
Abstract
Background:
TAR DNA binding protein 43 (TDP-43) has been shown to be associated with whole hippocampal atrophy in primary age-related tauopathy (PART). It is currently unknown which subregions of the hippocampus are contributing to TDP-43 associated whole hippocampal atrophy in PART.
Objective:
To identify which specific hippocampal subfield regions are contributing to TDP-43-associated whole hippocampal atrophy in PART.
Methods:
A total of 115 autopsied cases from the Mayo Clinic Alzheimer Disease Research Center, Neurodegenerative Research Group, and the Mayo Clinic Study of Aging were analyzed. All cases underwent antemortem brain volumetric MRI, neuropathological assessment of the distribution of Aβ (Thal phase), and neurofibrillary tangle (Braak stage) to diagnose PART, as well as assessment of TDP-43 presence/absence in the amygdala, hippocampus and beyond. Hippocampal subfield segmentation was performed using FreeSurfer version 7.4.1. Statistical analyses using logistic regression were performed to assess for associations between TDP-43 and hippocampal subfield volumes, accounting for potential confounders.
Results:
TDP-43 positive patients (n = 37, 32%), of which 15/15 were type-α, had significantly smaller whole hippocampal volumes, and smaller volumes of the body and tail of the hippocampus compared to TDP-43 negative patients. Subfield analyses revealed an association between TDP-43 and the molecular layer of hippocampal body and the body of cornu ammonis 1 (CA1), subiculum, and presubiculum regions. There was no association between TDP-43 stage and subfield volumes.
Conclusions:
Whole hippocampal volume loss linked to TDP-43 in PART is mainly due to volume loss occurring in the molecular layer, CA1, subiculum and presubiculum of the hippocampal body.
Keywords
INTRODUCTION
Primary age-related tauopathy (PART) describes a pathology that combines a limbic predominant distribution of neurofibrillary tangles (NFTs) in the absence or limited distribution of amyloid-β (Aβ) [1]. PART is diagnosed when the Braak NFT stage [2] is I-V and the Aβ Thal phase is 0 (Definite PART) or Aβ Thal phase is 1–2 (Possible PART) [3]. PART has been associated with variable cognition ranging from normal cognition to dementia, with the latter previously being referred to as ‘tangle-predominant senile dementia’ (TPSD), ‘tangle-only dementia’, ‘preferential development of NFT without senile plaques’, and ‘senile dementia of the neurofibrillary tangle type’ (SD-NFT) [1]. Studies show that patients with PART have atrophy of the hippocampus which was presumed to be due to accumulation of the protein tau in the form of NFTs [4–6].
The transactive response DNA-binding protein of 43 kDa (TDP-43) is present in different neurodegenerative diseases. It was first linked to frontotemporal lobar degeneration and amyotrophic lateral sclerosis [7], but later was shown to be present in a significant percentage of cases of with Alzheimer’s disease, as well as in other neurodegenerative diseases [8–10]; some researchers have advocated for the label limbic predominant age-related TDP-43 encephalopathy neuropathologic changes or LATE-NC [11]. In recent studies, we reported finding an association between TDP-43 and whole hippocampal volume loss in cases of PART [12, 13].
Studies on Alzheimer’s disease have shown that TDP-43 is associated with volume loss of specific hippocampal subfields, which together contributes to whole hippocampal volume loss [2, 15]. In addition, TDP-43 burden has been shown to be associated with inward deformation of the hippocampus in zones approximating CA1 and subiculum in AD [16]. In PART, similar to Alzheimer’s disease [9, 17], TDP-43 pathology starts in the medial temporal lobe and progresses to the hippocampus and neocortex [2]. Although there are studies that have investigated the distribution and burden of TDP-43 pathology in hippocampal subfields, volume loss in specific subfields of the hippocampus associated with TDP-43 in PART have not been investigated.
Consequently, the lack of knowledge of this relationship prompted us to investigate the associations between hippocampal subfields and TDP-43 in PART, to ascertain the specific hippocampal subfields that contribute to the association between TDP-43 and the overall loss of hippocampal volume. Results from this study will increase our understanding of TDP-43 and PART pathophysiology and highlight specific hippocampal subfields as potential targeted imaging biomarkers of TDP-43 pathology. We hypothesized that patients with PART and TDP-43 will exhibit smaller volumes in the CA1, subiculum and dentate gyrus of the hippocampus since these are the areas primarily affected by TDP-43 deposition.
METHODS
Study population
We identified 115 cases that met the criteria for possible or definite PART from a cohort of 1,720 autopsy confirmed cases who had been enrolled and followed longitudinally in the Mayo Clinic Alzheimer Disease Research Center, the Mayo Clinic Study of Aging (ADRC/MCSA), or the Neurodegenerative Research Group (NRG), completed at least one antemortem GE volumetric head MRI scan, died and underwent a comprehensive brain autopsy between January 1, 1999 and December 31, 2022. All MRI scans had been completed between this time period.
Pathological analyses
All 115 cases underwent standardized neuropathological examinations, including tissue sampling and semi-quantitative assessment of Alzheimer’s disease pathology following the National Institute on Aging and Alzheimer’s Association (NIA-AA) criteria [12, 13]. Each case was assigned a Braak NFT stage and a Thal Aβ stage in accordance with NIA-AA recommendations. Braak stages I and II correspond to NFT involvement primarily in the transentorhinal and entorhinal regions of the brain; Braak NFT stages III–IV are indicative of spread to the limbic regions, including the hippocampus; and Braak stages V–VI are characterized by extensive NFT involvement of the neocortex [2]. The Thal stages were utilized to categorize the distribution of Aβ deposition: Phase 0 = absence of Aβ throughout the entire brain; Phase 1 = presence of Aβ in the neocortex; Phase 2 presence of Aβ in allocortex/limbic areas; Phase 3 presence of Aβ in the diencephalon/basal ganglia; Phase 4 presence of Aβ in the brainstem/midbrain; and Phase 5 presence of Aβ in the cerebellum [18].
For this study we only included cases with Thal phases 0, 1, and 2, and Braak NFT stages I–IV which meets published inclusion criteria for PART [1, 3]. Hence, we excluded cases with Braak stage 0, V, and VI and Thal phase 3–5.
TDP-43 assessment
For all 115 cases, amygdala and hippocampal samples, from the left hemisphere, were sectioned and paraffin blocks created. The right hemisphere was frozen at the time of brain harvesting. Blank slides were then immunostained to detect TDP-43 using a conformation specific antibody that recognizes the C-terminal fragment of TDP-43 (MC2085; gift from Leonard Petrucelli) [7]. Immunostaining was done using a DAKO-Autostainer machine with 3, 3′-diaminobenzidine as the color indicator following by the addition of a light Hematoxylin stain. Amygdala and hippocampal stained slides were examined by a neuropathologist (DWD) and neuroscientist (KAJ) to assess for the presence of TDP-43 immunoreactive inclusions and for TDP-43 type [19]. We focused on these two regions because we and others have previous shown that these two regions are affected early by TDP-43 in PART [9, 10]. Cases were reviewed at a magnification of 200X and were considered positive if any TDP-43 immunoreactive inclusion including neuronal cytoplasmic inclusions, dystrophic neurites, neuronal intranuclear inclusions, fine neurites of the CA-1 region of the hippocampus, NFT-associated TDP-43 (TATs) [20], perivascular and granular inclusions were observed [2, 17]. If no TDP-43 was observed in either the amygdala or hippocampus, the case was designated TDP-negative. If TATs were the predominant lesion type or very prominent in the amygdala the case was labelled as TDP-43 type-beta [19]. However, if TATs were absent or relatively scant compared to the presence of neuronal cytoplasmic inclusions and/or dystrophic neurites, the case was labelled as TDP-43 type-alpha[19]. TDP-43 stage was also determined as previously described on a scale from stage 1 (amygdala only) –stage 6 (extending to frontal cortex or basal ganglia) [9, 17].
Brain MRI segmentation
In this study, we used the MRI scan nearest to the time of death in participants with multiple serial MRIs. All participants underwent a volumetric MRI using a standardized protocol [21], and all scans underwent correction for intensity inhomogeneity and were subjected to quality control for subsequent analysis. The segmentation of hippocampal subfields was performed using FreeSurfer version 7.4.1, which utilizes a probabilistic atlas constructed from ultra-high-resolution ex vivo MRI data (approximately 0.1 mm isotropic) to produce an automated segmentation of the hippocampal substructures [22, 23]. FreeSurfer subdivides hippocampal substructures into head, body, and tail, where applicable, and generates an additional set of segmentations of each hemisphere at different levels of hierarchy. This approach echoes historical methodologies, as delineated by an early foundational paper by Clifford et al., which pioneered the analysis of the hippocampus by its segmented region [24]. An example segmentation is shown in Figs. 1and 2. Averaging the volumetric data of the left and right hippocampi has been utilized to reduce lateralization bias, to increase statistical power by reducing variability due to side-specific differences, and for a holistic representation of hippocampal volume changes. To adjust for differences in head size, total intracranial volume (TIV) was calculated using the standard FreeSurfer processing pipeline.
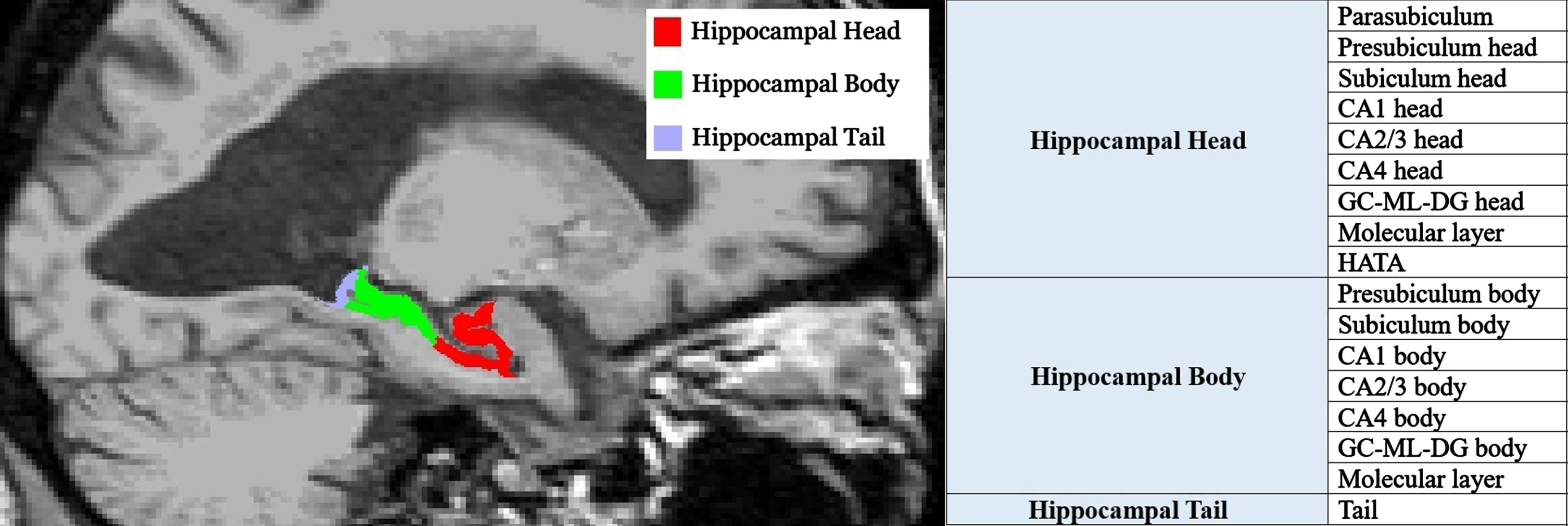
Hippocampal subfields’ sagittal view. GC-ML-DG, Granule Cell and Molecular Layers of the Dentate Gyrus; HATA, Hippocampal-Amygdala Transition Area.
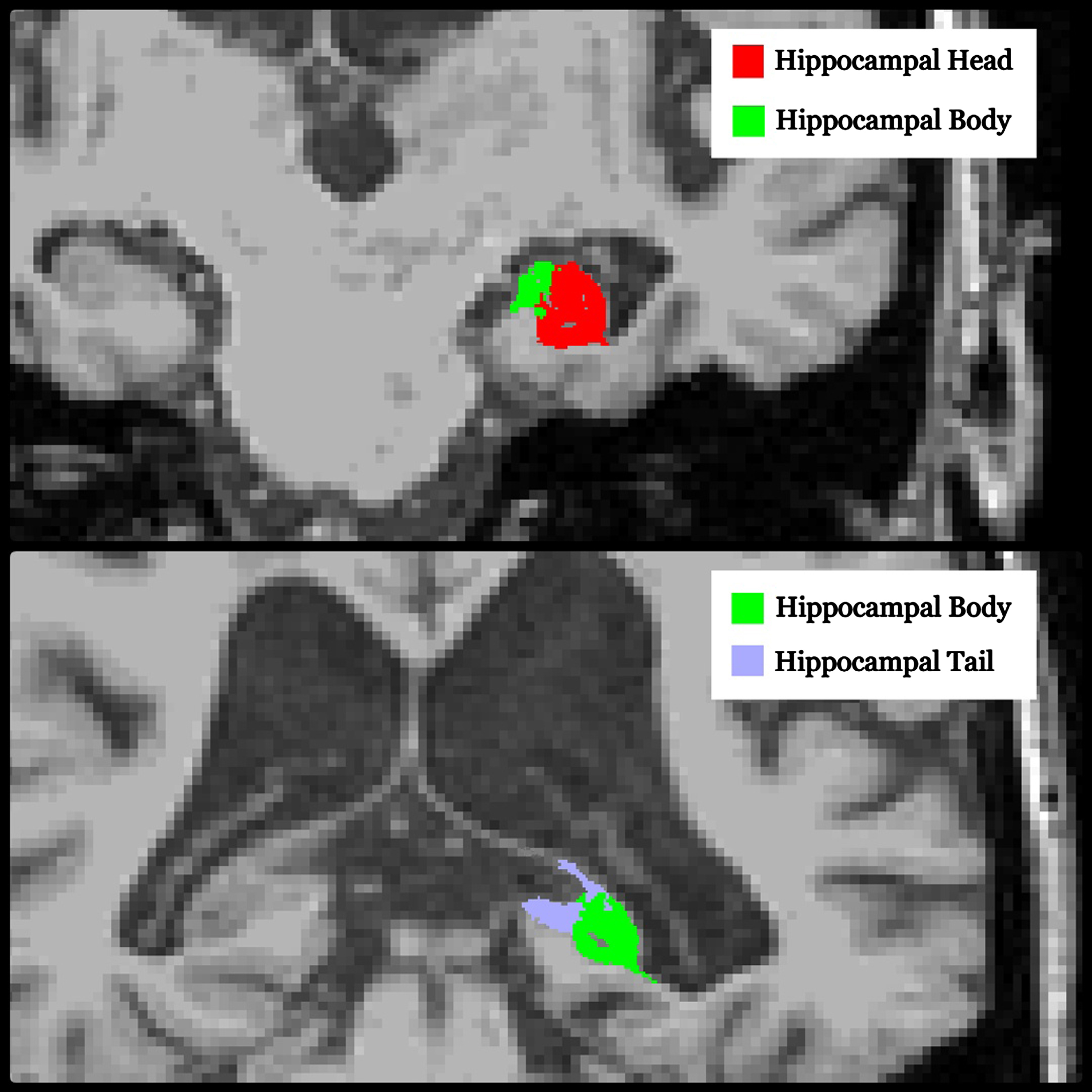
Hippocampal subfields’ coronal view.
Statistical analysis
Statistical analysis was performed using STAT/BE 18, with significance set at a threshold of p≤0.05. Demographics, clinical characteristics, and MRI volumes of the patients were analyzed using basic descriptive statistics, including frequency, percentage, mean, and standard deviation (SD). Differences between TDP-43 positive (+) and negative (–) cases were assessed by comparing the means and SDs of the two groups. Logistic regression was utilized to calculate the odds ratio for the association between TDP-43 positivity (dependent) and hippocampal subfield volumes (independent), adjusting for TIV (independent). The Pearson correlation coefficient and one-way analysis of variance (ANOVA) were used to assess for correlations between TDP-43 stages and volumes. The Benjamini-Hochberg procedure was employed to control for a false discovery rate from performing multiple comparisons in our logistic regression analyses.
RESULTS
Demographic characteristics are displayed in Table 1. Of the 115 participants, approximately 43% were female. The average age at the time of their last MRI was 84 years, and the average age at death was 88 years. Thirty-seven participants were TDP-positive (32%). TDP-43 type was available for 15 participants in the TDP-43 positive group, and all were type-alpha. There was a significant difference observed between TDP-positive and negative groups in the frequency of hippocampal sclerosis (p = 0.001). Ten participants had hippocampal sclerosis (8.7%), with eight in the TDP-positive group (21.62%) and two in the TDP-negative group (2.56%). Both patients in the TDP-43 negative group had vascular lesions in the hippocampus. No significant differences were noted in the remaining demographic variables between TDP-positive and TDP-negative groups. There was a trend, however, for the TDP-43-positive group to be older at the time of death.
Participant characteristics
Data is shown as mean (±standard deviation) or N (%). AVLT, Auditory Verbal Learning Test; CDR-SB, Clinical Dementia Rating Scale –Sum of Boxes; TIV, Total Intracranial Volume; MMSE, Mini-Mental State Examination; WMS-R LM1, Wechsler Memory Scale-Revised Logical Memory I, NFT, neurofibrillary tangle.
The findings for hippocampal volumes are shown in Table 2. Whole hippocampal volume was significantly smaller in the TDP-positive cases compared to the TDP-negative cases (p = 0.049). After stratification of the hippocampus into head, body, and tail, the TDP-positive cases showed smaller volumes of the hippocampal body (p = 0.031) and tail (p = 0.021), but not the hippocampal head (p = 0.129), compared to TDP-negative cases. When these regions were further subdivided into hippocampal subfields, the TDP-positive cases showed smaller volumes of molecular layer of hippocampal body (p = 0.046), CA1 body (p = 0.031), subiculum body (p = 0.023), as well as presubiculum body (p = 0.046) compared to TDP-negative cases. After separating definite from possible PART, no association was found between TDP-43 and volume in those with definite PART. TDP-43 staging was available for 32 participants of the TDP-43 positive group and there was no correlation found between staging and volumes of the whole hippocampi or subfields.
Hippocampal volume findings across TDP-43 status groups
Data is shown as mean (±standard deviation). Statistical analysis performed after adjusting for total intracranial volume. GC-ML-DG, Granule Cell and Molecular Layers of the Dentate Gyrus; HATA, Hippocampal-Amygdala Transition Area. Unitless normalized volumes are calculated as the ratio of hippocampal volumes to total intracranial volume, multiplied by 100,000.
Results from the logistic regression analysis, are presented in Table 3. After adjusting for TIV, we found that reduced volumes of the whole hippocampus, hippocampal body, and its constituents including the molecular layer, CA1, subiculum and presubiculum had higher odds of being TDP-43 positive. After adjusting for hippocampal sclerosis, the results were essentially unchanged. TDP-43 remained associated with hippocampal body (p = 0.046), hippocampal tail (p = 0.025), CA1 body (p = 0.041), and subiculum body (p = 0.036), and approached significance with whole hippocampus (p = 0.055).
Odds ratio of being TDP-43 positive
Statistical analysis performed with logistic regression adjusted for total intracranial volume. P-values were corrected for false discovery rates (FDR) by employing the Benjamini-Hochberg procedure.
DISCUSSION
In this cohort of 115 cases of PART, we assessed the association of TDP-43 status with hippocampal subfield atrophy. We found that TDP-positive cases exhibited smaller whole hippocampal volumes compared to TDP-negative cases, validating our previous results [12, 13]. Further stratification revealed that the body and tail of the hippocampus showed smaller volumes in those with TDP-43. By further dissecting the hippocampus into subfields, we were able to attribute volume loss to the molecular layer of hippocampal body, CA1 body, subiculum body, and presubiculum body.
Our finding of smaller whole hippocampal volume in TDP-positive cases with PART concurs with our previous studies demonstrating the influence of TDP-43 on brain volumes in PART [12, 13]. In this study, we further investigate the specific regions and subfields of the hippocampus and discovered that differences in volume loss are more prominent in the body and tail of the hippocampus in the TDP-positive cases. Moreover, with further stratification and analysis, volume loss was observed in the molecular layer of hippocampal body, CA1 body, subiculum body, as well as presubiculum body. To our knowledge, this is the first study to investigate the effects of TDP-43 in patients with PART on hippocampal subfields. Based on the FreeSurfer anatomical atlas [22], we note that the hippocampal tail has no subfield components. However, the body is comprised of various components, including the molecular layer, CA1, subiculum, and presubiculum, which are all significantly smaller in volume in TDP-positive cases. Notably, the same effect was not observed in absence of beta-amyloid which is consistent with our previous study that also found no association between TDP-43 and whole hippocampal volumes in patients with definite PART [5]. This is likely because the vast majority of patients with definite PART are cognitively normal with absent to minimal volume loss, and no range of data for volumes. We did observe a trend for the volume of the head of the hippocampus to be smaller in the TDP-positive cases compared to the TDP-43 negative cases.
In neuropathological studies where TDP-43 pathology has been observed, there is significant deposition of TDP-43 in regions and subregions of medial temporal lobe structures including the CA1 and subiculum regions of the hippocampus [25]. TDP-43 has been previously found to have an effect on the hippocampal subfields CA1 and subiculum in Alzheimer’s disease and has been associated with inward deformation of the hippocampus, which correlates with cognition scores [14]. It is not surprising that these two regions show volume loss associated with TDP-43 in Alzheimer’s disease and now in PART, because these two regions are directly affected by TDP-43 deposition [17]. In PART, however, it is vulnerability of the CA2 subregion due to NFT degeneration that is greatest [4, 26]. Our findings therefore are more keeping with volume loss due to TDP-43 than volume loss due to NFTs and tau.
We discovered that TDP-43 in PART predominantly impacts the hippocampal body, specifically affecting four main areas: the molecular layer, CA1, subiculum, and presubiculum. Given the observed volume loss in these areas, it may be worth exploring their potential as imaging biomarkers for TDP-43 associated atrophy in future longitudinal studies. However, the long-term performance and reliability of these biomarkers remain to be established through comprehensive research. Differential atrophy of the CA1 versus CA2 regions of the hippocampus could be utilized to assess treatment response to a tau versus TDP-43 targeted pharmacological molecule.
The CA1 and subiculum are also involved in hippocampal atrophy in Alzheimer’s disease where both Aβ and tau are present [27–29]. In individuals with Alzheimer’s disease, reduced volumes of both CA1 and subiculum have been observed, consistent with atrophy in these two subfields [30]. Given that we have demonstrated a similar pattern of involvement in PART that is related to TDP-43, one could hypothesize that it is TDP-43 that is possibly driving atrophy in these two subfields in Alzheimer’s disease. It is unclear however, whether there are differences between TDP-43 related subfield volume loss in Alzheimer’s disease and TDP-43 related subfield volume loss in PART. For example, our analysis suggests more specific involvement of these regions in the body of the hippocampus as opposed to the head. It is possible that the hippocampal head is more affected than the body in Alzheimer’s disease.
Unlike the CA1 and subiculum which have been discussed in previous studies, less is known about the molecular layer. In one study comparing the volume of the molecular layer in cognitively intact versus mild cognitively impaired patients, no significant difference was observed [31]. Similarly, little has been reported on the presubiculum in the body of the hippocampus.
There is a strong association between TDP-43 pathology and hippocampal sclerosis, with hippocampal sclerosis conceptualized as a downstream consequence of the presence of TDP-43 [32]. Hence, it is not surprising that those with TDP-43 were more likely to have hippocampal sclerosis compared to those without TDP-43. Unlike hippocampal sclerosis, there was no difference in the Braak NFT stage or the Thal phase between those with and without TDP-43. This is most likely due to the fact that our inclusion criteria limited cases to specific Braak and Thal stages. It is well known that patients with TDP-43 have higher Braak NFT stages when the full spectrum of Braak stage cases (I–VI) are included in the study [33, 34].
Strengths and limitations
One of the strengths of our study is the large sample size. In fact, to the best of our knowledge, this is the largest cohort to investigate the association between TDP-43 and hippocampal subfields in patients with PART. Another strength is that our cases were well characterized pathologically. Our study has certain limitations that warrant discussion. First, although segmenting the hippocampus into head, body, and tail provides more precision than analyzing the whole hippocampus, some subfield volumes are too small for accurate measurement using T1-weighted MRI. Consequently, the accuracy of some of our segmentations is likely poorer compared to larger parcellations. We also recognize that the hippocampal subfields lack distinct internal contrast in these roughly 1 mm clinical resolution T1-weighted MRI scans, resulting in the subfield segmentations being largely dependent on external atlas priors. In future studies, this limitation could be addressed by incorporating T2-weighted MRI [22]. Thirdly, the low odds ratios observed in our logistic regression model suggest that the differences in brain volume identified are small and may be influenced by other external factors. Fourth, we only identified one unaffected control case that was without PART and TDP-43 and had an antemortem GE MRI scan; therefore, we were unable to compare PART participants who were TDP-43 negative with unaffected control cases. Lastly, we were unable to assess difference by TDP-43 type as all cases were TDP-43 type-alpha. This is not surprising since TDP-43 type-beta is associated with high Braak NFT stages [19] and the cases in this study with PART were Braak NFT stages≤IV.
Conclusion
In conclusion, our study underscores the association between TDP-43 pathology and hippocampal subfields in PART. It highlights which subfields are contributing to whole hippocampal volume loss, and particularly implicate the body and tail regions and specifically volumetric loss to the subfields of the CA1 and subiculum in the body.
AUTHOR CONTRIBUTIONS
Hossam Youssef (Conceptualization; Data curation; Formal analysis; Methodology; Project administration; Visualization; Writing – original draft); Rodolfo G. Gatto (Validation; Writing – review & editing); Nha Trang Thu Pham (Methodology; Resources; Software); Ronald C. Petersen (Validation; Writing – review & editing); Mary M. Machulda (Validation; Writing – review & editing); R. Ross Reichard (Validation); Dennis W. Dickson (Validation); Clifford R Jack (Validation; Writing – review & editing); Jennifer L. Whitwell (Conceptualization; Supervision; Validation; Writing – review & editing); Keith A. Josephs (Conceptualization; Methodology; Supervision; Validation; Writing – review & editing).
Footnotes
ACKNOWLEDGMENTS
We would like to thank Ms. Monica Castanedes for performing the TDP-43 immunohistochemistry and Dr. Leonard Petrucelli for giving us the TDP-43 antibodies that were used in this study.
FUNDING
This study was funded by NIH grants R01 AG37491 (PI: Josephs), P50 AG16574 (PI: Petersen) and U01 AG006786 (PI: Petersen).
CONFLICT OF INTEREST
Jennifer Whitwell is an Editorial Board Member of this journal but was not involved in the peer-review process of this article nor had access to any information regarding its peer-review.
All other authors have no conflict of interest to report.
DATA AVAILABILITY
The data that support the findings of this study are available upon reasonable request to the corresponding authors (KAJ).