
Abstract
Background:
Alzheimer’s disease (AD) is the most common cause of dementia and remains incurable. This age-related neurodegenerative disease is characterized by an early decline in episodic and spatial memory associated with progressive disruption of the hippocampal functioning. Recent clinical evidence suggests that impairment of the spatial pattern separation (SPS) function, which enables the encoding and storage of episodic spatial information, may be an indicator of the early stages of AD.
Objective:
The aim of our study was to characterize SPS performance at a prodromal stage in 5xFAD transgenic mouse model of AD.
Methods:
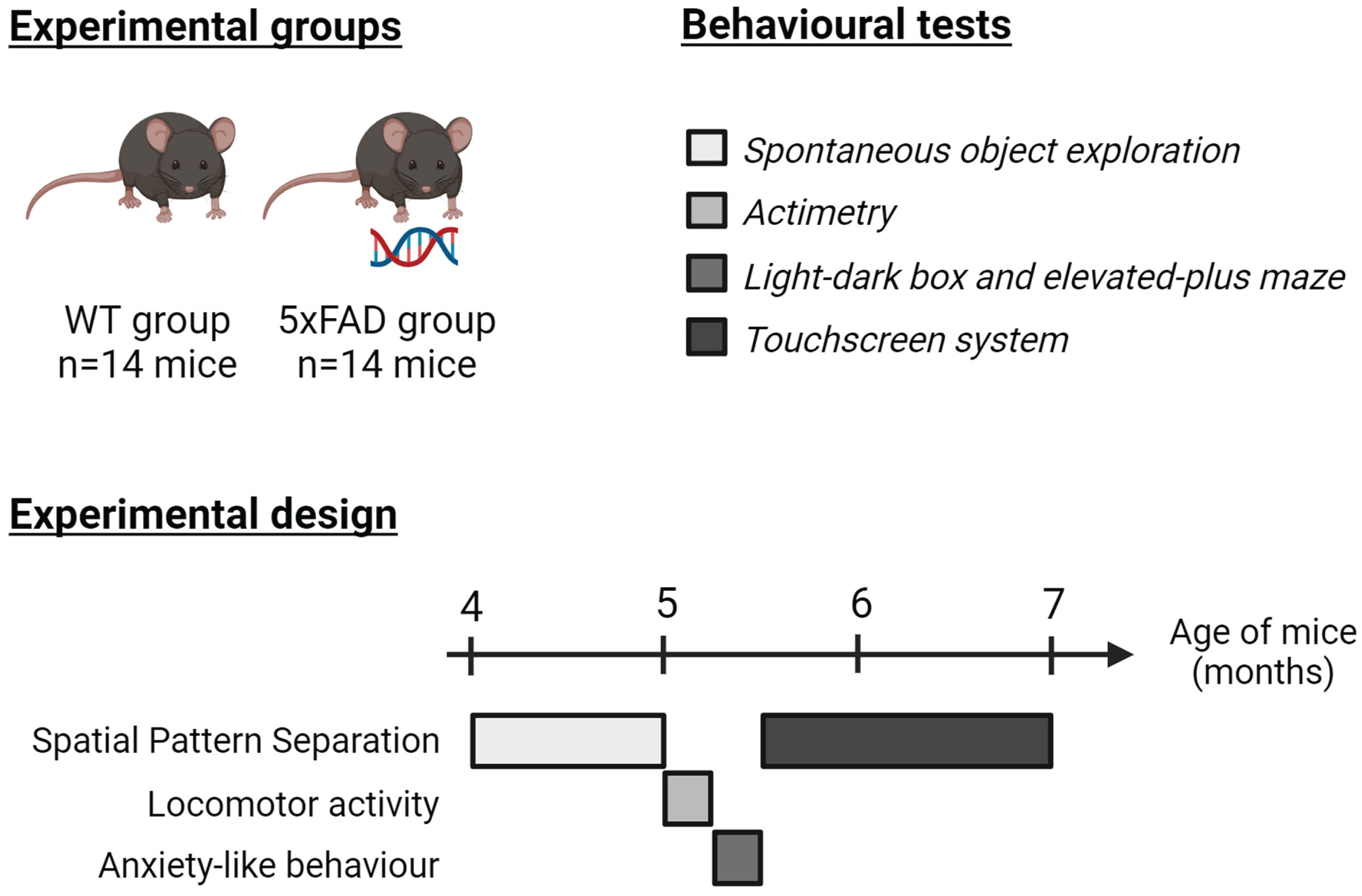
Behavioral performance of male wild-type (WT) and 5xFAD mice (n = 14 per group) was assessed from the age of 4 months in two validated paradigms of SPS function either based on spontaneous exploration of objects or on the use of a touchscreen system.
Results:
Compared with age-matched WT littermates, a mild deficit in SPS function was observed in the object recognition task in 5xFAD mice, whereas both groups showed similar performance in the touchscreen-based task. These results were observed in the absence of changes in locomotor activity or anxiety-like behavior that could have interfered with the tasks assessing SPS function.
Conclusions:
Our results indicate an early vulnerability of the SPS function in 5xFAD mice in the paradigm based on spontaneous exploration of objects. Our work opens up the possibility of examining the early neurobiological processes involved in the decline of episodic memory and may help to propose new therapeutic strategies in the context of AD.
INTRODUCTION
Alzheimer’s disease (AD) represents the most common cause of age-related dementia and is characterized by an early decline in episodic memory [1]. Associated hippocampal atrophy is negatively correlated with episodic information encoding abilities through a hippocampal-dependent neural process called spatial pattern separation (SPS) [2, 3]. SPS function allows distinct episodic memories to be created by encoding similar but distinct spatial information without any interference [4]. Indeed, past experiences can share overlapping content, hindering the retrieval of a specific event in the context of disruptive memories [5, 6]. The involvement of the dentate gyrus as a pattern separator has been demonstrated both in humans [7, 8] and animals [5, 10]. Structural and functional impairment in the dentate gyrus/CA3 region negatively correlated with SPS function over ageing [11]. In particular, altered pathway and dendritic integrity have been observed in these brain regions in elderly subjects [11], which translates into an inability to recall an episodic memory accurately [12].
Increasing the level of spatial interference between two identical items results in poor discrimination performance. This is notably true in the context of aging. An initial study showed that elderly subjects carrying the ɛ4 allele of apolipoprotein E (subjects at risk of developing AD) showed significantly impaired discrimination performance with increasing interference compared with elderly subjects not carrying the ɛ4 allele [13]. Using computer-based paradigm, such deficit in SPS function has also been identified in mild cognitive impairment and AD patients [2]. It is interesting to note that the more advanced the stage of AD, the greater the discrimination deficit and the degree of atrophy of the hippocampus and entorhinal cortex [2]. According to the results of these studies, identifying deficits in SPS function in subjects at risk could serve as an early indicator of progression to AD. This would offer the possibility to treat cognitive disorders earlier, i.e., before the stage of AD is too advanced and the deficits can no longer be countered. The development of paradigms dedicated to the assessment of SPS function in rodents [14, 15] could therefore facilitate the identification of early neurobiological dysfunctions linked to AD and thus improve the translational efficacy of preclinical trials testing pro-cognitive drugs.
The aim of our study was to characterize for the first time the SPS performance in a mouse model of AD in two behavioral tests that have already been validated in the literature as involving the function of the SPS. The first paradigm was based on spontaneous exploration of objects [15] while the second was based on the use of a touchscreen system [14]. The AD model chosen was the 5xFAD mouse, carrying five human APP and PSEN1 mutated transgenes, and characterized by an early brain accumulation of amyloid-β peptide from 2 months of age followed by gliosis and neuronal degeneration beyond 4 months [16]. These processes lead to the accelerated formation of amyloid deposits in the hippocampus as early as 6 months of age [16, 17], coinciding with the onset of episodic-like memory deficits [18]. We consequently hypothesized that early deficits in SPS function may be identified at a prodromal stage of the disease in 5xFAD mice, i.e., from 4 months of age. Such preclinical studies are essential to gain a better understanding of the mechanisms underlying the early SPS impairment associated with AD and may help to identify new therapeutic strategies.
MATERIALS AND METHODS
Animals
All experiments were performed on adult male transgenic 5xFAD mice [16] (n = 14, “5xFAD” group) and age-matched C57BL/6 littermates (n = 14; Wild-type “WT” group) (4-month-old at the start of testing; CURB, Caen, France). The 5xFAD mice were initially created on C57BL6/SJL background and were then derived with a C57BL/6 genetic background as described before [19]. Mice were housed in transparent polycarbonate cages (42 × 29 × 15 cm, 2–5 mice per cage; containing nesting material and a cardboard tube) and maintained under a 12:12 h reverse light-dark cycle (light on from 7 p.m. to 7 a.m.), at constant temperature (22°C) and hygrometry (55%). Mice had access to water and food ad libitum except when specifically mentioned. All experiments were conducted in compliance with national and European regulations concerning animal experimentation (EU directive N°2010/63; project authorization number APAFIS#29543).
Experimental design
All experiments were conducted between 8:30 a.m. and 12:30 p.m. Mice were acclimatized to the testing room for 30 min before experiment starts. All mice were exposed to the two tasks assessing SPS function. In addition, we evaluated locomotor activity and anxiety-like behavior since any disturbances of these factors may potentially affect mice’s performance in paradigms dedicated to the assessment of SPS function (Fig. 1).
Actimetry
Spontaneous locomotor activity was assessed in Imetronic® apparatus composed of 8 individual activity cages (11.3 × 26 cm) crossed by horizontal and vertical infrared sensors. During 60 min, the total number of crossings of sensors was counted and used as horizontal and vertical locomotor activity index [20].
Light-Dark box test
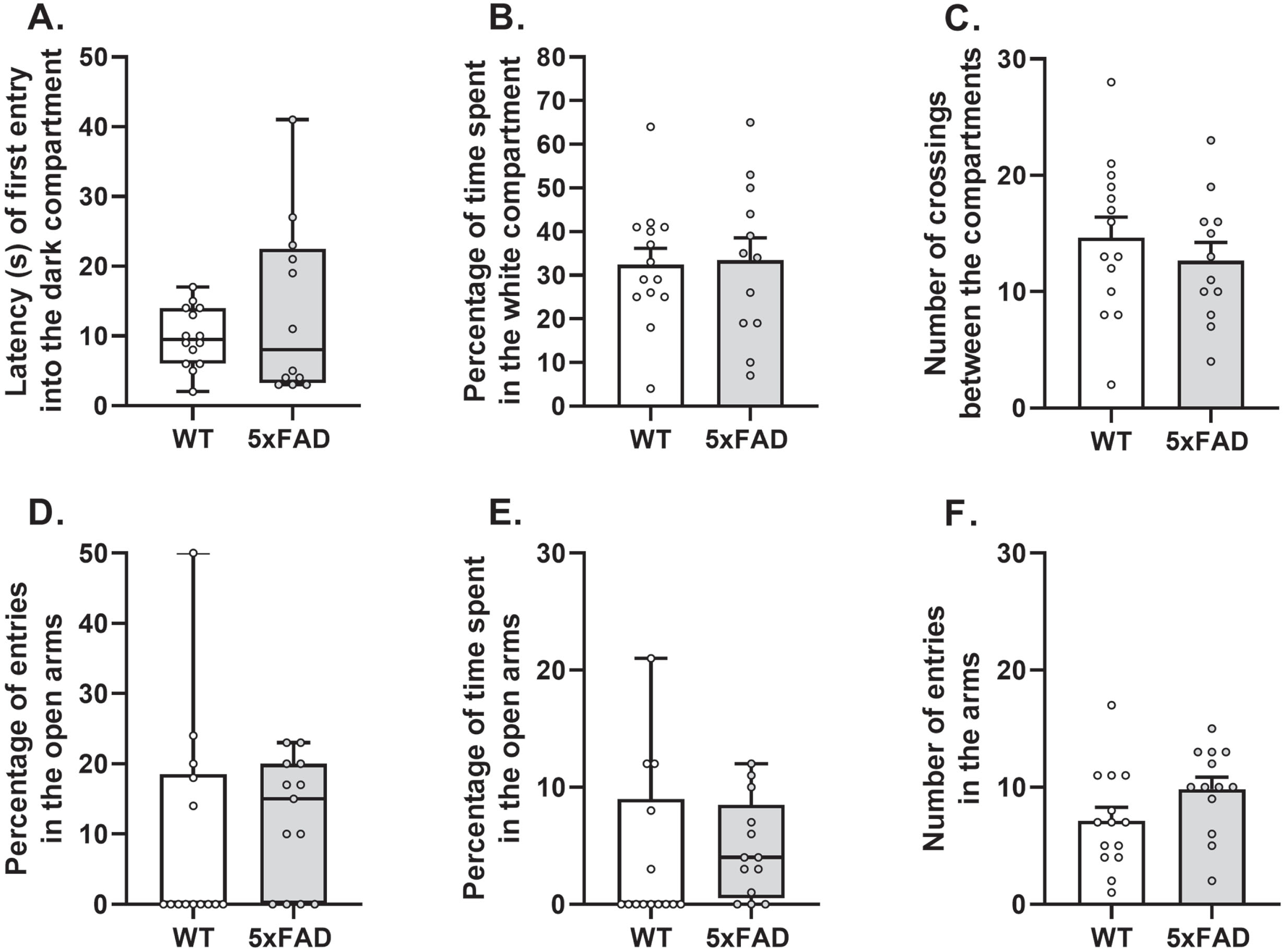
The apparatus (Bioseb®; 42.5 × 28 × 27 cm) is constituted of an illuminated white compartment (26 × 26 cm; 900 lux) and a dimly lit black compartment (17.5 × 6 cm; 100 lux) separated by an open trap door. This task is based on innate preference of mice for dark and confined spaces and was performed as described before [21]. Each mouse was initially placed in the center of the white compartment and left free to explore the apparatus for 5 min. The latency of first entry in the dark compartment and the total time spent in the white compartment were used to evaluate the anxiety-like behavior while the number of crossings between both compartments was used to evaluate locomotor and/or exploratory activity. One 5xFAD mouse was excluded from the analysis due to insufficient exploration (no entry in the dark compartment within 120 s).
Elevated-plus maze
The apparatus is constituted of two open arms (66 × 5 cm; 40 lux) crossing two closed arms (66 × 5 × 15 cm; 10 lux) connected by a central platform (5 × 5 cm) and raised at 40 cm above the floor. Each mouse was placed on the platform, facing an open arm. During 5 min, the time spent and the number of entries in each arm (open or closed ones) were collected, as described before [21]. Considering the innate preference of mice for dark and confined spaces, these parameters were used as an index of anxiety. The total number of entries in all arms was used to evaluate locomotor and/or exploratory activity.
Spatial pattern separation paradigm based on spontaneous object exploration
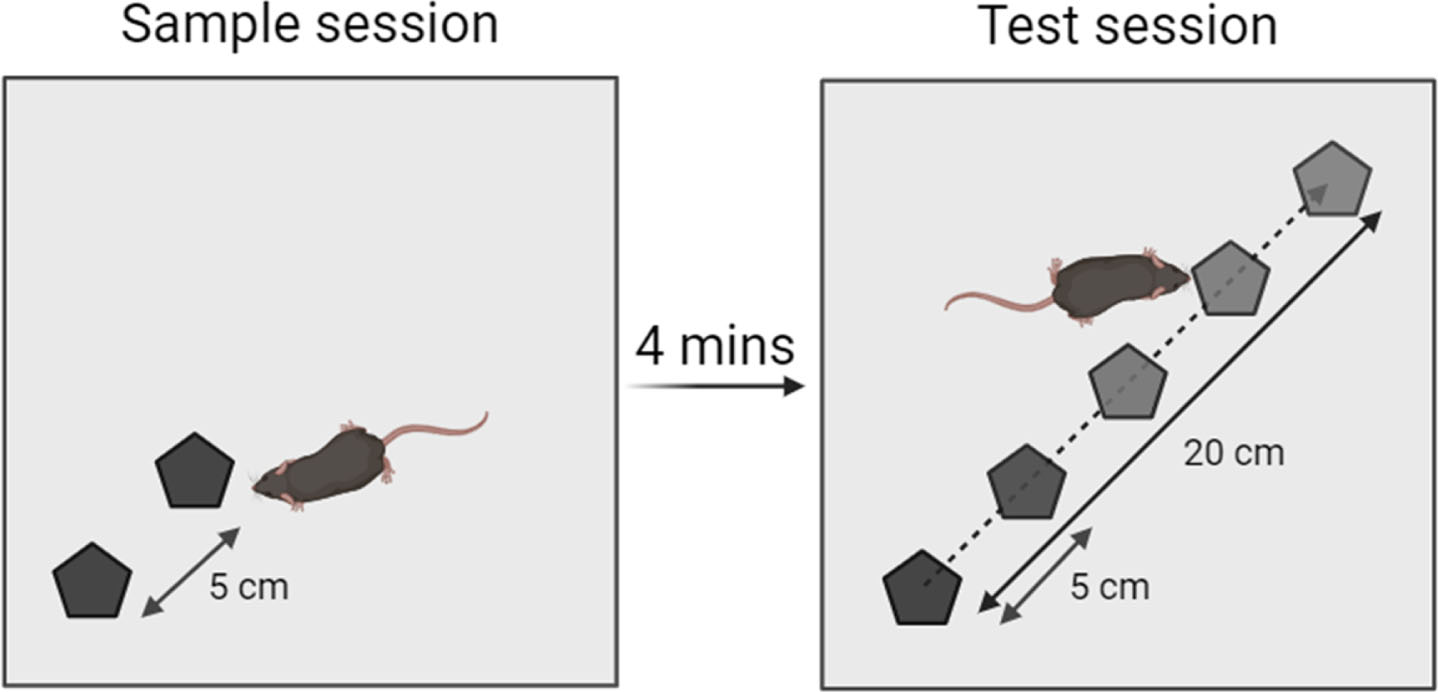
This test is based on the natural curiosity of rodents towards objects that have been displaced in a familiar environment [15]. It allows to assess SPS ability by identifying the minimum distance of object displacement detected by the mouse, resulting in greater exploration of the displaced object than of the stationary object. The apparatus consisted of a grey wooden open-field (32 × 32 × 32 cm; 30 lux in the center) containing 2 identical objects arranged diagonally (Fig. 2). The position of the objects (left or right diagonal) was randomized. Between each session, the objects and apparatus were carefully cleaned with 70° ethanol to remove olfactory cues.
Each mouse was first familiarized to the open-field containing 2 identical objects (placed diagonally 6 cm from the corners) for 10 min (2 sessions/day during 2 consecutive days).
The task consists in a sample session followed 4 min after (inter-session interval, ISI) by the test session. During the sample session, each mouse was placed in the open-field containing 2 identical objects, different from those used previously and placed 5 cm apart (Fig. 2). During the test session, the object farthest from the corner was diagonally displaced from either 5, 10, 15, or 20 cm. For each session, the mouse was allowed to freely explore both objects during 12 min. The time spent to explore each object, i.e., defined by active sniffing of the object at a distance below 2 cm, was manually measured. The exploration time of the displaced object was then expressed as percentage of exploration (time of exploration of the displaced object/time of exploration of both objects × 100) and compared to the theoretical value of 50% (chance level). One 5xFAD mouse was excluded from the analysis due to insufficient exploration behavior (objects exploration for less than 3 s in one 12-min session).
Each mouse was exposed to the task for four consecutive days. For each mouse, a single, randomly chosen object displacement distance (5, 10, 15, or 20 cm) and a new pair of objects (plastic tubes, Lego® towers, ceramic flasks or dolphins) were used per test day.
Spatial pattern separation paradigm based on touchscreen system
Apparatus. An automated touchscreen operating chamber system was used (Bussey Mouse Touchscreen Chamber; Campden instruments®) to assess the ability of mice to distinguish between two similar visual objects placed near or far from each other [14]. Briefly, the apparatus consisted of a trapezoid sound- and light-attenuating chamber (40 × 34 × 42 cm), with touchscreen wall (25 × 31 × 5 cm). A black Plexiglas mask (28 × 24.3 cm) allowed to project the two identical visual stimuli in either of the 6 horizontally aligned rectangles (3 × 2 cm each). On the opposite side of the chamber, a drop dispenser (creamy strawberry yogurt, Yoplait®, France) was used to reward the mouse for every nose-poke made correctly on the screen. All data were automatically collected with ABET II Touch v.3.3 software (Lafayette Instruments®).
Food restriction. A mild food restriction protocol was initiated one week prior to training. Mice were then maintained at 85–95% of their original body weight [22, 23].
Pre-training. Mice were first familiarized to the chamber and to the liquid reward for a 30-min daily session (3 days; no visual stimulus on touchscreen). Then, the mice were progressively pre-trained to initiate a trial and point their noses towards the screen to obtain a reward. During the initial touch stage, a visual stimulus was randomly displayed for 30 s and a food reward was delivered independently of the mouse’s interaction with the screen. The amount of reward was 3 times greater if the mouse interacted with the screen. During the must-touch stage, the reward was only delivered if the mouse interacted with the screen during the stimulus presentation time. During the must-initiate stage, the mouse had to initiate a trial by interacting with the reward dispenser when it was switched on. The criterion for passing from one of these stage to the next was the completion of 30 trials in 60 min. This was finally followed by the punish incorrect stage during which the mouse had to learn that only interaction with the stimulus on the screen triggered reward dispensing. An incorrect response (i.e., touching with the nose outside the visual stimulus displayed) was followed by switching off the chamber for 5 s. The visual stimulus was displayed again on the same location until a correct response was given. The completion criterion to move to the location discrimination task was set at 77% correct responses (not including correction trials) within 30 min over 2 consecutive days. One mouse of each group failed to reach this criterion and was excluded from the analysis.
Training phase of the location discrimination paradigm. Mice were trained to distinguish between two identical visual stimuli displayed on screen and separated by an intermediate separation (two unlit windows left between, i.e., illumination of the 2nd and 5th windows) corresponding to a distance of 9 cm. One of the stimuli was randomly assigned to a correct response and rewarded (Fig. 3). The completion criterion was set at 7 out of 8 consecutive correct responses. Once achieved, the rewarded stimulus was reversed. The daily session ended either when 60 trials were completed or after 60 min had elapsed, whichever occurred first. The location of the first rewarded stimulus was randomized between mice and changed at each daily session. The training phase ended when the completion criterion was reached in 3 out of 4 consecutive sessions.
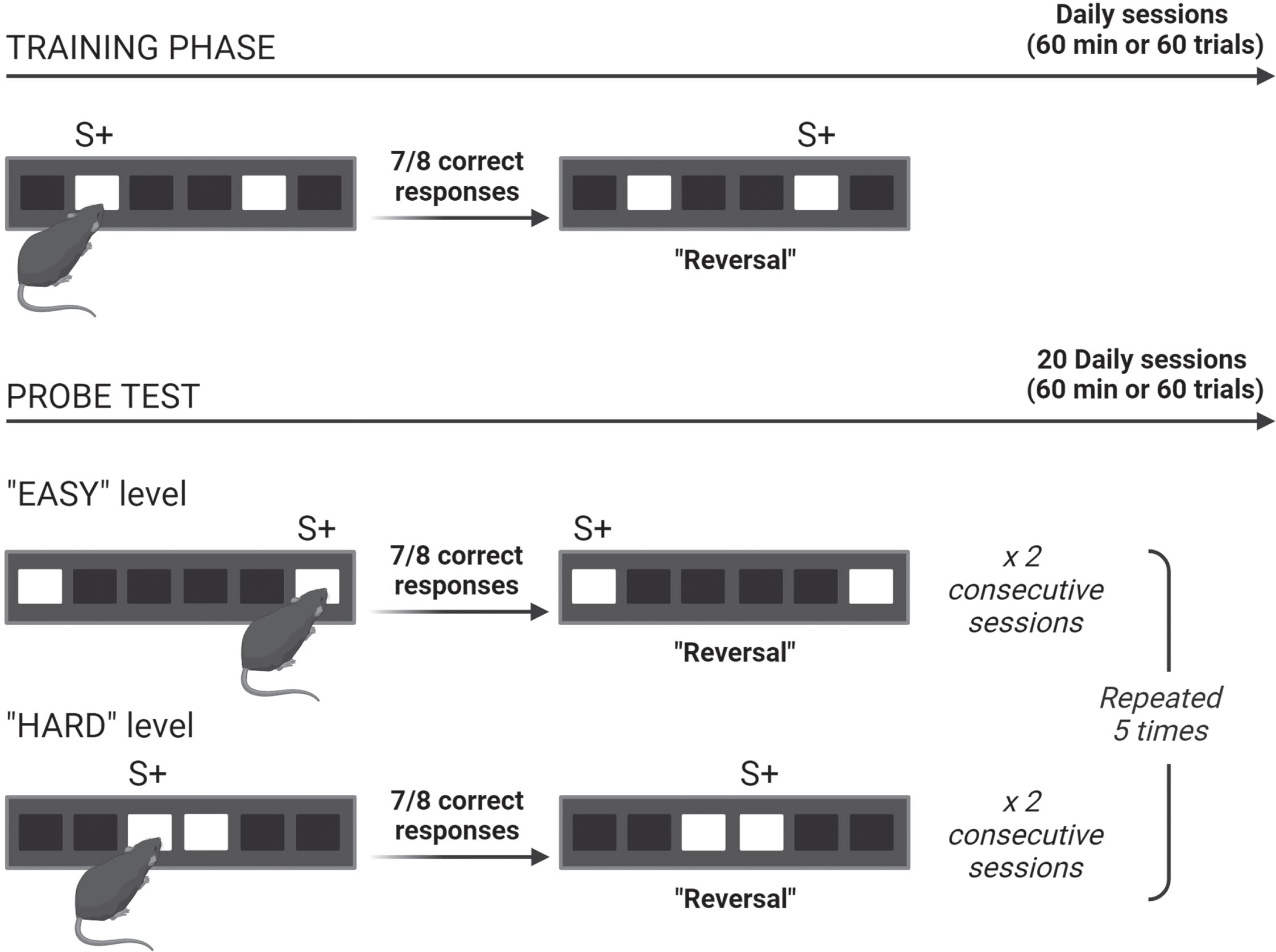
Probe test of the location discrimination paradigm. At each daily session, two identical visual stimuli were presented simultaneously at a large separation of 17 cm (“EASY” session) or at a small separation of 1 cm (“HARD” session). The mouse’s ability to distinguish between the stimuli was assessed over 20 sessions comprising 10 repetitions of 2 consecutive “easy” and then “hard” sessions. The starting order of the session, i.e., easy or hard one, was randomly selected. As before, one of the two stimuli was randomly assigned to a correct response and was rewarded. A similar completion criterion (7 out of 8 consecutive correct responses) was applied before reversing the rewarded stimulus. The session ended when 60 min had elapsed or when the mouse had completed 60 trials. The average performance of the 10 sessions of each difficulty level was measured. The mean percentage of correct responses and the mean number of trials required to reach the criterion over the 10 sessions of each difficulty level were collected and used as indices of the SPS performance. The mean number of reversals performed during the same session (when the stimulus rewarded was reversed) was used as an index of cognitive flexibility. Finally, the latencies to interact with the touchscreen and to retrieve the reward were used to assess the animal’s motivation.
Statistical analysis
Statistical analysis and graphical representations were performed using RStudio® and GraphPad Prism®, respectively. The homogeneity of variances (Levene’s test) and normality of the data (Shapiro-Wilk test) were first assessed. When these two assumptions were met, data were analyzed using parametric statistical tests (ANOVA) and represented as mean±standard error of the mean (SEM). Otherwise, non-parametric equivalent statistical tests were used (Mann-Whitney tests and two-way non-parametric statistics tests for longitudinal data with R package “nparLD” and followed by Wilcoxon rank sum tests for pairwise comparisons) and data were represented as median±quartiles. Comparisons with the chance level were analyzed using univariate t-tests or Wilcoxon signed rank tests. One 5xFAD mouse died during the study and was excluded from all analysis. A significant difference was considered when the p value was lower than 0.05.
RESULTS
5xFAD mice showed locomotor hyperactivity without anxiety-like behavior alteration
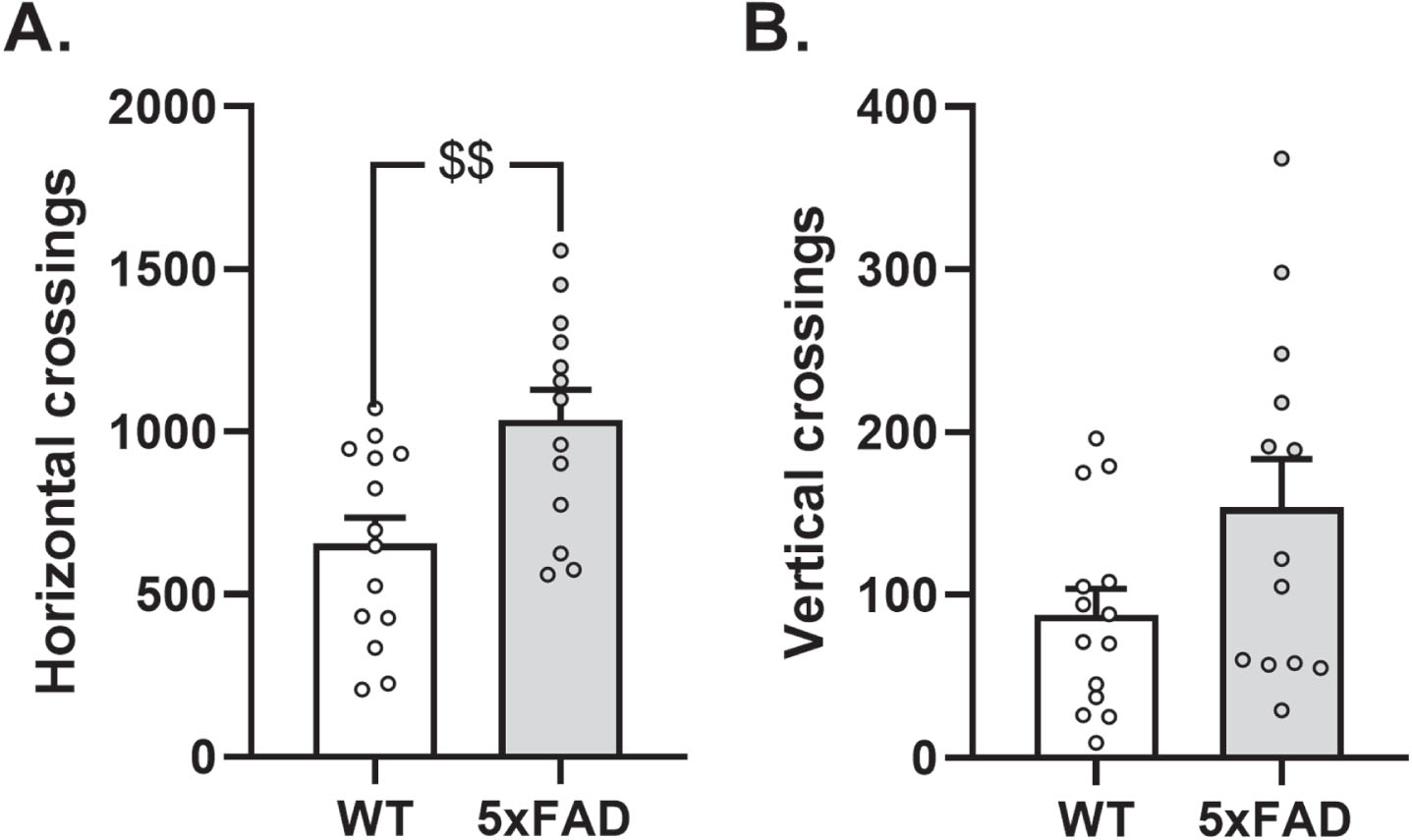
Analysis of locomotor activity during the 60-min actimetry session revealed a significantly higher cumulative number of horizontal crossings through the infrared sensors in the 5xFAD group than in the WT group (ANOVA: F (1 ;25) = 9.79, group effect: p < 0.01 and F (1 ;25) = 3.95, p > 0.05 for horizontal and vertical crossings respectively; n = 14 WT mice and n = 13 5xFAD mice; Fig. 4A, B). These results indicate that 5xFAD mice exhibited greater horizontal locomotor activity.
In the light-dark box, the latency of first entry into the dark compartment and the percentage of time spent in the white compartment did not differ between groups (Mann-Whitney test: U = 82.5, p > 0.05 and ANOVA: F (1 ;24) = 0.03, p > 0.05; respectively; n = 14 WT mice and n = 12 5xFAD mice; Fig. 5A, B). The total number of crossings between the two compartments did not differ either (ANOVA: F (1 ;24) = 0.68, p > 0.05; Fig. 5C). In the elevated plus maze, the percentage of entries and the time spent in the open arms did not differ between groups (Mann-Whitney test: U = 69.0, p > 0.05 and U = 66.5, p > 0.05, respectively; n = 14 WT mice and n = 13 5xFAD mice; Fig. 5D, E). The cumulative number of entries in the arms did not differ either (ANOVA: F (1 ;25) = 3.10, p > 0.05; Fig. 5F). Taking together, these results suggest comparable level of anxiety-like behavior in two groups.
5xFAD mice showed impaired performance in SPS paradigm based on spontaneous object exploration
During the sample session, mice showed no preference for either of the two objects explored (Univariate t-tests, p > 0.05 at all distances tested, whatever the group considered). No differences between groups were detected (2-way non-parametric statistics for longitudinal data: ATS (ANOVA-Type Statistic) = 1.55, no group effect, p > 0.05; no distance effect: ATS = 1.69, p > 0.05; no interaction: ATS = 0.53, p > 0.05; Fig. 6A).
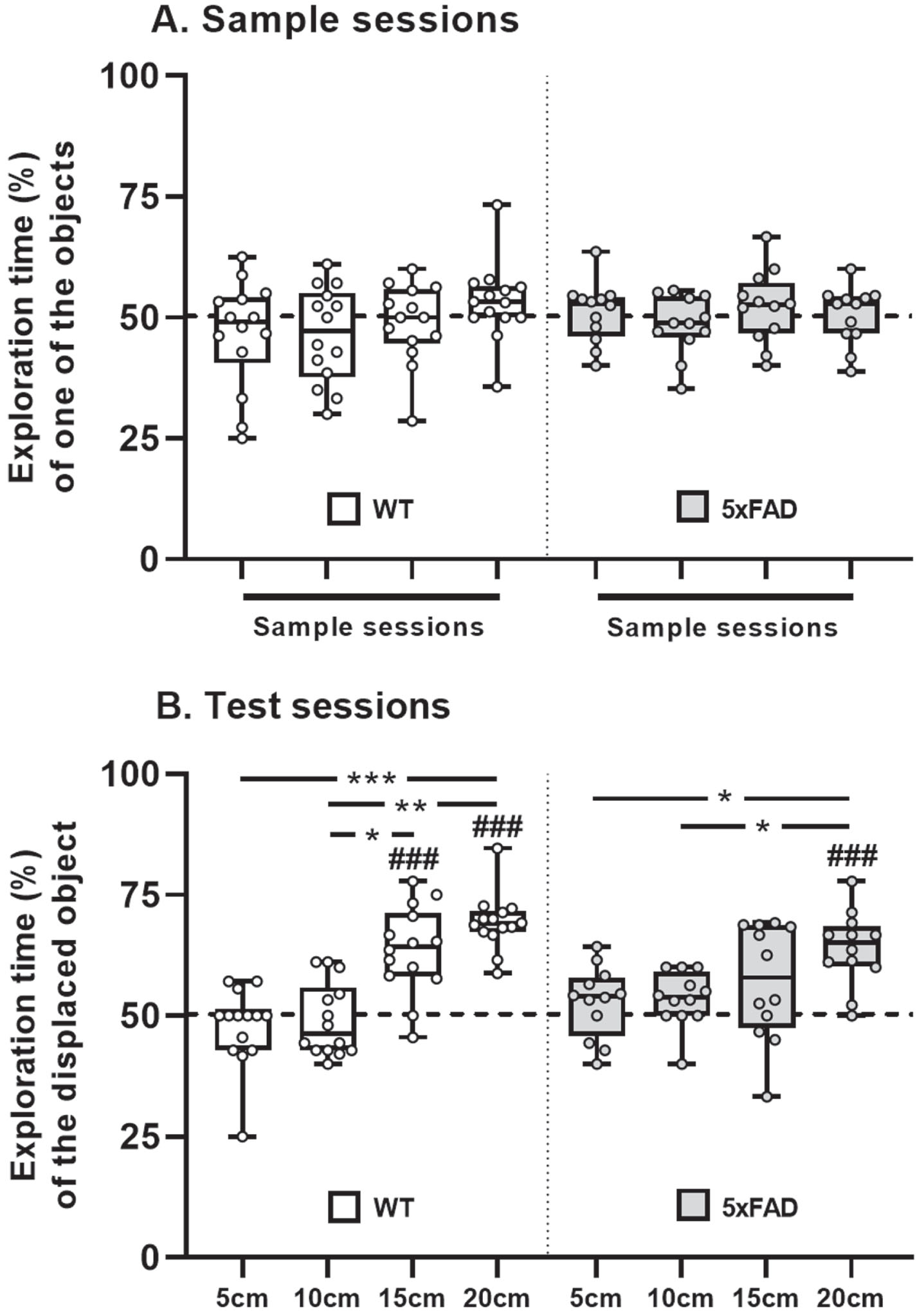
During the test session, WT mice significantly detected object displacement at the distance of 15 and 20 cm (Univariate t-test, t = 5.58, p < 0.001 and Wilcoxon signed rank test, p < 0.001, respectively). In contrast, the displaced object was only detected at the distance of 20 cm in 5xFAD mice (Univariate t-test, t = 6.42, p < 0.001). Two-way non-parametric statistics for longitudinal data did not detect a significant difference between groups (ATS = 0.15, p > 0.05) but revealed a significant effect of the distance between the objects (ATS = 27.96, p < 0.001) and a significant interaction (ATS = 4.17, p < 0.05). In both groups, a significantly higher percentage of time spent to explore the displaced object was observed at the distance of 20 cm in comparison with 10 and 5 cm (Pairwise comparisons using Wilcoxon signed rank exact test: for WT group, p < 0.01 and p < 0.001; for 5xFAD group, p < 0.05 and p < 0.05, respectively). By contrast, the percentage of time spent to explore the displaced object was found to be significantly higher at the distance of 15 cm in comparison with 10 cm (p < 0.05), in the WT group only (Fig. 6B).
5xFAD mice showed intact performance in SPS paradigm based on touchscreen system
The number of sessions required to complete each stage of the pre-training and the training phase did not differ between groups (Mann-Whitney test: U = 72, p > 0.05 for the Initial touch stage; ANOVA: F (1,23) = 1.15, p > 0.05 for the Must-touch stage; Mann-Whitney test: U = 60, p > 0.05 for the Must-initiate stage and ANOVA: F (1,23) = 0.69, p > 0.05 for the Punish incorrect stage; Mann-Whitney test: U = 78, p > 0.05 for the Training phase; data not shown), indicating similar basal performance between WT and 5xFAD mice.
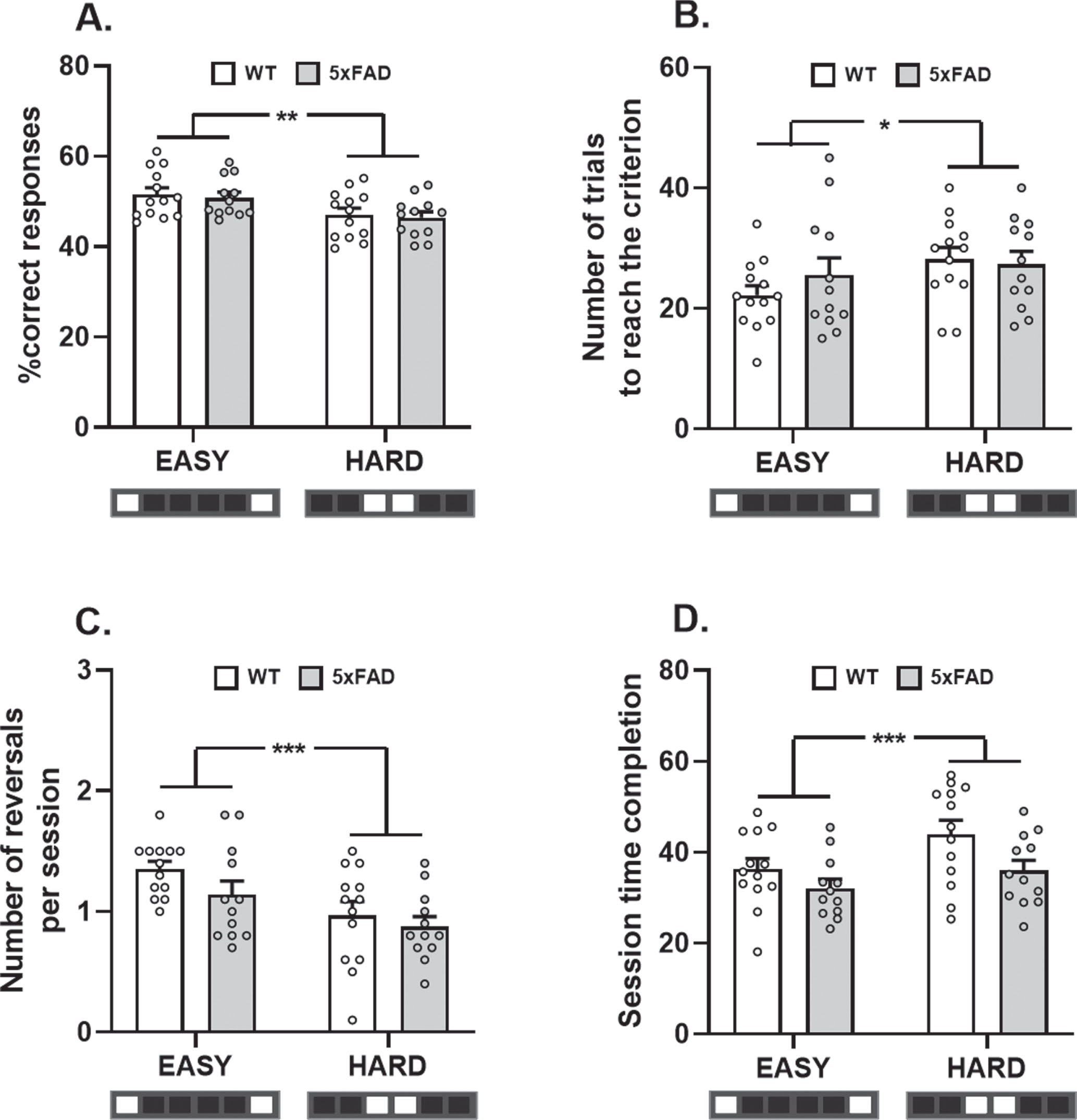
During the probe sessions, SPS performance was significantly reduced with increasing level of difficulty. Indeed, the percentage of correct responses was significantly reduced and the number of trials to reach the criterion was significantly increased in the “hard” separation as compared to the “easy” one (ANOVA with repeated measurements: effect of difficulty level: F (1,23) = 10.46, p < 0.01 and F (1,23) = 5.06, p < 0.05, respectively; Fig. 7A, B). However, no difference between groups was detected (no group effect: F (1,23) = 0.26, p > 0.05 and F (1,23) = 0.25, p > 0.05; and no interaction: F (1,23) = 0.01, p > 0.05 and F (1,23) = 1.43, p > 0.05, respectively). The mean number of reversals reached per session was also found to be decreased with increasing difficulty level (effect of difficulty level: F (1,23) = 14.84, p < 0.001; Fig. 7C). The lack of statistical difference between groups suggests similar cognitive flexibility performance in WT and 5xFAD mice (no group effect: F (1,23) = 2.11, p > 0.05 and no interaction: F (1,23) = 0.49, p > 0.05).
Finally, the time spent completing session did not differ between groups (no group effect: F (1,23) = 3.61, p > 0.05), but was significantly increased when the level of difficulty was harder (effect of difficulty level: F (1,23) = 20.26, p < 0.001 and no interaction: F (1,23) = 2.15, p > 0.05; Fig. 7D). Latencies to interact with the touchscreen and to retrieve the reward did not change between groups (no group effects: ATS = 0.01, p > 0.05 and ATS = 0.01, p > 0.05, respectively; data not shown), indicating a similar level of motivation between groups. It should be noted that the latency to interact with the touchscreen was significantly increased when the difficulty level was higher (effect of difficulty level: ATS = 33.20, p < 0.001 and no interaction: ATS = 1.67, p > 0.05).
DISCUSSION
We aimed here to assess SPS ability in a preclinical model of AD using two complementary hippocampal-dependent tasks, based either on spontaneous objects exploration [15] or touchscreen device [14, 24]. To our knowledge, this is the first study to examine the function of SPS in two different tasks in the same mouse model of AD. While the spontaneous object exploration task allowed to highlight a mild deficit of SPS function in 5xFAD mice, the touchscreen-based assay did not.
In order to exclude any interference due to differences in locomotor and/or anxiety-like behavior on the assessment of SPS function, actimetry and anxiety-like behavior tasks were performed at 5 months of age, i.e., between the two paradigms evaluating the SPS function. In the actimetry test, 5xFAD mice showed motor hyperactivity compared with WT mice, as previously reported at 6 months of age [25]. According to the literature, which reports anxiety-like behavior changes in 5xFAD mice from 6 months of age [17, 26] and later [27–30], no change was detected in our study. Additional analyses were conducted on the data collected during the familiarization session to evaluate the animal’s baseline behavior before the object-based paradigm. The analysis of the overall distance travelled in the open field, the percentage of distance travelled, and the percentage of time spent in the central arena zone indicated no significant differences in locomotor activity and anxiety-like behavior between the groups (data not provided). Overall, our results indicate that 5xFAD mice did not show a decrease in locomotor activity or changes in anxiety-like behavior that could have interfered with the tasks assessing SPS function.
The SPS paradigm based on object exploration is of ecological interest as it is based on the spontaneous exploratory behavior. This free-training test assesses the rodent’s ability to discriminate an object whose location has changed between two sessions. Accordingly, WT mice spent more time exploring the displaced object when the distance between objects was greater than 10 cm apart. A similar displacement detection limit has been described in the literature [15]. In contrast, 5xFAD mice failed to discriminate the two objects as different when the separation distance was less than 20 cm. Despite the lack of significant differences between groups, the results indicate that 5xFAD mice display a mild deficit in SPS function. Although this test has already been validated in adult rodents [31] and under age-related forgetting conditions by showing an SPS deficit as early as 11 months of age [15], no study to date has assessed SPS function in a mouse model of AD using this paradigm. Interestingly, amyloid deposition starts to develop as early as 2 months of age in 5xFAD mice [16, 32] and is associated with neuro inflammatory processes (activated astrocytes and microglia) [16, 33]. At 6 months of age, a severe reduction in cerebral metabolism was reported in both hippocampus and cerebral cortex [34]. Finally, synaptic impairments were observed [35, 36] at the time when cognitive deficits are most frequently reported [16, 37]. Among these deficits, episodic-like memory has been reported to be impaired as early as 6 months of age in 5xFAD mice [18]. Our results thus corroborate those described in the human literature [2], suggesting an early deficit in SPS function before the onset of episodic memory deficits in 5xFAD mice.
The second SPS paradigm used is based on visual discrimination of stimuli projected on a touchscreen in an operant chamber. This test also assesses the rodent’s ability to discriminate between two visual stimuli whose separation distance varies. In adult rodents, the smaller the distance between visual stimuli, the more difficult it is for the animal to distinguish between them [14, 24]. Such a device offers several advantages, such as higher standardization of the protocol than the one performed in an open-field, minimal intervention by the experimenter and the transposition of cognitive tasks from humans to animals providing a greater translational value [38, 39]. However, as it is based on the use of positive reinforcement (liquid reward), the results are sensitive to differences in motivation level between experimental groups. In this operant chamber paradigm, all the parameters collected indicated a significant reduction in performance in WT mice when the distance separating the visual stimuli was small (“hard” condition). Indeed, the number of trials required to reach the criterion was increased, while the percentage of correct responses and the number of reversals reached per session were significantly reduced. In addition, the time to complete the session and the latency to interact with the screen were longer with reducing separation distance. These results in WT mice are in line with expectations from the literature. Surprisingly, no difference was detected between the 5xFAD group and the WT one. Moreover, both groups showed a similar level of motivation since the latency to obtain the reward did not change. These results therefore indicate that 5xFAD mice are capable of performing the touchscreen-based task successfully, which does not support previous findings reported in transgenic animal models of AD. Indeed, both APP transgenic mouse model [40, 41] and 5xFAD mice [42] showed lowered performance during probe sessions. AD mice needed more trials to reach the criterion when the distance separating the stimuli was small compared with to the age-matched WT littermates [41, 42]. This deficit was however not observed in our study.
Several possible explanations may account for the difference in results, including variations in the protocol used during the probe test assessment. The study by Li and colleagues [42] used fewer daily sessions than our own study (3 sessions compared to 20 sessions). It is possible that a deficit only appeared at the early stage of the probe test. To investigate this hypothesis, we conducted a complementary analysis on the first four days of the probe test, consisting of two “easy” sessions followed by two “hard” sessions. However, no difference in the percentage of correct responses and the number of trials to reach the criterion was detected between groups (data not shown), which suggests the lack of SPS dysfunction in 5xFAD mice during the initial sessions of the probe test.
It should also be noted that there is a difference in the age at which the performance of 5xFAD mice was evaluated: around 4 months in the literature [42] and 6 months in our study. Interestingly, hippocampal neurogenesis is strongly implicated in SPS function [6, 43–45]. According to the literature, 5xFAD mice show an early decline in hippocampal neurogenesis from 2 months of age. From 6 months of age, a physiological decline in neurogenesis was also reported in age-matched WT mice, resulting in levels comparable to those of 5xFAD mice [46]. Thus, the decline in neurogenesis in 5xFAD mice, specifically between 2 and 6 months, may contribute to the early SPS dysfunction identified by Li et al. through the touchscreen task [42]. This could also explain the lack of impairment noticed in the same task in our 6-month-old 5xFAD mice. Although it has to be confirmed, the early deficit in SPS function based on object recognition detected in 4-month-old 5xFADs also coincides with this period of impaired neurogenesis. In this context, it appears that a crossover design, which lacks to our study, would have been of high interest to assess the behavioral performance in the two paradigms evaluating SPS function in distinct cohorts of 5xFAD mice at the same age. Alternatively, different cognitive processes are involved in these two paradigms and we might expect to see different results. For example, the object recognition task requires the memorization of their spatial configuration for several min (4 min-ISI). Moreover, the nature of the object changes across sessions. In contrast, recall of the rewarded stimulus between trials of the touchscreen-based task is shorter (less than 1 min) and the nature of the visual stimuli is always the same. Besides, the touchscreen task requires a higher number of daily training sessions and relies on food restriction which must be maintained for several weeks to motivate the performance of the mice. The touchscreen paradigm may therefore involve working memory, attentional processes, cognitive flexibility, and operant conditioning. A crossover design would enable to determine if the early SPS function deficit detected is dependent on the behavioral paradigm used. Finally, this study was conducted solely in male mice to allow for comparison with the study performed by Cès and colleagues in adult male C57BL6 mice [15]. An inclusive study using female 5xFAD and WT mice may also be an interesting avenue to follow.
In conclusion, our results showed that a hippocampal-dependent task based on object exploration revealed SPS deficits in 5xFAD mice as early as 4 months of age. This paradigm could therefore help to gain a better understanding of the neurobiological processes involved in early SPS dysfunction and to test the efficacy of new drug candidates in reducing or delaying the onset of early cognitive deficits associated with AD.
Footnotes
ACKNOWLEDGMENTS
We gratefully thanks Gerald Nee, Stacy Largilliere, and Melody Mahier for technical assistance. Illustrations have been created with BioRender.com.
FUNDING
This work was supported by the Université de Caen Normandie, INSERM. Lucas Gephine is supported by funding from the regional council (Region Normandie, COMUE, FRANCE).
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
All the data supporting the behavioral findings of this study are available on request from the corresponding author.