 microglial cells compared to the LPS control. MTT assay was carried out concurrently with the Griess assay to confirm that the suppressive effect produced was from the treatment itself, not due to low cell viability after treatment (Table 3, Column 3, and Supplementary Figure 6). It showed that the cell viability results at all treatment concentrations were statistically insignificant compared to negative control(C + M).
Suppression of pro-inflammatory cytokines by SECA and its fractions on LPS-induced BV2 microglial cells
BV2 microglial cells induced by LPS will produce and secrete pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, which can be detrimental to the neuronal cells. This study assessed the inhibitory effects of SECA and its fractions on the production of these pro-inflammatory cytokines using ELISA.
TNF-α
The result is shown in Table 3, Column 5, and Supplementary Figure 7. LPS significantly increased the production of TNF-α as compared to the negative control (C + M), approximately 48-fold. DEXA significantly reduced the production of TNF-α as compared to the LPS control, approximately 3-fold. Only SECA, hexane, and DCM fractions at the highest concentration (100μg/mL) showed a significant inhibitory effect on the production of TNF-α induced by LPS on BV2 cells, approximately by 2.7-, 3.6-, and 2.4-fold, respectively as compared to the LPS control results. Compared with DEXA, the SECA, hexane, and DCM fractions at 100μg/mL concentrations produced a similar suppressive effect.
IL-6
The result is shown in Table 3, Column 6, and Supplementary Figure 8. LPS significantly increased the production of IL-6 compared to negative control (C + M). DEXA significantly reduced the production of IL-6 as compared to the LPS control. Only SECA, hexane, and DCM fractions at the highest concentrations (100μg/ml) showed a significant inhibitory effect on the production of IL-6 induced by LPS on BV2 cells compared to the LPS control results. Also, hexane fraction at 10μg/ml significantly reduced the production of IL-6 compared to the LPS control cells. Compared with DEXA, the SECA, hexane, and DCM fractions at 100μg/mL concentrations produced a similar suppressive effect.
IL-1β
The result is shown in Table 3, Column 7, and Supplementary Figure 9. All values were below zero (negative values), which indicates that the protein could not be detected. No trend can be seen between different concentrations of the treatments.
Inhibition of intracellular ROS production by SECA and its fractions on LPS-induced BV2 microglial cells
2’, 7’– dichlorofluorescein diacetate (DCFH2-DA) assay was performed to determine the inhibitory effect of SECA and its fractions on the production of intracellular ROS in LPS-induced BV2 microglial cells. Mechanistically, the DCFH2-DA is diffused into the cells. Then, the acetyl group of DCFH2-DA is cleaved off by cellular esterase, which then forms a highly fluorescent DCF molecule. DCF emits green fluorescence, which can be detected by fluorescence spectroscopy at an excitation/emission wavelength of 485 nm/530 nm. The results on the level of intracellular ROS obtained are shown in Table 3, Column 8 and Supplementary Figure 10. LPS significantly increased the production of ROS as compared to the negative control (C + M). All SECA and its fractions at the highest concentration (100μg/ml) showed a significant inhibitory effect on the production of ROS induced by LPS on BV2 cells compared to the LPS control results. Also, SECA and hexane fraction at 10μg/ml and DCM fraction at 1 and 10μg/ml significantly reduced the production of ROS as compared to the LPS control. Meanwhile, quercetin, as a positive control of this experiment, showed an insignificant reduction in fold change of ROS level.
Inhibition of lipid peroxidation products by SECA and its fractions on LPS-induced BV2 microglial cells
Lipid peroxidation is one of the oxidative stress markers that can be determined by evaluating the level of its final product, MDA. The MDA produced in the samples binds with thiobarbituric acid (TBA), forming the MDA-TBA adduct. This MDA-TBA adduct is easily quantified at a wavelength of 532 nm. Based on results obtained in DCFH2-DA assay experiments, a few concentrations of SECA and its fractions, which significantly inhibited the production of intracellular ROS in the previous experiment, were selected for this MDA assay experiment. As shown in Fig. 2, LPS significantly increased the production of MDA as compared to negative control (C + M) by approximately 3-fold. As a positive control in this experiment, Quercetin was significantly reduced, approximately by 2.5-fold, the MDA production compared to the LPS control. The selected concentration of SECA and its hexane and dichloromethane fractions at 100μg/ml concentration was significantly reduced, by approximately 2.4-fold, the production of MDA in LPS-induced BV2 microglial cells compared to the LPS control. Meanwhile, at the concentration of 10μg/ml, SECA and its fractions were significantly reduced, by approximately 1.3-fold, the production of MDA in LPS-induced BV2 microglial cells compared to the LPS control. Hence, all SECA and its fractions at selected concentrations showed an inhibitory effect on the production of MDA induced by LPS on BV2 microglial cells compared to the LPS control results.
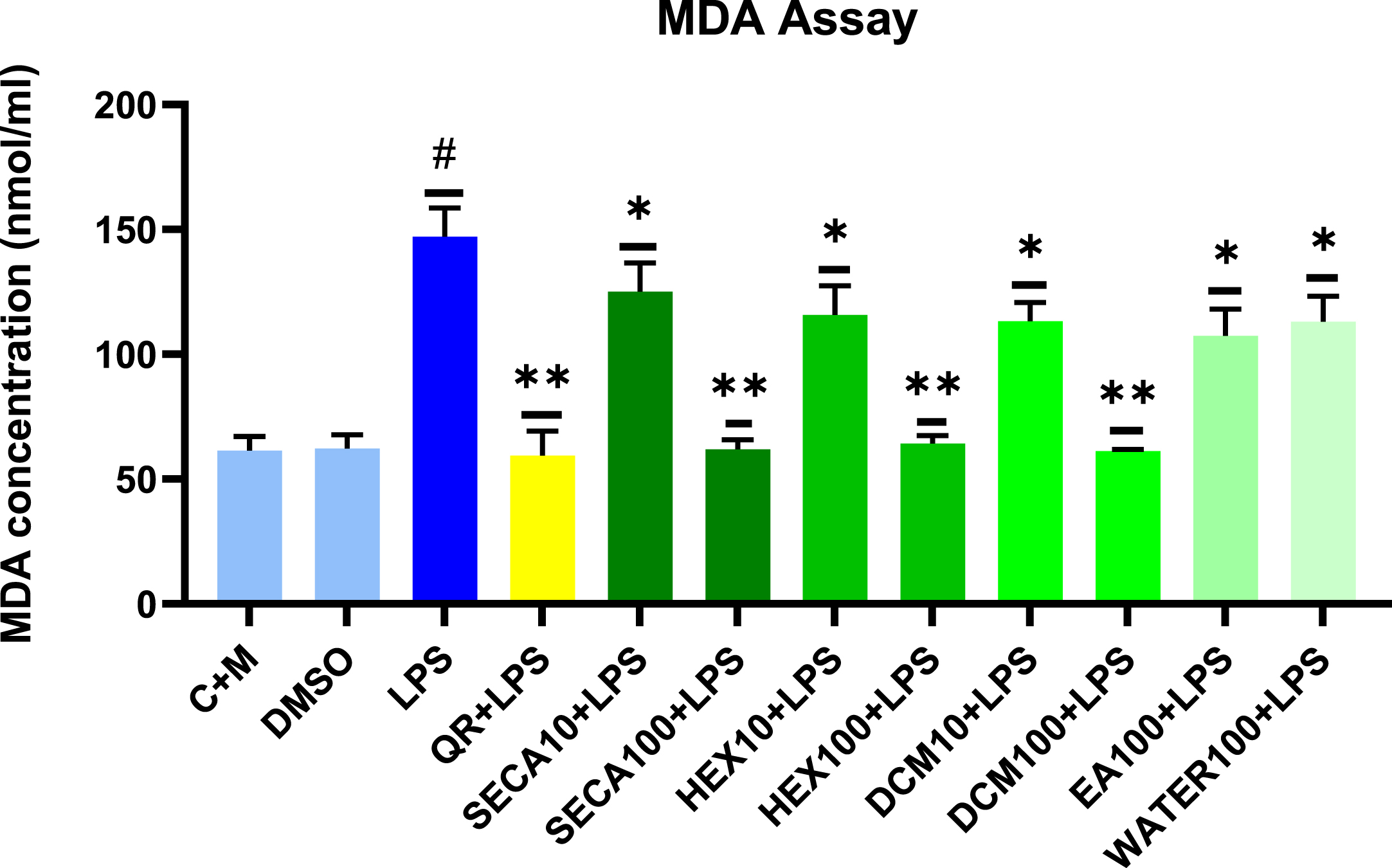
Effects of SECA and its fractions on lipid peroxidation by assessing the level of MDA produced in LPS-induced BV2 microglial cells. The MDA levels are expressed as mean±S.D. (n = 3). The extract showed dose-dependent inhibition of lipid peroxidation product, MDA release. The values are statistically significant (#p < 0.05) compared to non-treated cells (C + M), *p < 0.05 and **p < 0.01 when compared with LPS control cells.
Enhancement of antioxidant protein markers of SECA and its fractions on LPS-induced BV2 microglial cells
Western blot was carried out on LPS-induced BV2 microglial cells, which were pre-treated with SECA and its fractions at a constant concentration to determine their effects on the antioxidant markers (Nrf2 and HO-1). A 100μg/mL concentration was selected as it exhibited the most potent biological activity for SECA and most of its fractions in previous experiments while possessing a non-toxic effect on the BV2 microglial cells. GAPDH, a loading control protein, was used to normalize the total protein in the sample loaded in each well. As shown in Fig. 3, all pre-treatment of SECA and its fractions on LPS-induced BV2 microglial cells upregulated the expression Nrf2 compared to the negative control (C + M). The protein expression of Nrf2 and HO-1 was slightly increased even with LPS alone. However, the expression of LPS-induced BV2 microglial cells pre-treated with SECA and its fraction was much higher than those with LPS alone. In addition, there was significant upregulation of Nrf2 expression level in SECA- and hexane fraction-treated groups. Meanwhile, for HO-1 expression, only the hexane fraction-treated group was significantly upregulated. As a positive control, Dexamethasone showed an upregulation of the expression of Nrf2 and HO-1; however, it was statistically insignificant (Fig. 4).
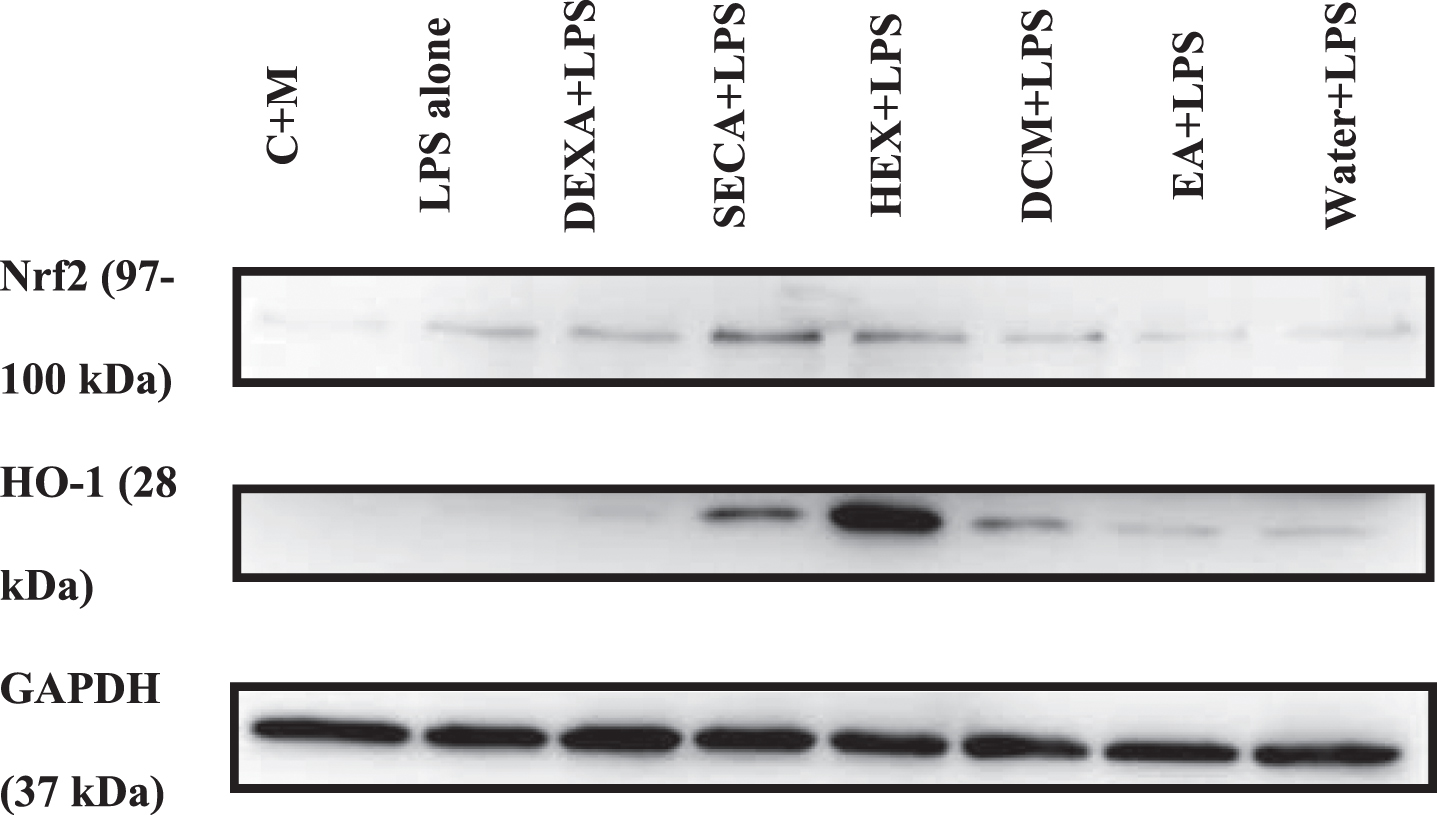
Representative images of western blotting of Nrf2/HO-1 signaling protein expression in total protein lysates of BV2 microglial cells. The concentration used for SECA and its fraction was 100μg/mL. GAPDH was used as a loading control.

Semi-quantitative analysis of (A) Nrf2 and (B) HO-1 proteins. Protein bands were quantified using ImageJ software. All values were normalized with GAPDH. Data presented as mean±S.D. (n = 5). The values are statistically significant, *p < 0.05, **p < 0.01, and ***p < 0.001, compared to non-treated cell (C + M).
DISCUSSION
In the present study, SECA-K017—an ethanolic extract of CA—was selected among other extracts designated by Jusril’s group (2020). This SECA-K017 was extracted from one of the CA accessions found in Perlis, Malaysia known as pegaga kampung, which is characterized by its light green and crenate with dentate-based leaf and has a glabrous stem. A previous study has shown that this extract possesses the highest anti-acetylcholinesterase (AChE) activity, which was believed to be employed by the presence of asiatic acid and madecassic acid. These two bioactive compounds showed a promising AChE inhibitory profile as very good interactions with the active sites were demonstrated and fulfilled the docking parameters against AChE [41]. A similar extract has also been utilized in a previous study by Hafiz’s group (2018). They demonstrated that this extract exhibits anti-acetylcholinesterase, anti-inflammatory, and antioxidative stress against LPS-induced neuroinflammation in rat models [28]. Therefore, a further study on this extract was carried out to explore the potential neuroprotective effects and its mechanisms, which may be beneficial as an alternative treatment for cognitive function and in developing a novel and effective AD treatment.
Triterpenes, alkaloids, and flavonoids are plant-derived secondary metabolites that have been demonstrated to provide therapeutic benefits for treating cancer, cardiovascular disease, skin illnesses, and neurological disorders [45]. It is believed that triterpenes in CA may be one of the potential inhibitors against AD. A previous study showed that the composition of triterpenes is influenced by plant location, where a constant fingerprinting analysis is required to standardize herbal plants in developing phyto-drugs or functional foods. This may also increase the yield of active phytoconstituents, resulting in optimum triterpenes employed as biomarkers [41]. It is known that every bioactive compound has its respective polarity index, which can be dissolved and extracted into a similar or closely related polarity index [46, 47]. In a previous study, SECA was demonstrated to be an active crude extract in some biological activities. However, a specific bioactive compound(s) at which the polarity index is responsible for the biological activities remains unclear. In the present study, SECA was fractionated into four different polarity indices of solvents, where a specific amount of soluble phytochemical dissolves into a highly specific solvent, resulting in a different composition of triterpenes in each fraction. HPLC analysis demonstrated that the detection of four triterpenes in SECA was in line with Jusril’s group [41], where both madecassoside and asiaticoside are the most abundant triterpenes found in the extract, followed by its aglycones, madecassic acid, and asiatic acid. However, the total concentration of each compound in the present study was slightly different in comparison to Jusril’s HPLC analysis results. Despite being a similar accession of SECA, the differences were still present due to several factors, including seasonal, geographical conditions, climate, harvesting times, and storage conditions [48, 49].
As expected, the composition of these four major triterpenes differed in each fraction, where only a small quantity of asiatic acid, not the other three triterpenes, can be detected in the hexane fraction. This finding suggests that among four triterpenes, asiatic acid is the most non-polar, yet not a truly non-polar compound, as only 0.1624±0.02 mg/g was found in the hexane fraction. Meanwhile, both dichloromethane and ethyl acetate fractions had all four triterpenes, but in a different quantity, where madecassic acid and asiatic acid were more abundant in dichloromethane fraction as compared to madecassoside and asiaticoside. Meanwhile, madecassoside and asiaticoside were more abundant in ethyl acetate fraction than madecassic and asiatic acid. This phenomenon happens due to the nature of the compounds, which can be determined by their molecular structures. Both madecassoside and asiaticoside compounds have additional sugar structures (glucose-glucose-rhamnose), which increase the polarity index of the compounds [50], resulting in higher amounts found in solvents with a higher polarity index, such as ethyl acetate.
The activation of microglia can occur when there is injury or stress, which results in increased production of various mediators depending on the polarization phenotype of the activated microglia. Previous studies have demonstrated that the presence of activated microglia influenced the progression of AD [51]. Therefore, in this study, LPS-induced BV2 microglial cells were utilized as an in vitro AD model, representing the brain’s activated microglia. The BV2 microglial cells are immortalized cells, which have similar functions as primary microglial cells, such as motility, activation mediators, and releasable factors, including cytokines and NO, and 90% of similar genes induced in response to LPS [52]. Thus, they are considered an appropriate alternative to the primary microglial cells despite the upregulation of genes in BV2 microglial cells being far less noticeable [53, 54]. In the present study, all SECA and its fractions were initially assessed against BV2 microglial cells to determine their cytotoxicity activity using an MTT assay. All tested extracts and fractions of SECA at any concentration (ranging from 0.1–100μg/mL) exhibited non-cytotoxic effects towards the cells after 24- and 48-h exposures, as the percentage viabilities of the cells were statistically not significant when compared to the non-treated cells. In the preliminary experiment of the MTT assay that we conducted using different time points (24, 48, 72, and 96 h), the cells were observed to develop a stress morphology and begin to die off after 72 h. In line with the previous studies, the ethanolic extracts of CA were non-toxic against a wide range of cultured cells, including SH-SY5Y neuroblastoma, RAW 264.7 macrophage [28, 56], BV2 microglia [25], and BJ fibroblast [57] cell lines. Besides, there is limited literature on the adverse effects and toxicity of CA. CA is safe to consume with a recommended dosage, and the side effects are uncommon. However, high doses may cause headaches, upset stomach, nausea, dizziness, and drowsiness [58]. The most common daily dose of CA was reported to be 600 mg of dried leaves or infusion, single-dose capsules (ranging from 300–680 mg, thrice a day), and a capsule form of 10 mg concentrated extract. A standardized CA extract containing up to 100% total saponins (triterpenoids) can be consumed at 60 mg once or twice daily [24]. Hence, this finding strengthens the potential of CA as an alternative medicine, as it is safe for consumption.
Once the microglial cells are activated, pro-inflammatory mediators and cytotoxic factors such as iNOS, TNF-α, IL-1β, IL-6, and COX-2 are being overly produced at the local injury sites [59]. NO is a crucial, highly reactive chemical involved in the inflammatory cascade, whereas the enzyme iNOS is essential for NO production from the L-arginine. NO inhibits ATP synthesis in neural mitochondrial metabolism, enhancing ROS formation [60]. In the present study, SECA and its fractions were assessed for their suppressive activity against LPS-induced BV2 microglial cells on the production of pro-inflammatory mediators, including NO, TNF-α, IL-6, and IL-1β. The current study showed that LPS-induced BV2 microglial cells significantly increased the production of mediators, such as NO, TNF-α, and IL6, among the downstream proteins in the neuroinflammation cascade. Remarkably, pre-treatment with SECA and its fractions at various concentrations between 0.1 to 100μg/mL significantly suppressed NO production compared to the non-treated cells in a dose-dependent manner. Both hexane- and dichloromethane fractions showed a similar pattern as SECA-treated cells, where at 100μg/mL, the release of NO was suppressed almost to the basal level and maintained a high cell viability percentage, suggesting that a low level of NO releases by the same quantity of the viable cells, not due to low cell viability. Meanwhile, the suppressive activity of SECA’s ethyl acetate- and water fractions was lower than SECA and its hexane- and dichloromethane fractions. However, at the highest concentration, they significantly suppressed NO production and release by approximately 30% compared to the non-treated LPS-induced BV2 microglial cells. Previous studies have demonstrated similar results in suppressing NO production by natural products’ extracts in LPS-induced BV2 microglial cells. Among the natural products that have been tested against a similar in vitro AD model as current study are Ficus deltoidea aqueous extract [42], Atractylodis Rhizoma Alba ethanolic extract [61], aryl benzofuran isolated from Dalbergia odorifera, Curcuma longa and its main compound, curcumin [7, 63], and Menthae Herba ethanolic extract [64]. In addition, the CA as a whole extract or its derived compound, asiatic acid, also showed a suppressive effect in LPS-induced BV2 microglial cells [25, 26]. The results obtained from previous studies were consistent with the current study, in which natural products, particularly CA-containing bioactive compounds, exhibited a suppressive effect of NO production on LPS-induced BV2 microglial cells. These findings suggest that SECA and its fractions can be a potent anti-inflammatory agent for the activated microglial cells by suppressing NO production.
Moreover, the production of TNF-α and IL-6 in LPS-induced BV2 microglial cells was significantly increased, and pre-treatment with SECA, hexane- and dichloromethane fractions managed to reverse the effects. Pre-treatment of LPS-induced BV2 microglial cells with SECA and its hexane- and dichloromethane fractions significantly attenuated the production of these two cytokines at the highest concentration, 100μg/mL. Consistently, previous studies have shown that the CA extracts and their bioactive compounds inhibited the production of TNF-α and/or IL-6 in LPS-induced BV2 microglial cells. The CA ethanolic extract has significantly attenuated the production of TNF-α, approximately 3.5-fold, with a concentration of 100μg/mL [25]. Similarly, both asiatic acid and madecassoside, bioactive compounds of CA, showed a significant, dose-dependently, attenuation of TNF-α and IL-6 production as well as their mRNA expression in LPS-induced [26] and oxygen-glucose deprived [65] BV2 microglial cells. Nevertheless, in the present study, ethyl acetate- and water fractions of SECA containing asiatic acid and madecassoside failed to attenuate the production of TNF-α and IL-6 in activated BV2 microglial cells despite the treatment of the bioactive compound alone has been proven in attenuating the production and expression of TNF-α and IL-6. Various factors can contribute to the failure of the attenuation of TNF-α and IL-6 production in the current study, including the solvent used to extract the compounds. It should be noted that an indirect correlation is reflected in the presence of certain bioactive compounds to biological activities. Although fewer biological activities were observed in ethyl acetate and water fractions, the bioactive compounds found in the fractions still can be active on their own. To some extent, the different mixtures of bioactive compounds may produce synergistic, additive, or antagonistic effects [66]. Hence, the present study analyzed the effects of the total bioactive compounds on biological activities solely based on their contents, regardless of the potential biological activity of each bioactive compound or the interaction between other bioactive compounds. Presumably, another bioactive compound(s) might be presented in the SECA and its hexane- and dichloromethane fractions, which possess a greater anti-inflammatory activity.
In previous studies, the CA extract and its bioactive compounds were proven to attenuate the production of pro-inflammatory cytokines, including IL-1β, in BV2 microglial cells. In contrast, in the present study, the IL-1β produced by LPS-induced BV2 microglial cells failed to be detected using a highly sensitive assay, ELISA. The inadequate signal peptide of IL-1β makes this secreted protein have a different secretion pathway. A previous study has demonstrated that IL-1β released by the activated immune cells needs a secondary stimulus to shed the microvesicles containing the bioactive IL-1β through lysosomal exocytosis [67, 68]. An efficient release of the bioactive form of IL-1β requires extracellular ATP binding to the P2X7 receptors, which are highly expressed in immune and epithelial cells [69]. Similarly, BV2 microglial cells, the immune cells in the brain, may follow the secretion pathway of IL-1β. However, in the present study, no additional stimulus was given to the supernatant of the spent media prior to measuring IL-1β via ELISA. This absence may affect the results as the processing and release of IL-1β are inefficient in the absence of a secondary stimulus, with most of the newly synthesized products remaining intracellular and unprocessed or degraded [67]. Hence, the present study could not evaluate the effect of SECA and its fractions on the production of IL-1β in LPS-induced BV2 microglial cells.
An overstimulation of inflammatory response promotes ROS generation, and oxidative stress stimulates microglial activation. Oxidative stress and excessive inflammatory response may be the primary causes or aggravators of neurodegenerative diseases such as AD [5]. Therefore, in the present study, LPS has been used to activate the BV2 microglial cells, eventually stimulating oxidative stress events. The level of oxidative stress in LPS-induced BV2 microglial cells was evaluated by assessing the production of intracellular ROS via DCFH2-DA assay. The results showed that LPS significantly increased the production of ROS in BV2 microglial cells by approximately 1.3-fold compared to the non-induced BV2 microglial cells. This result was consistent with the results of ROS production in previous studies that have stimulated the BV2 microglial cells with LPS [25, 64]. The increase in intracellular ROS was attenuated with the pre-treatment of SECA and its fractions in a dose-dependent manner. The previous studies have demonstrated that the production of intracellular ROS by LPS-induced BV2 microglial cells was significantly attenuated by various natural products, including Vignae Radiatae Semen ethanolic extract [5], Menthae Herba ethanolic extract [64], Ficus deltoidea aqueous extract [42], as well as CA extract [25] and its bioactive compound, madecassoside [27]. In the present study, the intracellular ROS production was significantly attenuated by pre-treatment of LPS-induced BV2 microglial cells with the highest concentration (100μg/mL) of SECA and its fractions. Meanwhile, only SECA and its hexane- and dichloromethane fractions could attenuate intracellular ROS production at a lower concentration–10μg/mL. These findings suggest that SECA and its fractions can be a potent antioxidant agent for the activated microglial cells by attenuating the production of intracellular ROS.
When oxidative stress occurs, the ROS attacks cellular membranes, resulting in functional and/or structural degradation of the membranes and the generation of hazardous lipid products such as 4-hydroxy-2,3-nonenal, MDA, acrolein, and F2-isoprostanes. MDA, one of the most extensively investigated aldehydes, is related to numerous clinical diseases, including AD [70]. In the present study, the LPS treatment significantly increased the production of MDA in BV2 microglial cells, which was evaluated by measuring the MDA-TBA adduct formation using an MDA assay. This result supports the previous ROS experiment, where LPS increased the production of ROS in BV2 microglial cells, which increased lipid peroxidation. The pre-treatment of LPS-induced BV2 microglial cells with SECA and its fractions significantly decreased lipid peroxidation, reflected in MDA production, in line with the previous studies [71, 72]. However, the pre-treatment of SECA and its fractions, particularly hexane- and dichloromethane fractions, at the highest concentration was identified as the most potent in lowering lipid peroxidation by significantly attenuating the production of MDA. A study by Khan et al. (2019) utilized antioxidant curcumin to inhibit the LPS-induced oxidative stress in BV2 microglial cells by reducing the production of MDA. Until now, limited studies have evaluated the effect of CA extract and/or its bioactive compounds on the production of lipid peroxidation markers, particularly in LPS-induced BV2 microglial cells. The results obtained in the present study suggest the potential of SECA and its fractions as a neuroprotective agent against oxidative stress.
An antioxidant transcription factor, nuclear factor-erythroid 2-related factor 2 (Nrf2), induces the expression of heme oxygenase-1 (HO-1) to maintain intracellular redox homeostasis and neuroinflammation. Nrf2 suppresses neuroinflammation as a secondary outcome of antioxidants, inhibiting pro-inflammatory and oxidative stress mediators [73]. The ROS activates the Nrf2 signaling pathway by dissociating Nrf2 from its inhibitor, Keap1. Nrf2 is normally linked to the cytosolic repressor Kelch-like ECH-associated protein 1 (Keap1) and labelled for polyubiquitination-mediated proteasomal destruction. Growing evidence suggests that Nrf2 is expressed in neurons, astrocytes, and glial cells [74]. However, during neuroinflammation and/or oxidative stress, Keap1 releases Nrf2, which subsequently translocates into the nucleus and binds to antioxidant response regions of the genome and small Maf proteins. This results in the production of antioxidant genes and phase II enzymes such as reduced nicotinamide adenine dinucleotide phosphate (NADPH), reduced glutathione, superoxide dismutase, catalase, and HO-1, all of which have neuroprotective effects on neuronal cells [75]. Besides, Nrf2 has been reported to inhibit LPS-induced expression of pro-inflammatory cytokines through the Nrf2/HO-1 signaling pathway [73, 77]. Therefore, in the present study, to confirm the effects of the Nrf2/HO-1 signaling pathway on the suppressive activities of pro-inflammatory and oxidative stress mediators by the SECA and its fractions against LPS-induced BV2 microglial cells, the protein expression of Nrf2 and HO-1 were evaluated. The antioxidant transcription factor Nrf2 plays a crucial role in regulating LPS-TLR4 signaling, potentially through the modulation of oxidative stress. In this study, LPS stimulation alone increased the expression of Nrf2 and HO-1 proteins in BV2 microglial cells. This result indicates that LPS activated the Nrf2 signaling pathway, which supports the previous literature that demonstrated an upregulation of antioxidant proteins in Nrf2 wild-type models as opposed to the Nrf2-knockout cells and rat models upon treated with LPS [78]. In addition, an intervention with bioactive compounds in the previous studies has effectively reduced the inflammatory response induced by LPS [64, 77–80]. Similarly, this study showed that SECA and its hexane fraction pre-treatment significantly upregulated Nrf2 expression for 24 h. This result was in line with the previous studies that demonstrated the upregulation of Nrf2 proteins with the treatment of CA in diabetic kidney disease [81] and age-related macular degeneration [82] rats’ models. This result further supports the important role of Nrf2 in regulating LPSsignaling.
The treatment of dexamethasone has been demonstrated to stimulate the movement of Nrf2 from the cytosol to the nucleus. This translocation results in the continuing accumulation of Nrf2 in the nucleus for up to 24 h following drug administration. This sustained accumulation of Nrf2 is responsible for promoting the expression of phase II antioxidant genes [83, 84]. In a preliminary study, a ROS assay was conducted with dexamethasone as a positive control along with different concentrations of SECA (ranging from 0.1–100μg/ml) prior to the western blot experiment. The results showed a significant inhibition of intracellular ROS production with the pre-treatment of dexamethasone (data not shown). Hence, we used dexamethasone as a positive control in the western blot experiment. Indeed, there was an upregulation of Nrf2 and HO-1 protein expression with the pre-treatment of dexamethasone; however, it was not statistically significant. This could be due to the activation of glucocorticoid receptor signaling by dexamethasone, which can repress Nrf2-mediated cytoprotection from oxidative stress [85, 86].
Nonetheless, only the pre-treatment hexane fraction of SECA has significantly upregulated the HO-1 expression. The present results suggest that hexane fraction activates the Nrf2 signaling pathway faster than SECA within 24 h as the downstream protein expression, HO-1, was higher with the pre-treatment of hexane fraction. Nrf2 translocation may be followed by Nrf2 phosphorylation because of SECA/other fractions-induced Keap1 degradation, masking the Nrf2 nuclear localization signal. Because translocated Nrf2 has transcriptional activity, it may boost HO-1 expression [64]. The polarity of the compounds in the hexane fraction may contribute to the bioactivity exhibited by the fraction. The non-polar and hydrophobic compound is more permeable than the polar and hydrophilic compound due to the structure of the phospholipid bilayer of the cells. Therefore, it explains the effectiveness of hexane fraction in exhibiting its biological activities compared to other fractions. Among four triterpenes, asiatic acid is the only triterpene compound detected in the hexane fraction, which was in a small amount. Interestingly, some previous studies have demonstrated the effects of asiatic acid on the Nrf2/HO-1 signaling pathway. The asiatic acid significantly upregulated the expression of both Nrf2 and HO-1 in in vitro t-BHP-induced HepG2 cells [87] and in vivo LPS-induced acute kidney injury broiler model [88]. Although asiatic acid has been proven to have a protective effect by activating the Nrf2/HO-1 signaling pathway, the effect of hexane fraction containing asiatic acid on this pathway remains unclear. It is postulated that the presence of another bioactive compound(s) could be responsible for activating the Nrf2/HO-1 signaling pathway in the hexane fraction. Based on the chromatogram of hexane fraction in Supplementary Figure 2B, there are two unidentified peaks at retention times of 18.94 and 19.399 min, which is worth exploring further and might be the bioactive compounds of hexanefractions.
To support the present results, several previous studies utilized CA extract and its triterpenes, such as asiaticoside and asiatic acid, in both in vitro and in vivo neuroinflammation models. The CA extract improved the behavior, oxidative damage and neuroinflammation effects in the sleep deprivation-induced anxiety mice model [89]. The administration of CA at 150 and 300 mg/kg doses for eight days resulted in significant enhancements in locomotor activity and anti-anxiety effects. In addition, CA reduced biochemical markers of oxidative stress and neuroinflammation marker (TNF-α) levels compared to the sleep-deprived group deprived of sleep for 72 h [89]. Similarly, a standardized CA extract, ECa233, significantly reduced the microglial activation in the chronic inflammatory temporomandibular disorder pain mice model [90]. Furthermore, asiaticoside significantly ameliorated the memory deficits in the neuroinflammation mice models induced by Aβ1 - 42 [91] and transient cerebral ischemia and reperfusion [38]. These studies proved that asiaticoside exhibited anti-inflammatory effects, which inhibited microglial activation by reducing the production of neuroinflammatory mediators such as TNF-α, IL-β, and IL-6 [38, 91]. It is worth noting that previous studies utilized asiatic acid to similar neuroinflammation in vitro model as in the present study. A treatment with asiatic acid compound against LPS-induced BV2 microglial cells had significantly reduced the pro-inflammatory mediators, including NO, iNOS, TNF-α, IL-β, and IL-6 [26, 92], which was in line with the results obtained in the present study. Both in vitro and in vivo models in the previous studies that utilized CA extract, and its triterpenes support the results obtained in the present study, in which CA has remarkable neuroprotective effects despite the exact mechanisms yet to be discovered.
Other than these four triterpenes identified within the CA, several bioactive compounds exhibited antioxidative and anti-inflammatory effects that have been studied previously. For example, a compound named araliadol from a fraction of CA extract exhibited protective effects against oxidative stress in glutamate-induced HT22 cells. The treatment with araliadol demonstrated a significant reduction in ROS production of glutamate-induced HT22 cells and protected HT22 cells from tunicamycin-induced cell death [30]. Besides, the caffeic acid has been detected within CA, in which the administration of caffeic acid resulted in cognitive-enhancing effects in cerebral ischemia-reperfusion injury in rat models. Specifically, the animals treated with caffeic acid exhibited reduced memory deficits, decreased levels of pro-inflammatory markers and lipid peroxidation, increased activity of antioxidant enzymes, and decreased hippocampal cell injury and death [93]. In addition, the flavonoids such as rutin, kaempferol, and quercetin that can be found in CA possess antioxidative and anti-inflammatory effects. A study of these flavonoids derived from CA demonstrated an inhibition of induction of matrix metalloprotease-9 in H2O2-induced human dermal fibroblasts. The flavonoids in the CA extract are believed to be the bioactive compounds that exhibited the antioxidative effects in the study [57]. Quercetin was used as a positive control in our research in some assays, exhibiting a significant antioxidative effect against LPS-induced BV2 microglial cells.
In the context of SECA, this extract has been utilized in both in vitro and in vivo in various neurodegenerative disease models by other research groups. This extract has been shown to lower the acetylcholinesterase activity in LPS-induced neuroinflammation in rat models [28, 41]. Furthermore, this extract has been demonstrated to have a neuroprotective effect against chronic stress-induced rats. The treatment of SECA resulted in a significant enhancement in the thickness of the dentate gyrus and a decrease in the occurrence of neuronal cell death in both the dentate gyrus and CA3 regions, as well as a reduction in the expression of the c-fos protein [31]. Interestingly, the study also demonstrated a significant increase in various metabolites (lactate, isoleucine, proline, methionine, valine, leucine, and glutamine) in rats subjected to SECA treatment. These metabolites of significance are known to fulfil crucial roles in various metabolic functions of the brain, including protein synthesis, energy metabolism, neurotransmitter synthesis, defense against oxidative stress, and compartmentalization of glutamate [94]. In another study by Hafiz et al. (2018), the same extract was used for LPS-induced inflammation in rat models. They demonstrated that SECA exhibited the ability to suppress AChE, level of oxidative stress and neuroinflammation [28]. Besides, a previous study by Chiroma’s group demonstrated that SECA improved cognitive abilities in D-gal- and AlCl3-induced rats with cognitive deficits. The SECA that they used was capable of: 1) restoring the cholinergic dysfunction by lowering AChE level, 2) reducing oxidative stress by lowering MDA levels and increasing superoxide dismutase levels, and 3) ameliorating the cognitive impairment by inhibiting aberrations of ultrastructural morphological of neurons in rats’ prefrontal cortex [33]. In the recent study, done by Mansor et al. (2023), SECA exhibited its effect in reducing intracellular ROS activity in H2O2-damaged stem cell-derived neurons, thereby enhancing their viability upon SECA treatment. Besides, neurite outgrowth of these neurons was also significantly enhanced by SECA treatment, with increased expression of neuronal protein markers, specifically MAP2, and antioxidant genes—Trx1 and HO-1 [95]. These previous studies support the results obtained in the present study that SECA have the potential to be a future treatment and management for neurodegenerative disease like AD.
Despite having numerous pharmacological benefits, the exact mechanism of CA in exhibiting antioxidative and anti-inflammatory properties remains unclear. A study on the time-dependent Nrf2 signaling could be advantageous as the expression of Nrf2 protein might be upregulated at different times with different treatments. Furthermore, some other signaling pathways must be involved in the mechanism of SECA and its fractions exhibiting antioxidative and anti-inflammatory properties. For instance, in the present study, Nrf2 and HO-1 proteins were significantly upregulated only in SECA and/or hexane fraction-treated cells but not in DCM fraction-treated cells despite it also resulted in a significant reduction in antioxidative and anti-inflammatory mediators production. Although dealing with this complex extract will be challenging since its multiple components may simultaneously influence multiple signaling pathways, it is nevertheless worthwhile to conduct a time-dependent future study with some other signaling pathways that may beinvolved.
CONCLUSION
The current study highlights that CA has the potential to be an anti-neuroinflammatory and antioxidant agent by regulating the Nrf2/HO-1 signaling pathway in vitro. The presence of four triterpenes, madecassoside, asiaticoside, madecassic acid, and asiatic acid, in the SECA and its fractions may contribute to its beneficial effects. Based on the study findings, SECA and its fractions were not toxic within the range of tested concentrations (0.1 to 100μg/mL), which is an initial step towards determining its potential application in an in vivo setting. In addition, SECA and its fractions, at their highest concentration (100μg/mL), exhibited anti-neuroinflammatory and antioxidant properties by attenuating the expression levels of pro-inflammatory and oxidative stress mediators, including NO, TNF-α, IL-6, ROS, MDA, Nrf2, and HO-1 in LPS-induced microglial cells via respective in vitro assays. Although the anti-neuroinflammatory and antioxidant potential of CA has been reported in previous studies, the results of this study have strengthened the use of SECA and its fractions as an anti-neuroinflammatory and antioxidant agent for the modulation of oxidative stress associated with inflammatory responses. To date, limited studies utilize CA extract and/or its fractions, specifically targeting the Nrf2/HO-1 signaling pathway in a neuroinflammation model. Thus, the antioxidative and anti-neuroinflammatory effects of SECA and its four fractions shown in the present study provide an initial insight into the effectiveness of the bioactivity of bioactive compounds in each fraction.
Previous studies demonstrated that neuroinflammation and oxidative stress induced by Aβ plaques and tau hyperphosphorylation are key factors in the pathogenesis of AD. The formation of Aβ aggregation and tau proteins are mediated by pro-inflammatory cytokines and signaling pathways induced by microglial activation. Therefore, the downregulation of these pro-inflammatory and oxidative stress mediators via activating the antioxidant signaling pathway may directly lower oxidative stress and neuroinflammation. It is safe to conclude that SECA and its fractions, particularly hexane and dichloromethane, are a potential anti-neuroinflammatory and antioxidant agent for treating and protecting against AD. Nonetheless, further studies are needed to understand the detailed composition of SECA and its fractions, particularly hexane fraction, which exhibited a remarkable bioactivity among others. It is worth to explore further on the other possible molecular mechanisms of SECA and its fractions involving other related signaling pathways such as mitogen-activated protein kinase, mammalian target of rapamycin, and phosphoinositide 3-kinases/protein kinase B signaling pathways, which eventually leads to the activation of NF-κB signaling pathway.
Footnotes
ACKNOWLEDGEMENT
The authors have no acknowledgement to report.
FUNDING
The study was funded by the Ministry of Higher Education under Fundamental Research Grant Scheme (FRGS/1/2019/SKK08/UPM/02/5) and Universiti Putra Malaysia under Putra Grant (GP/2017/9566200).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings on this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.