
Abstract
Background:
Alzheimer’s disease (AD) is a progressive neurodegeneration disease. Physical activity is one of the most promising modifiable lifestyles that can be effective in slowing down the progression of AD at an early stage.
Objective:
Explore the molecular processes impaired in AD that were conversely preserved and enhanced by physical activity.
Methods:
Integrated transcriptomic analyses were performed in datasets that contain AD patients and elders with different degrees of physical activity. The changes of the hub genes were validated through analyzing another two datasets. The expression of the hub genes was further detected in the hippocampus and cortexes of APP/PS1 transgenic mice with or without physical activity by Quantitative polymerase chain reaction (qPCR).
Results:
Cross-comparison highlighted 195 DEGs displaying opposed regulation patterns between AD and high physical activity (HPA). The common DEGs were predominantly involved in synaptic vesicle recycling and synaptic transmission, largely downregulated in AD patients but upregulated in the elders with HPA. Two key modules and four hub genes that were related to synaptic vesicle turnover were obtained from the PPI network. The expression of these hub genes (SYT1, SYT4, SH3GL2, and AP2M1) was significantly decreased in AD transgenic mice and was reversed by HPA training.
Conclusions:
HPA may reverse AD pathology by upregulating a range of synaptic vesicle transport related proteins which might improve the efficiency of synaptic vesicle turnover and facilitate inter-neuronal information transfer. The study provides novel insights into the mechanisms underlining the protective effects of HPA on AD.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is one of the most common neurodegenerative diseases among the aged, which accounts for more than two-thirds of the total number of dementia patients. As the global elderly population increases, AD poses a serious threat to human health. According to the latest statistics released by Alzheimer’s Disease International, the global prevalence of dementia is expected to increase to 78 million by 2030 [1]. The total estimated worldwide cost of dementia now averages more than $800 billion annually, which represents 1.09% of global GDP [1]. Therefore, both research on the mechanisms of the development of AD and effective treatment are urgently needed.
The main pathological characteristics of AD are represented by brain atrophy, synaptic loss, senile plaques, and neurofibrillary tangles [2, 3]. Reduction in synaptic plasticity caused by deposition of amyloid-β (Aβ) and hyperphosphorylation of tau proteins has been shown to be the underlying cause of the progressive decline in learning and memory in AD [4]. The presence of tau oligomers and soluble Aβ in the brains of AD patients has been demonstrated to allow glial cells to potentially over-phagocytose synapses in the brain, causing early synapse loss [5–7]. In addition, studies have revealed that AD model mice develop synaptic deficits in neurons early in the pathogenesis before the appearance of Aβ plaques, which underlie the pathology of cognitive dysfunction [8, 9]. However, the molecular mechanisms of synaptic dysfunction have not been clearly defined within the brains of AD patients.
Considering the complexity of AD pathogenesis, there is no definitive clinical evidence suggesting that validated medicines exist to cure AD, and most of them merely alleviate the clinical manifestations [10, 11]. Since the pathology of AD is a progressive process, studies suggest that controlling modifiable risk factors (including lifestyle, diet, etc.) can be effective in slowing down the progression of AD in an early stage, and perhaps even preventing it in the future [12–14]. Physical activity is one of the most promising modifiable lifestyles. Increasing evidence supported a protective effect of physical activity against AD related cognitive dysfunction [15–18]. A series of randomized controlled trials in humans have demonstrated the direct benefits of physical activity on brain function. These appear to be mediated by changes in white matter integrity [15], cerebral blood flow [16], hippocampal volume [17], and enhanced functional connectivity of the hippocampus [18]. However, the underlying biological basis in the brain tissues by which physical activity improves cognitive function in AD patients was unclear. In this study, integrated transcriptomic analyses and molecular biological methods were used to explore the molecular processes impaired in AD that were conversely preserved and enhanced by physical activity. The results would contribute to a better understanding of the role of physical activity in the prevention and treatment of AD.
MATERIALS AND METHODS
Data acquisition
As shown in Fig. 1, the microarray datasets used in this study were obtained from the GEO database. The search criteria were as follows: (A) the samples were from human brain tissues, (B) gene expression was analyzed, (C) the datasets included AD patients and both elderly people with normal cognition as well as elderly people with varying degrees of physical activity, and (D) all AD patients were diagnosed according to the National Institute of Neurological and Communicative Disorders and the Alzheimer’s Disease and Related Disorders Association criteria. Finally, we extracted GSE110298 and GSE48350 as the target datasets, downloaded the Series Matrix File, and saved it in TXT format. Given that individuals with physical activity measured above 26 METs-h/wk (3.71 METs-h/day) were reported to be the minimal physical activity that would improve cognitive function [19], the transcriptome profiles of 23 hippocampal region samples [low physical activity group (LPA, 11 cases, < 1.5 METs-h/day) and high physical activity group (HPA, 12 cases, > 3.75 METs-h/day)] in the GSE110298 dataset were chosen. The expression profiles of the healthy elderly control group (64–99 years old, 26 cases) and the AD group (60–99 years old, 19 cases) in the GSE48350 dataset were selected for the follow-up study. To subsequently validate hub genes, we downloaded the GSE5281 dataset (n = 23) as a validation dataset for AD patients (70–88 years old, 10 cases) and elderly control people (63–102 years old, 13 cases), and the GSE203554 dataset (n = 15) for AD model mice with or without HPA. The human GEO datasets included in the study were all microarray arrays based on the GPL570 [HG-U133_Plus_2] Affymetrix human genome U133 Plus 2.0 array.
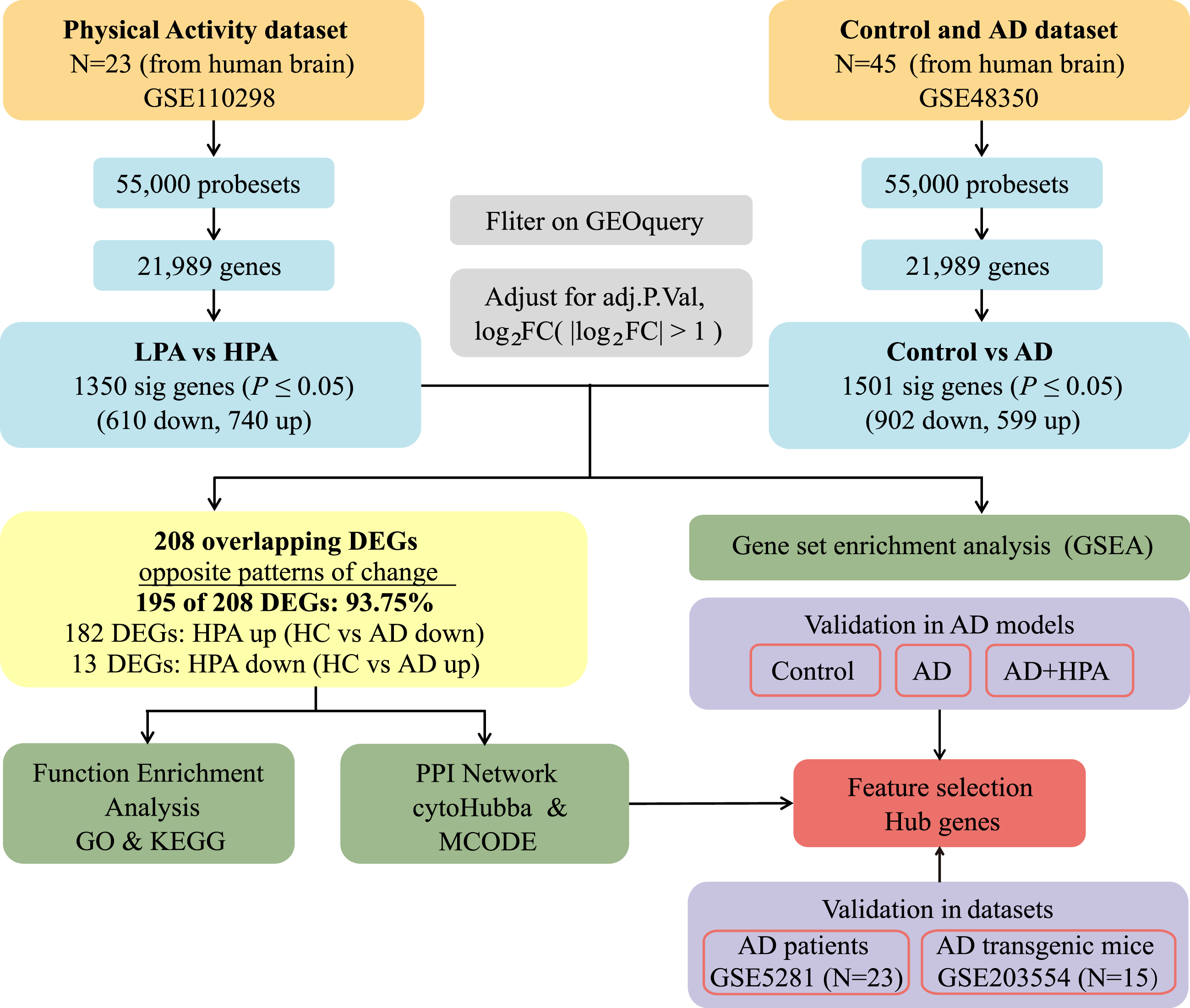
Flow chart of the study design. AD, Alzheimer’s disease; LPA, low physical activity; HPA, high physical activity; FC, fold change; adj.P.Val, adjust p value; DEGs, differentially expressed genes; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; MCODE, Molecular Complex Detection.
Identification of common DEGs
Before the procedure of differential analysis, the datasets were normalized using the “Normalizeweenarrays” function from the limma package (version: 3.56.2) in R (version: 4.3.0) [20], and then we used the GEOquery (version: 2.68.0) and limma package (version: 3.56.2) for analyzing the processed data. The batch effect of data was shown by box plots using the boxplot function in the gplots package (version: 3.1.3), and three-dimensional PCA clustering plots were plotted by the prcomp function in the FactoMineR package (version: 2.8) [21]. The effects of metadata (age or gender) on the principal components were analyzed using the PCAtool package (version: 2.12.0). The cutoff criteria (|log2FC (Fold change) |>1 and p < 0.05) were used to screen the differentially expressed genes (DEGs). Volcano maps and heatmaps of DEGs were then constructed using the R packages ggplot2 (version: 3.4.2) and pheatmap (version: 1.0.12) to visualize the genes. We performed Venn diagram analysis using Venny 2.1.0 (https://bioinfogp.cnb.csic.es/tools/venny/index.html) to identify the common DEGs across physical activities and among ADs. Pearson method in Corrplot packages (version: 0.92) was used to explore the correlation indexes of the expression of common DEG between AD and HPA. Results with p < 0.05 were considered statistically significant.
Function annotation and pathway enrichment of DEGs
Gene Ontology (GO) is a database commonly used in the field of bioinformatics for a wide range of species. GO database characterizes the function of genes or proteins, covering the class of molecular functions, the biological processes facilitated by these functions, and the locations where these occur in the cell [22]. The classical pathway database, Kyoto Encyclopedia of Genes and Genomes (KEGG), gives functional meaning to genes and genomes at the molecular and holistic level, and has been described as “a reference library for understanding the higher functions and utility of biological systems” [23]. Gene set enrichment analysis (GSEA) is an enrichment method that preserves the correlation between genes based on enrichment analysis of all genes in the data, rather than those genes that are above a threshold in terms of fold change or p-value. GSEA includes a variety of annotation files such as GO, KEGG, Reactome, etc., which reveal the role of genes together in health and disease [24]. Enrichment analysis of the GO, KEGG pathway was performed in common DEGs that showed opposite expression trends in AD and HPA using the clusterProfiler package (version: 4.8.2) [25]. The ReactomePA package (version: 1.44.0) [26] was applied to all DEGs for GSEA enrichment analysis. Results with p < 0.05 were considered statistically significant.
Construction of protein-protein interaction network
To explore regulatory interactions between common DEGs that showed opposite expression trends in AD and HPA, we used the STRING (https://string-db.org/) database to search for protein-protein interactions (PPI) networks and then visualized by Cytoscape (version: 3.9.1) for subsequent analysis [27].
Key module analysis and hub gene identified
The plugin in Cytoscape, Molecular Complex Detection (MCODE), can be applied to discriminate functional modules in PPI networks through the search for gene clusters by exploiting the inherent relationships between proteins in the network [28]. Here we used the default parameters (Node Score Cutoff = 0.2, Degree Cutoff = 2, k-Score=2, Maximum Depth = 100) for the analysis. To extract hub genes from the PPI network, we performed network topology analysis on the cytoHubba, which is a plugin of Cytoscape, scored and ranked all the DEGs through three methods, MCC, MNC, and Degree, and filtered the top 10 node genes [29]. The reliability of hub genes was further analyzed using the validation datasets. The ggplot2 package (version: 3.5.0) was used to draw validation boxplots.
Animals
Breeding pairs of APPswe/PSENldE9 transgenic mice were originally purchased from Jackson Laboratories, USA. A breeding colony of APPswe/PSENldE9 mice was established at Ningbo University Laboratory Animal Centre in an SPF-grade environment with temperature: 22±3°C, humidity: 55% ±5%, and day/night alternation time. Seven-month-old male APPswe/PS1dE9 (APP/PS1) transgenic mice and wild-type (WT) mice were subjected to a series of training progress and fed freely in cages with water and a diet of about 8 g per day. The procedures of experimental animals were based on the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85–23, Revised 1996) and approved by the Animal Care and Use Committee of the School of Medicine, Ningbo University, Ningbo, China (No. NBU20230158). Finally, the mice were sacrificed at about eight months.
Training program
The HPA training program was executed according to a previously described program which contained an adaptation stage followed by four weeks formal training [30, 31]. The program has been demonstrated to effectively prevent the cognitive impairment of AD model mice. As shown in Fig. 2, the first three days were the adaptation stage, and the mice in the HPA group were trained on the treadmill at the speed of 5 m/min for 10 min on the first day, 8 m/min for 20 min on the second day, and 10 m/min for 30 min on the third day, with a rest of 2 min for every 10 min training. Then the mice were treated with the four-week formal training, which was conducted at the speed of 10 m/min for 40 min, with a rest of 2 min for every 10 min training. The training took place 5 days a week (Monday to Friday).
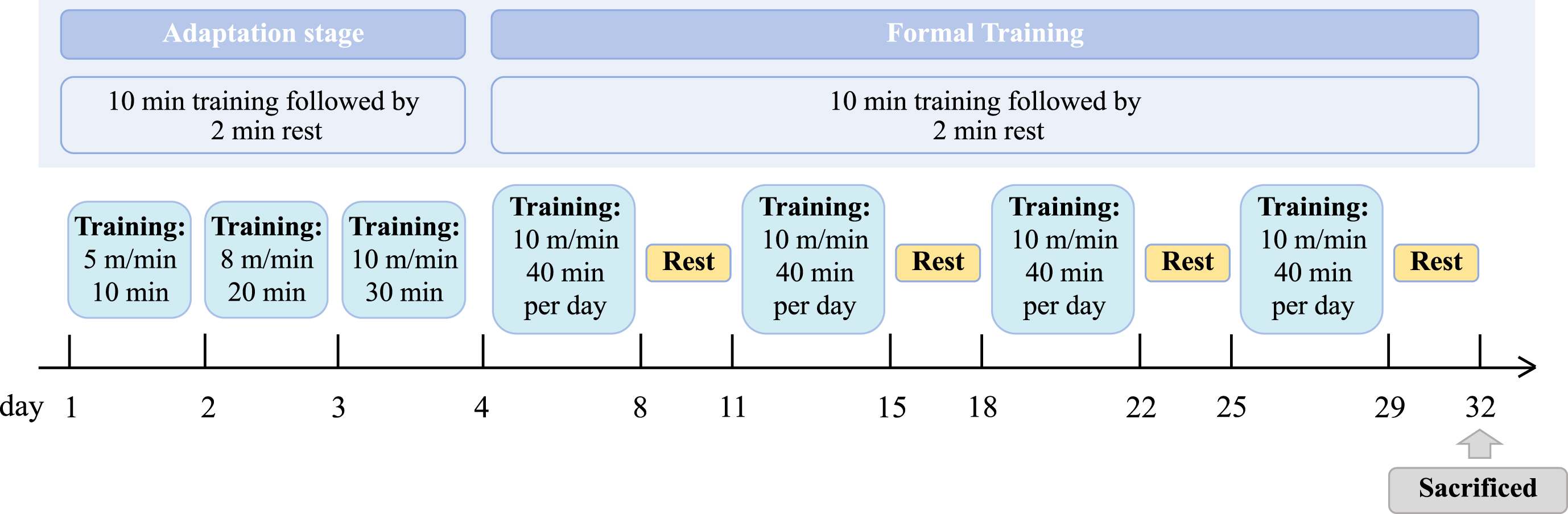
The HPA training procedures of mice. The HPA training program was executed with an adaptation stage followed by four weeks formal training. The first three days were the adaptation stage. Then the mice were treated with the four-week formal training, which was conducted at the speed of 10 m/min for 40 min, with a rest of 2 min for every 10 min training. The training took place 5 days a week.
Real-time quantitative reverse transcription PCR (qRT-PCR)
Total RNA was extracted from the prefrontal cortex and hippocampus tissues of mice in each group using TRIzol Reagent (Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) according to the manufacturer’s instructions. cDNA was synthesized using TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix kit (TransGEN, Beijing, China) at 1μg of total RNA per reaction. Relative mRNA levels were quantified by real-time fluorescence quantitative PCR using the PerfectStart Green qPCR SuperMix (TransGEN, Beijing, China) with the specific primers (Table 1).
Nucleotide sequences, amplification sites, and GenBank accession numbers for each primer pair
Statistical analysis
Statistical analysis was performed with GraphPad Prism software (GraphPad Software Inc., La Jolla, CA, USA, RRID: SCR_00298). The data were presented as mean±SEM. Two groups data were analyzed using Student’s t-test. Multiple groups data were analyzed using one-way analysis of variance (ANOVA) followed by a Tukey post hoc test. p < 0.05 was considered statistically significant.
RESULTS
Datasets had been normalized and the source of samples was reliable
The data from the GSE110298 and GSE48350 were normalized. No batch effect in the data from the GSE110298 and GSE48350 was found (Fig. 3A, C). Principal component analysis was applied to explore whether the source of samples was reliable. The results showed that the samples of AD and control groups were non-overlapping and exhibited a significant difference (Fig. 3B). The samples of the physical activity groups with varied intensity were partially overlapped, indicating that there were also certain discrepancies between the LPA and HPA groups (Fig. 3D). These results suggested that the source of samples was reliable. The related clinical information of the samples from different groups were also analyzed. No statistical difference in age and sex was found between AD and control groups or between different physical activity groups (Tables 2 and 3). We further analyzed the effects of age or gender on the principal components. No significant difference in the correlation indexes between age/gender and the principal components was found (Fig. 3E, F).
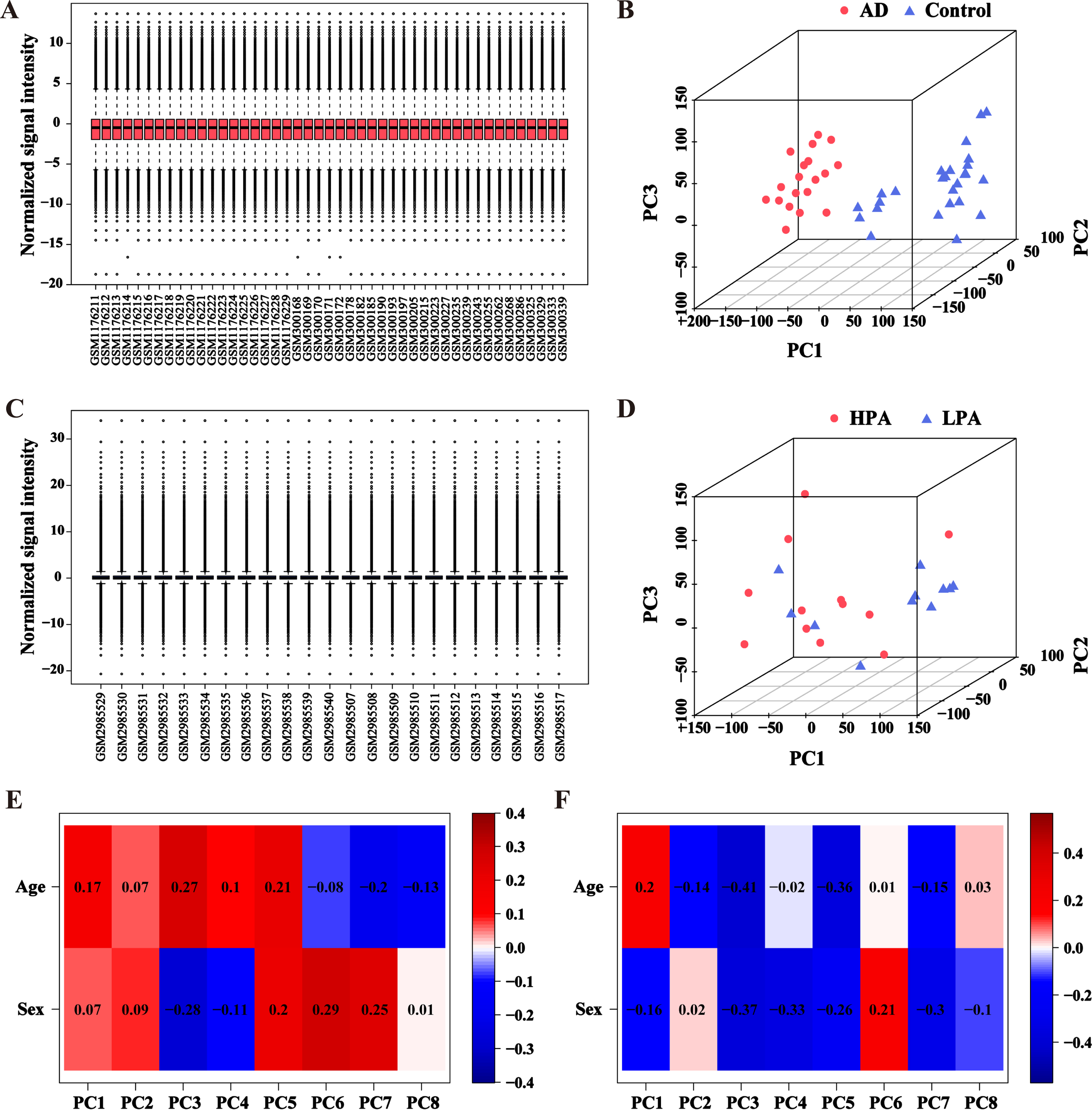
PCA plots and boxplots of normalized data from AD and different physical activity datasets. A) Boxplot for AD and control dataset after normalization. B) PCA plot for AD and control dataset after normalization. Samples from the AD group and control group were represented by red dots and blue triangles, respectively. C) Boxplot for different physical activity dataset after normalization. D) PCA plot for different physical activity dataset after normalization. Samples from the HPA group and LPA group were represented by red dots and blue triangles, respectively. E) Eigencor plot for the correlation indexes between age/gender and the principal components in AD dataset. F) Eigencor plot for the correlation indexes between age/gender and the principal components in different physical activity dataset. PC, principal components.
Clinical characteristics of the AD and control dataset
Clinical characteristics of the different physical activity dataset
The DEGs of elders with high physical activity and AD patients showed opposite trends
The DEGs of elders with high physical activity and AD patients were found. In accordance with the criteria |log2FC| > 1 and p < 0.05, 1350 DEGs, including 740 upregulated and 610 downregulated genes, were extracted between AD and control groups (Fig. 4A). Similarly, 1501 DEGs (599 upregulated and 902 downregulated) were extracted between LPA and HPA groups (Fig. 4B). The heatmap displayed the top 50 genes with remarkable discrepancy between the two datasets (Fig. 4C, D). After excluding genes with common expression trends among AD group and different physical activity groups through cross-comparison analysis, we identified 195 common DEGs, including 13 genes upregulated and 182 downregulated genes in AD group (Fig. 4E). The top 6 downregulated common genes in AD group while were upregulated in HPA group were CALB1, PDYN, GABRG2, LAMP5, NWD2, and MAL2 (Fig. 4F). The top 8 upregulated common genes in AD group which showed an opposite trend in HPA group were SFPQ, NFE2L2, SRGAP1, CYYR1, HIPK2, RAB30, SERPINA5, and CDK19 (Fig. 4F). The correlation indexes of the expression of these DEG between AD and HPA were calculated, and the expression levels of NFE2L2 and LAMP5 in the HPA group were significantly negative correlated with those of the AD group (Fig. 4G, p < 0.05).
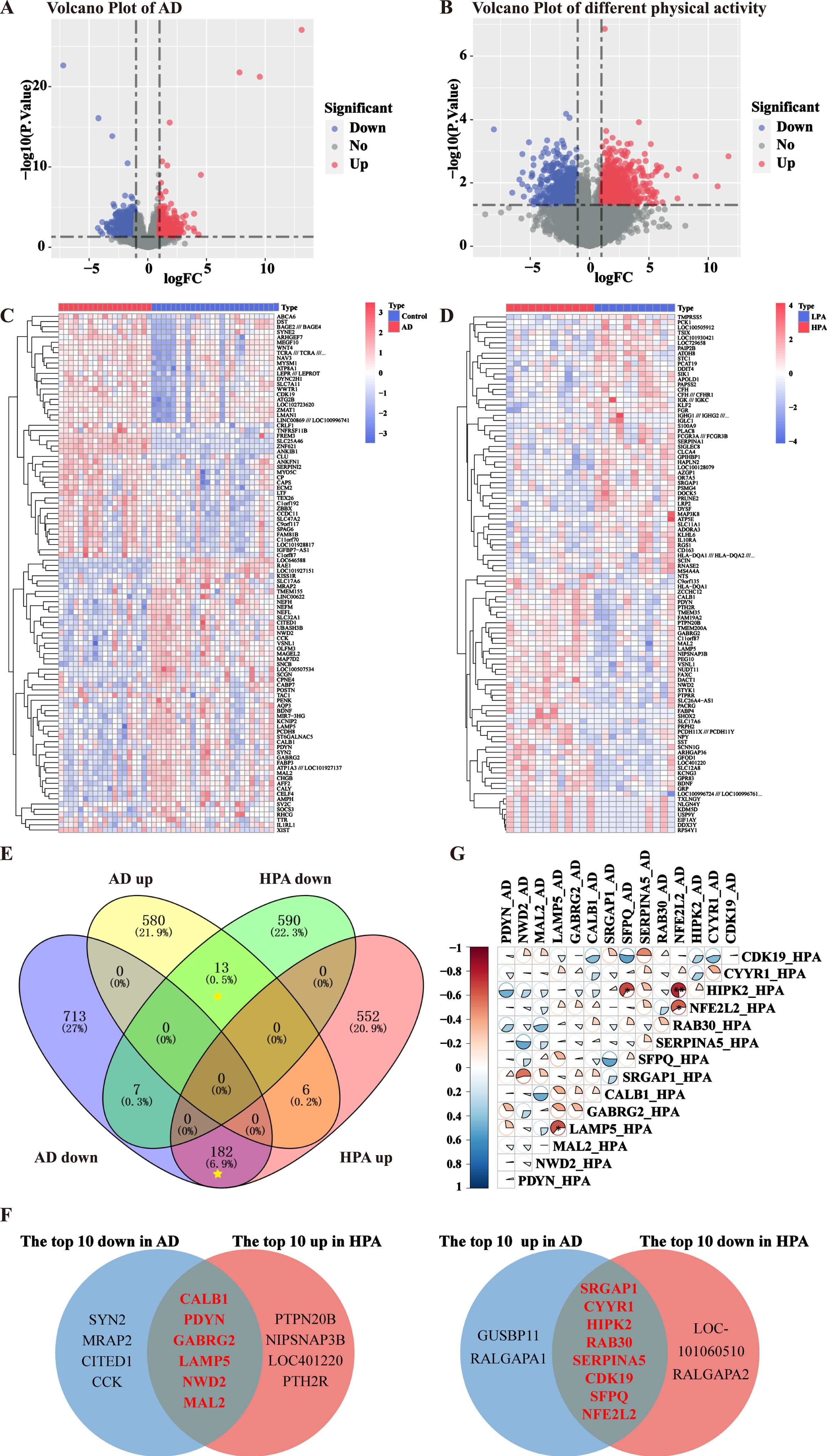
The DEGs of elders with high physical activity and AD patients showed opposite trends. A) Volcano plots of AD dataset. Upregulated genes were marked in red, downregulated genes were marked in blue, and grey dots represent non-differentiated genes. B) Volcano plots of different physical activity dataset. C) Heatmap of the top 50 upregulated DEGs and downregulated DEGs in AD dataset. D) Heatmap of the top 50 upregulated DEGs and downregulated DEGs in different physical activity dataset. E) DEGs of elders with high physical activity and AD patients were displayed in Venn diagram. 195 common DEGs after excluding genes with the same expression trend (yellow asterisks) were found. F) The top 10 upregulated common DEGs and downregulated common DEGs in both datasets were shown in Venn diagrams. There were six genes downregulated in AD group while were upregulated in HPA group, and eight genes upregulated in AD group which showed an opposite tendency in HPA group. G) The correlation indexes of the expression of the DEG between AD and HPA. Red (negative correlation), blue (positive correlation), sector direction (clockwise for positive correlation, and counterclockwise for negative correlation). The darker the colors, the larger the sector areas, and the higher the correlations. **p < 0.01, *p < 0.05.
Function annotation and pathway enrichment of DEGs
Differential genes were analyzed using three widely used databases for excavating potential biological features, including GO, KEGG pathway, and GSEA. GO enrichment analysis was used to annotate gene products and identify molecular features associated with genes. In terms of biological processes, the top 10 biological processes were visualized with bubble plots. AD and physical activity-associated DEGs were mainly enriched in synaptic signaling and regulation, such as synaptic organization, vesicle-mediated transport, synaptic vesicle cycle, and exocytosis (Fig. 5A, Table 4, p < 0.05). According to cellular components, glutamatergic synapse, neuron-to-neuron synapse, synaptic membrane, and postsynaptic specialization were the most important compositions involved (Fig. 5A, Table 4, p < 0.05). For pathway enrichment analysis, chordal plots revealed that these common DEGs were predominantly enriched in synaptic vesicle cycle, endocrine and other factor-regulated calcium reabsorption, steroid biosynthesis, Ras signaling pathway, and gap junctions (Fig. 5B, Table 5, p < 0.05). The genes got enriched in at least 2 KEGG pathways were PDYN, NPY, TUBA4A, GNG3, TUBB2A, MAP2K1, AP2S1, AP2M1, and COX6 C (Fig. 5B, Table 5). Further independent analysis of DEGs suggested that the above GO categories and pathways were downregulated in the majority in AD, which is consistent with the general notion that synaptic function is defective in AD patients. These GO categories and pathways were mostly found to be upregulated in the HPA group, suggesting that HPA can improve the function of synapse.
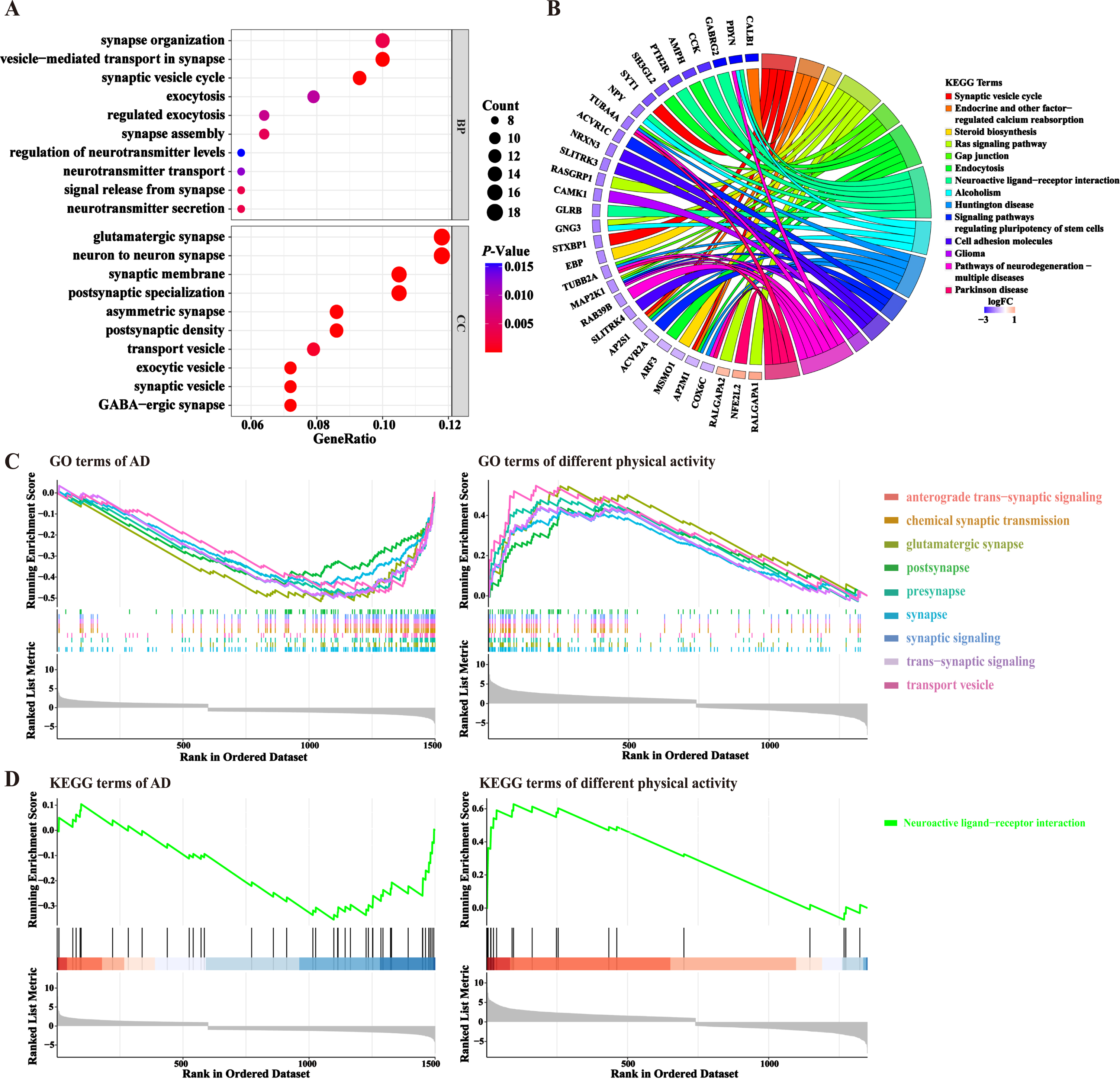
Functional annotation and pathway enrichment of DEGs. A) GO enrichment analysis of common DEGs. The top 10 categories with the most significant p-values, sorted by gene ratio were shown by bubble plots. BP, biological process; CC, cellular component. The horizontal axis indicates the proportion of differential genes in each GO category. B) KEGG enrichment analysis of common DEG. The top 10 KEGG pathways that have significant p-values and the DEGs being enriched were demonstrated by Chordal plots. Different colors corresponded to different pathways. C, D) GSEA enrichment analysis of the two datasets. The common GO (C) and KEGG (D) categories enriched to both datasets were shown here.
GO analysis results of common DEGs. The top ten enriched GO terms of each category were
BP, biological process; CC, cellular component.
Pathway enrichment analysis of common DEGs. The top 10 enriched pathways of the KEGG
In addition, the GO terms and KEGG pathways of the respective differential genes in both AD and physical activity datasets were further analyzed by GSEA enrichment (Fig. 5C, D). The results showed that anterograde trans-synaptic signaling, chemical synaptic transmission, glutamatergic synapse, and neuroactive ligand-receptor interaction pathway might be the important processes that were regulated by HPA. These synapse transmission related genes were downregulated in AD group, while they were upregulated in HPA group (Fig. 5C, D).
Construction of PPI network and hub genes screening
The PPI network consists of proteins through their interactions with each other, which is important for understanding the functioning principles of proteins in biological systems. This is a key step in cell biology research and crucial for studying the response mechanisms of biological signals and energy metabolism in particular physiological states like diseases. We constructed a PPI network of 195 DEGs from AD and HPA groups using the STRING database and visualized it in Cytoscape (Fig. 6), which consists of 104 nodes and 150 edges. Additionally, we used the MCODE plugin to perform a functional module analysis of this network, and the top two closely connected clusters were shown (Fig. 7A). Cluster 1 consisted of 9 nodes and 17 edges, including SYT1, SYT4, SH3GL2, GABRG2, AMPH, SNAP91, SYNPR, LIN7B, and NRXN3. Cluster 2 had 3 nodes and 3 edges, with AP2M1, AP2S1, and NECAP1. Clusters 1 and 2 were subjected to GO enrichment analysis via the ClueGO plugin, and we found that cluster 1 was mainly associated with synaptic vesicles and cluster 2 was concerned mainly with clathrin vesicle (Fig. 7B). Three widely used sorting methods (MCC, MNC, and Degree) were chosen to score the PPI network using the cytoHubba plugin. The top 10 genes for each algorithm were demonstrated (Fig. 7C). Then, to explore the hub genes that play indispensable roles in the biological mechanisms underlying the neuroprotective effects of HPA, we cross-referenced these genes with genes present in functional gene modules constructed by MCODE. Finally, we identified four common genes as pivotal genes, including SH3GL2, AP2M1, SYT1, and SYT4. We validated these hub genes in AD transgenic mice with or without HPA dataset (GSE203554) and AD patients dataset (GSE5281). No batch effect was found after normalizing the two datasets (Supplementary Figure 1). The age and sex were not significantly different between the AD and control groups (Supplementary Table 1). The expression of the hub genes was significantly decreased in AD patients or AD transgenic mice (Fig. 7D, E, p < 0.05). HPA treatment significantly increased the expression of these hub genes in AD mice (Fig. 7E, p < 0.05).
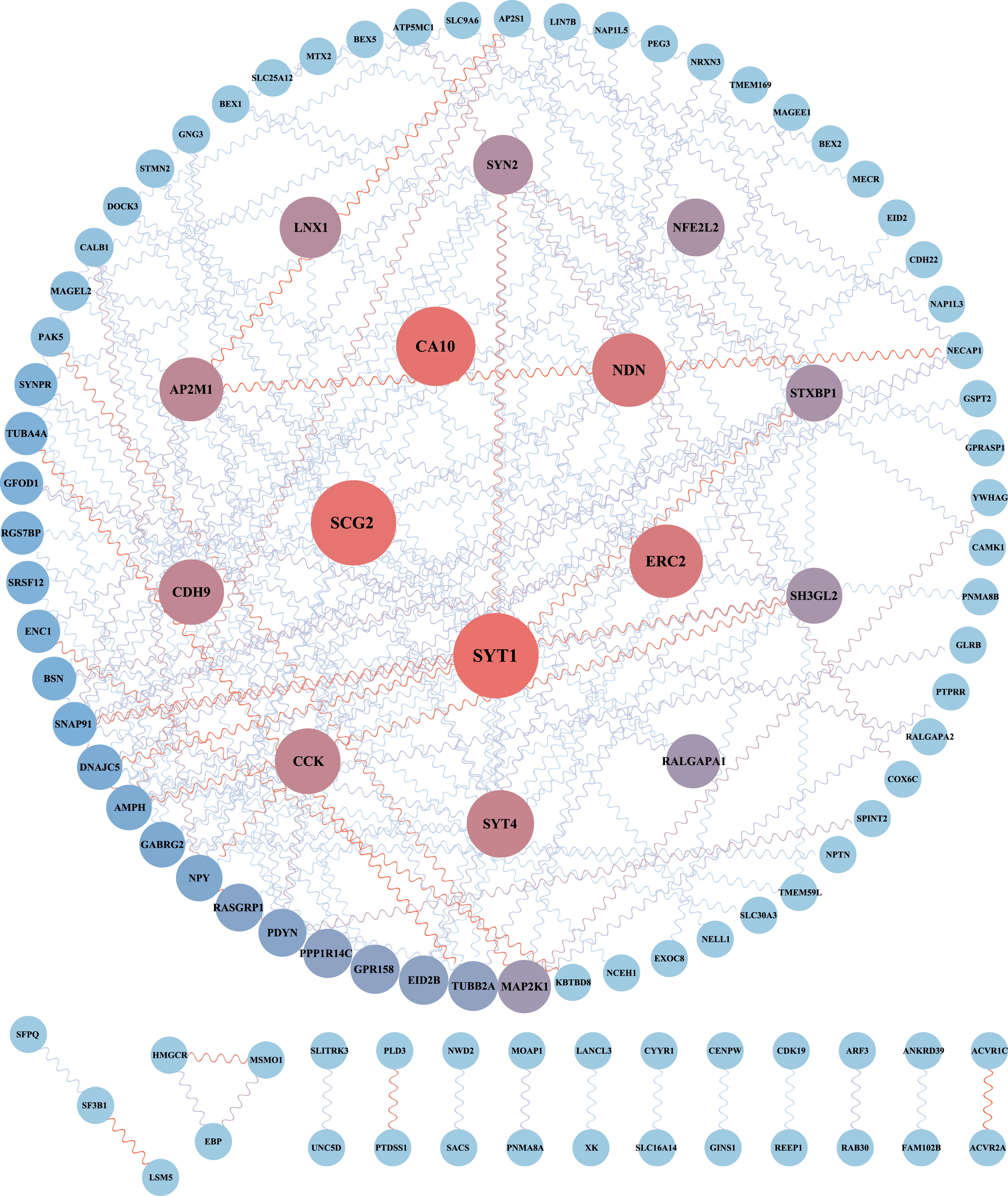
PPI network of common DEGs between different physical activities and AD. The network consists of 104 nodes and 150 edges, was generated using the String database and visualized in Cytoscape. Circular nodes denoted DEGs and edges denoted interactions between nodes; node size and color shades were based on Betweenness Centrality derived from the cytoCNA plugin in Cytoscape. The larger the node, the higher the algorithm score.
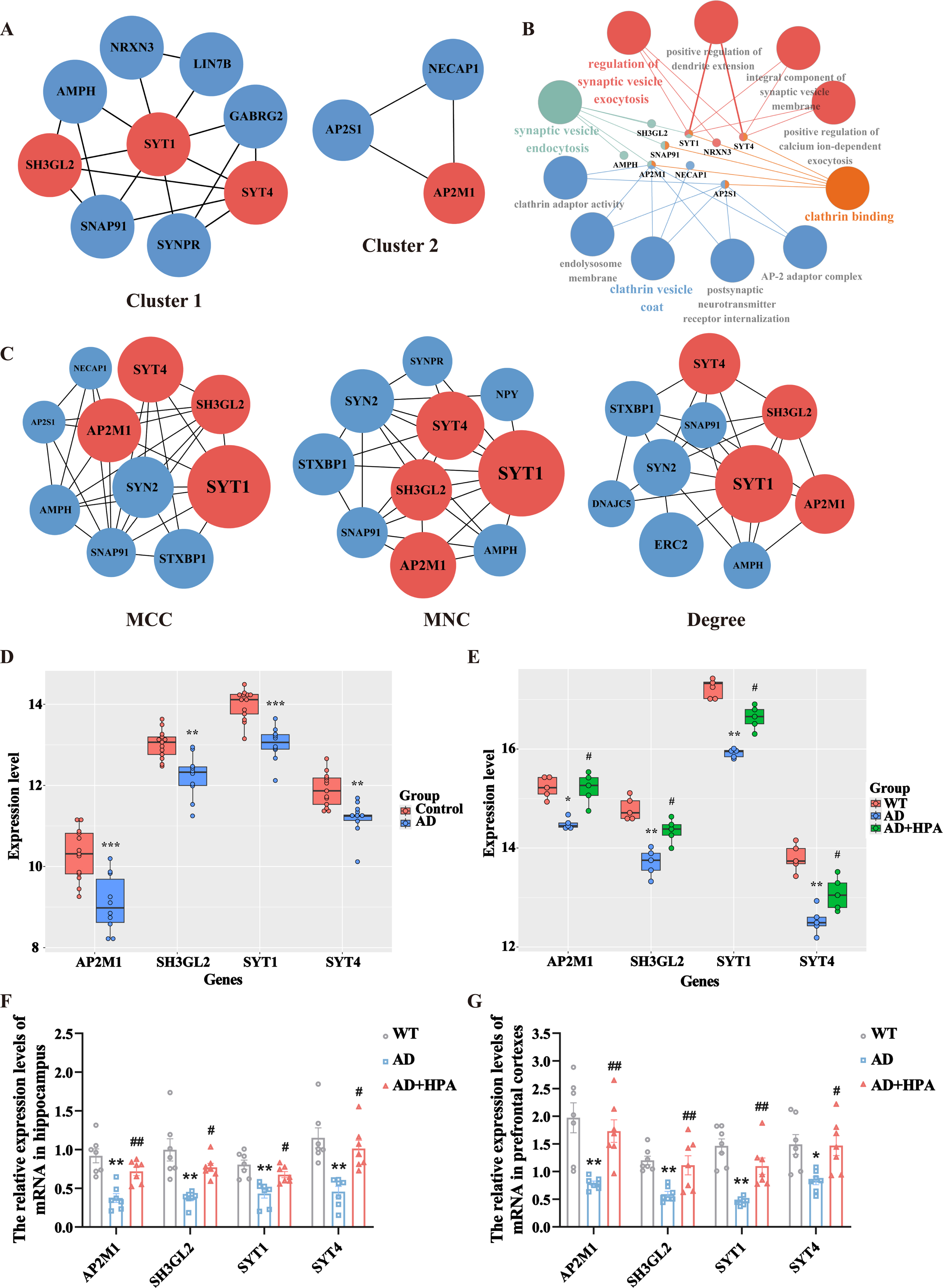
Hub gene screening and key module analysis of the PPI network. A) The two highest-scoring gene clusters were extracted from the PPI network using the MCODE plugin in Cytoscape. B) GO annotation of key modules. C) The top 10 genes were constructed by MCC, MNC, and Degree algorithms using the Cytohubba plugin, respectively. The larger the node, the higher the algorithm score. The red nodes represented the hub genes, which were common genes that intersected with the key modules. D) The verification of the hub genes in AD patients dataset (GSE5281). ***p < 0.001 versus control group, **p < 0.01 versus control group, N = 10-13. E) The verification of the hub genes in AD transgenic mice with or without HPA dataset (GSE203554). **p < 0.01 versus WT group, *p < 0.05 versus WT group, #p < 0.05 versus AD group, N = 5. F, G) Validation of hub gene transcript levels in the hippocampal tissues (F) and prefrontal cortexes (G) in different groups. **p < 0.01 versus WT group, *p < 0.05 versus WT group, # #p < 0.01 versus AD group, #p < 0.05 versus AD group, N = 7.
The decreased mRNA expression of hub genes in APP/PS1 mice were significantly reversed by physical activity
It is difficult to obtain the brain tissues of AD patients with and without physical activity. We further verified the expression of four hub genes in the hippocampus and prefrontal cortexes of AD transgenic mice which were treated with or without HPA by qRT-PCR. The mRNA expression levels of AP2M1, SH3GL2, SYT1, and SYT4 were significantly decreased in APP/PS1 mice compared with those in the WT group, and HPA reversed the decreased expression levels of synaptic function related genes in the AD transgenic mice (Fig. 7F, G, p < 0.05).
DISCUSSION
Numerous clinical trials in humans supported a protective effect of physical activity against AD related impairments which reflected in higher MMSE scores, increased psychomotor speed, and reduced hippocampal atrophy [32–34]. In this study, we tried to explore the underlying biological basis by which physical activity improved cognitive function in AD patients and provided evidence for physical activity as an effective prevention and treatment for AD.
Through the transcriptomic analysis research, a total of 195 common DEGs, including 13 genes downregulated and 182 upregulated genes in HPA group, were identified in the current study. This result indicated that physical activity might cause an activation of cellular functions in patients’ hippocampus. The cross-comparison analysis suggested the top 8 upregulated common genes in AD group which showed an opposite trend in HPA group were SFPQ, NFE2L2, SRGAP1, CYYR1, HIPK2, RAB30, SERPINA5, and CDK19. These genes were primarily enriched for transcriptional regulation genes or kinase [35–39]. Dysregulation of these proteins will cause hyperphosphorylation of Tau protein, dysregulation of vesicle turnover, and then impairment of synaptic function and cognition [35, 39]. The top 6 significantly downregulated common genes in AD group while upregulated in HPA group were CALB1, PDYN, GABRG2, LAMP5, NWD2, and MAL2. These genes were important genes that are related to synapse transmission and synapse function. For example, MAL2 encodes a multi-spanning transmembrane protein that is a member of the polarized transport machinery which has been shown to be required for membrane components of synaptic vesicles [40]. Consistent with our results, the expression of MAL2 was downregulated in the cortex of 8-month-old AD mice [41]. GABRG2, a GABA subunit necessary for postsynaptic localization and aggregation, is closely related to synaptic transmission function and genetic mutations of which are often reported in AD [42, 43]. Meanwhile, CALB1 is one of the calbindin proteins which plays a physiological role in buffering cytosolic calcium. Several studies have revealed that increased levels of CALB1 in the hippocampal dentate gyrus improve spatial memory through increasing synaptic activity in a mouse model of AD [44, 45]. Lysosome-associated membrane protein 5 (LAMP5) deserves as a sorting factor for localization of the vesicular GABA transport to synaptic vesicles. The loss of synaptic LAMP5 in AD mice brains was proven to impair learning and memory formation [46]. The dysregulation of these genes in AD and the possibility that physical activity may reverse the expression of these genes would represent the underlying mechanism that could have far-reaching consequences for hippocampal function.
Further, GO enrichment analysis of common genes underlines synaptic signaling processes to be suppressed in AD but conversely upregulated by physical activity. These genes were mainly involved in the formation, release, transport, and recycling of vesicles. At the same time, the KEGG pathway enrichment was consistently enriched to the pathway of the synaptic vesicle cycle. Furthermore, the GSEA of the two datasets also demonstrated that synapses, trans-synaptic signaling, transport vesicle, and neuroactive ligand-receptor interaction were upregulated after increasing the intensity of physical activity. The role of synaptic vesicles in transmitting neurotransmitters requires a complex transport cycle divided into synaptic vesicle formation, transporter protein-mediated filling of synaptic vesicles, docking, initiation, and fusion of synaptic vesicles at the active zone of the presynaptic membrane, release of neurotransmitters and endocytosis of vesicles [47]. Our results showed that synaptic vesicle transport and docking relative proteins were decreased in AD patients and physical activity had a positive effect on these aspects of the synaptic vesicle cycle, thereby facilitating inter-neuronal information transfer. In line with our results, the research showed that the expression of synaptic vesicle transport and docking relative proteins such as synaptotagmin, synaptophysin, and syntaxin was increased in postmortem brain tissues of older adults with HPA in late life [48].
Based on four different topological methods of PPI network analysis, we identified two key modules and four hub genes. The GO pathway analysis of the key modules was consistent with the previous results and highlighted the improvement of synaptic vesicle release and recycling as the main molecular mechanisms by which physical activity improves cognitive dysfunction in AD patients. Four important genes SYT1, SYT4, SH3GL2, and AP2M1 were included in two key modules. Our results indicated the expression of the four genes was decreased in an AD mouse model and HPA reversed the decreased expression of these four genes in the AD mouse model. The four hub genes are associated with activity-dependent synaptic vesicle cycling, which is important for synaptic plasticity and connectivity [49, 50]. SYT1 is a synaptic binding protein that regulates synaptic vesicle exocytosis in a calcium-dependent manner [51]. It has been demonstrated that n-cadherin mediates transsynaptic signaling for presynaptic vesicle turnover through increasing SYT1 [52]. Besides the regulation of presynaptic vesicle turnover, SYT1 was reported to inhibit the progerin gene (PS1), which is one of the components of the γ-secretase enzyme [53–55]. The combination of SYT1 and PS1 was reduced in primary neurons of AD mice [53, 55]. SYT4 is also a member of the synaptic binding protein family and binds to SNARE [soluble NSF (N-ethylmaleimide-sensitive factor) attachment protein receptors] in a calcium-independent manner [56]. Consistent with our results, the research demonstrated that reduction in SYT4 expression decreases the probability of kiss-and-run release, slowing down the efficiency of cellular recycling of vesicles [57]. Meanwhile, studies revealed that postsynaptic cells can mediate activity-dependent retrograde signaling into the synaptic gap by releasing SYT4 [58, 59]. The AP2M1 and SH3GL2 genes respectively code the μ-subunit of the clathrin junction complex AP2 and the membrane-bound protein endorphin 1 [60, 61]. They are involved in clathrin-mediated endocytosis (CME), a major pathway for internalization of plasma membrane proteins and a major mechanism for cycling the components of the mammalian synaptic vesicle (SV) [62]. The research showed that missense variants of AP2M1 and the absence of SH3GL2 gene-encoded product impaired CME which led to neurodevelopmental abnormalities and impairment of synaptic transmission [60, 63]. In addition, it was noted that the AP2 complex mediates retrograde transport of autophagosomes to increase neuronal complexity and prevent neurodegeneration in mouse hippocampus [64]. Our results indicated that HPA might reverse the impairments in AD patients and AD model mice by increasing the expression of SYT1, SYT4, AP2M1, and SH3GL2 genes and improving the efficiency of synaptic vesicle turnover. Strengthening the intensity of physical activity may serve as a preventive strategy for AD.
Other mechanisms such as decreased amyloid load, reduced tau phosphorylation and neuroinflammation might also be involved in the prevention of AD pathogenesis through HPA [65–67]. AD transgenic rats with HPA showed attenuated Aβ deposition and tau hyperphosphorylation [65]. Lower brain amyloid was found in AD patients with HPA through PET imaging and CSF biomarker analysis [66]. A previous study demonstrated that AD patients with HPA were correlated with a reduction of pro-inflammatory microglial states [67]. Nevertheless, alterations in synaptic vesicle turnover and synaptic strength might be a key mechanism of the beneficial effects of HPA in the early stage of AD. Reduced levels of synaptic proteins have been detected in mild cognitive impairment (MCI) patients [68]. The presynaptic terminal is particularly vulnerable and represents a critical site for the manifestation of initial AD etiology [69]. Positive associations between physical activity and the synaptic proteins were found in all three diagnoses, including no cognitive impairment, MCI, and dementia, the effect size was strongest in participants diagnosed with MCI [48]. Regulation of the synaptic strength and function might be the same way through which physical activity might improve cognition in the aged population and AD patients. Physical activity interventions may have optimal therapeutic windows for synaptic outcomes and be particularly effective in individuals in the early stage of cognitive impairment [48].
In summary, through integrated transcriptomic analysis and molecular biological method, we found that physical activity may reverse AD pathology by upregulating a range of synaptic vesicle transport related proteins which might improve the efficiency of synaptic vesicle turnover and facilitate inter-neuronal information transfer. The study provides novel insights into the mechanisms underlining the protective effects of high physical activity on AD and a better understanding of physical activity as an early treatment and prevention of AD.
AUTHOR CONTRIBUTIONS
Can Wu (Data curation; Formal analysis; Investigation; Methodology; Project administration; Validation; Visualization; Writing – original draft; Writing – review & editing); Tingting Ruan (Data curation; Formal analysis; Methodology; Visualization); Yalan Yuan (Formal analysis; Methodology; Visualization); Chunshuang Xu (Validation); Lijuan Du (Investigation; Validation); Fang Wang (Data curation; Methodology); Shujun Xu (Funding acquisition; Investigation; Project administration; Writing – original draft; Writing – review & editing).
Footnotes
ACKNOWLEDGMENTS
We would like to thank the Core Facilities, Ningbo University School of Medicine for the technical support, and Ningbo University Laboratory Animal Centre, China for assisting with the study. We acknowledge the GEO databases for providing their platforms and contributors for uploading meaningful datasets.
FUNDING
This work was supported by grants from the National Natural Science Foundation of China (81771166), Natural Science Foundation of Zhejiang Province (LY23H090005, LQ23H090003), Ningbo Key Research and Development Plan Project (2023Z173, 2022Z147), Natural Science Foundation of Ningbo (2022J118, 2022J250, 2021J250), Zhejiang Medical and Health Technology Project (2023KY1135), and the K. C. Wong Magna Fund in Ningbo University.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.