
Abstract
BACKGROUND:
Andean berry contains several classes of phenolic compounds which have showed antioxidant and cytotoxic activity.
OBJECTIVE:
To characterize the Andean berry aqueous extract and to study their anti-proliferative mechanisms on SW480 and SW620 cell lines (human colon adenocarcinoma).
METHODS:
Total phenolic and total anthocyanins contents were determined by Folin-Ciocalteau and differential pH methods, respectively. Antioxidant activity was measured by FRAP, ORAC and DPPH methods. Antiproliferative effect was determined by sulforhodamine colorimetric method and apoptosis was analyzed by flow cytometry using propidium iodide/Annexin-V. Mitochondrial potential was evaluated using DIOC6 and ROS levels were measured by 2,7-Dicholorodihydrofluorescein diacetate (DCFH-DA).
RESULTS:
The total phenol and anthocyanin content were 4409.78± 63,05 mg equivalents of gallic acid/100 mL and 106,57± 1.43 mg equivalents of cyanidin-3-glycoside/100 mL, respectively. Andean berry extract showed antioxidant activity by FRAP, ORAC and DPPH methods and antiproliferative effect on SW480 and SW620 cells. It was observed a cell cycle arrest at S and G2/M phases on SW480 and at G0/G1 phase on SW620 cells. Aqueous extract did not induce mitochondrial depolarization or affect intracellular ROS levels.
CONCLUSIONS:
Andean berry aqueous extract has antioxidant capacity and induces apoptosis involving cell cycle arrest in SW480 and SW620 cells without mitochondrial damage.
Introduction
Colon cancer (CC) is a heterogeneous disease, which can be sporadic or hereditary [1, 2]. According to the latest GLOBOCAN report, this type of cancer gave rise to 1.8 million cases worldwide and remains one of the leading causes of cancer deaths [3].
Most of colon cancers arise sporadically (60–65%) due to the progressive accumulation of genetic and epigenetic alterations, subsequently leading to malignant transformation [2]. Approximately 80 to 85%of cases follow the adenoma-carcinoma sequence, which can be divided into four stages: initiation, progression, promotion, and metastasis [2]. During initiation, the normal cell is transformed into an initiated cell after exposure to an initiating factor; later, a small adenoma (promotion) forms, which becomes a large adenoma over time. Finally, during the progression stage, transformation to carcinoma occurs, and later metastasis may occur [2]. Metastasis to regional and distant nodules occurs in 36%and 20%of cases, respectively, and it is one of the leading causes of cancer-related deaths [4, 5].
Among the factors that have been associated with an increased risk of CC are: a high consumption of red meat, processed meats and alcohol, as well as sedentary lifestyle, and inflammatory bowel diseases [6–8]. Furthermore, it has been suggested that the consumption of fruits and vegetables can help to prevent CC [9], for which numerous studies have been carried out with different fruits including berries to evaluate their antitumor potential [10].
Currently, berries are recognized as the fruits that have one of the most positive health impacts, and therefore are included within the group of functional foods [11]. These fruits are rich in minerals, vitamins, fiber, organic acids and also phenolic compounds mainly from the group of flavonoids, such as anthocyanins, flavanols, flavonols, hydrolyzable tannins, ellagitannins and phenolic acids [10–12].
In vitro and in vivo studies have shown the chemopreventive activity of berries, through different molecular mechanisms that include: protection of DNA against oxidative damage caused by reactive oxygen species (ROS), induction of apoptosis, cell cycle arrest at the G2/M phase, inhibition of pro-inflammatory cytokine production, inhibition of angiogenesis and metastasis [11, 13].
The berries belong to several families, and among the most studied are Vitaceae, Rosaceae and Ericaceae [14]; the latter family includes the genus Vaccinium which comprises more than 400 species that are distributed in the northern hemisphere, the Andean region of Colombia, South Africa and Madagascar [14]. The fruit of Vaccinium meridionale Swartz is also known as Andean Berry [14] and it has demonstrated capacity to neutralize ROS [14]. Andean berry juice induce cytotoxic and antiproliferative effects on human colon adenocarcinoma SW480 cell line and its derived metastatic SW620 cell line [14]. The juice of Andean Berry has also shown antioxidant capacity in vivo, evidenced by the improvement of the antioxidant capacity in plasma of healthy individuals who consumed the juice for 14 days [15].
Thus, in order to get more insight into the mechanisms involved in the cell death induced by aqueous extract of Andean Berry, it was evaluated the apoptotic and cytotoxic effect on SW480 and SW620 cell lines. These two cell lines have been validated as an in vitro model of colon cancer progression from a primary tumor to metastatic disease [16].
Materials and methods
Plant material
Fresh Andean Berry (V. meridionale Swartz.) fruits were collected in June 2019, at the vicinity of El Retiro (Antioquia, Colombia) at 2175 m above sea level (6°8’ 6” N; 75°25’ 3” W) and an average temperature of 16°C. The material was registered with the voucher number ILS14050070. Ripe berries (black-violet color) selected, washed, disinfected with sodium hypochlorite (100 ppm) for 2 min at room temperature according to the Guide for the cleaning and disinfection of hands and surfaces END-147 by ICONTEC [17], washed several times with water and ground in a commercial blender at 2500 rpm for 2 min. The 50%of this pulp was stored at –10°C± 2°C, the other 50%of pulp was freeze-dried at 4.27± 0.5 mm H, –50°C, and the powder was stored at room temperature (25± 2°C) in polyethylene packages protected from light until the preparation of the aqueous extract.
Preparation of the aqueous extract of Andean Berry
The aqueous extract was prepared as described by Franco Tobón et al (2016) [18] with some modifications. In brief, the freeze-dried powder was reconstituted in distilled water at 12%, sonicated (42 KHz 135 W, 3 hours) in a sonicator (Branson B3510, Ultrasonic Corporation, Boston, MA, US), filtered with 0.22μm membrane, and stored in darkness at –20°C until use. The final concentrations of the extract used to treat the cells were 24 mg/mL (40%), 18 mg/mL (30%), 12 mg/mL (20%) and 6 mg/mL (10%).
Determination of total phenolic content
Total phenolic content was measured using Folin-Ciocalteau method [18]. Extract sample were mixed with Folin-Ciocalteau reagent (1:6 v/v dilution), Na2CO3 was then added to the solution (7.5% m/v) and incubated for 2 h. Finally, optical density (OD) was measured at 630 nm in a microplate reader (Biorad iMARK, Berkeley, CA, US). A calibration curve was constructed (0 –100μg/mL) using Gallic Acid (GA) as the standard, and results were presented as milligrams of Gallic Acid Equivalents per 100 mL extract or 100 g of fresh weight (pulp).
Determination of total anthocyanin content
The total anthocyanins content was determined by the differential pH method [19]. Optical density was measured at 530 nm and 700 nm in pH 1.0 and 4.5 buffers; using the equation A = [(A530 - A700) pH1.0 - (A530- A700) pH4.5], and cyanidin-3-glucoside with a molar extinction coefficient of 26,900 nm in a microplate reader (Biorad iMARK, Berkeley, CA, US). A calibration curve was constructed (0 –20μg/mL) using Cyanidin-3-Glucoside (Cy3G) as the standard, and results were presented as milligram Cy3G Equivalents per 100 mL extract or 100 g of fresh weight (pulp).
Determination of antioxidant capacity
Antioxidant capacity of aqueous extract was analyzed by different methods: FRAP (Ferric Ion Reducing Antioxidant Power), ORAC (Oxygen Radical Absorbance Capacity), and DPPH (2, 2-diphenyl-1-picrylhydrazyl).
The FRAP assay was used to evaluate the antioxidant activity of sample according to its ability to reduce the ferric iron (Fe+3) present in a complex with the 2,4,6-tri(2-pyridyl)-s-triazine (TPTZ) to the ferrous form (Fe+2), which had a maximum absorbance at a wavelength between 590–595 nm in a microplate reader (Biorad iMARK, Berkeley, CA, US) [20]. This assay was performed on a sodium acetic acid-sodium acetate buffer (pH 3.4), which contained TPTZ and FeCl3. A total of 900 mL of this solution, 50 mL of sample and 50 mL of distilled water were used. The absorbance reading of the blank without chromophore was taken into account for each sample. Calibration curve (0 –900μM) was prepared using Trolox as a standard. Results were expressed as μmol Trolox Equivalents/100 mL extract or 100 g of fresh weight (pulp) (TEAC± SEM).
The ORAC value was determined by the following methodology, 3 mL were prepared from the following solution: 21μL of a 10μM solution of fluorescein, 2899μL of 75 mM phosphate buffer (pH 7.4), 50μL of 600 mM AAPH (2,2’-azobis-2-methyl-propanimidamide, dihydrochloride) and 30μL of extract [21]. Fluorescence was recorded on a Perkin Elmer LS45 spectrofluorometer with a thermostatic multicell. The ORAC value was calculated by a calibration curve using different concentrations of Trolox (0 –100μM). Results were expressed as μmol equivalents Trolox/100 mL extract or 100 g of fresh weight (pulp) (TEAC± SEM).
The free radical DPPH assay was done following Rojano et al. [20]. Aliquots of 20μL of the extract and 280μL of DPPH/methanol were incubated for 30 min in the dark. Optical Density was measured at 490 nm in a microplate reader (Biorad iMARK, Berkeley, CA, US). A calibration curve was constructed (0 –30μM) using Trolox as a standard. Results were expressed as μmol Trolox Equivalents/100 mL extract or 100 g of fresh weight (pulp) (TEAC± SEM).
Cell culture
The cell lines used, SW480 and SW620 were obtained from the European Collection of Animal Cell Culture (ECACC, Salisbury, UK). They were cultured according to the procedure described by Maldonado-Celis et al, 2009 [22]. Briefly, the cells were cultured in 75 cm2 Falcon vials in Dulbecco’s modified Eagle’s (DMEM) medium supplemented with 25 mM glucose, 2 mM L-glutamine, 10%horse serum inactivated at 56°C, 100 U/mL penicillin, 100μg/mL streptomycin, and 1%non-essential amino acids (Invitrogen, Cergy-Pontoise, France). Incubations were carried out at 37°C in a humidified atmosphere with 5%CO2. The supplemented DMEM was replaced every 48 h. For all experiments, horse serum was used at a concentration corresponding to 3%, and the medium was supplemented with 10μg/mL insulin, 5μg/mL transferrin, and 5 ng/mL selenium (ITS; Invitrogen).
Cell viability assay
The cell viability was determined using Sulforhodamine B (SRB) colorimetric assay. This assay relies on the ability of SRB to bind to protein components of cells that have been fixed to tissue culture plates [23]. Briefly, 20.000 cells per well were seeded in a 96-well plate. After incubation for 24 h, the cells were treated for 24 h with different concentrations of extract (10%, 20%, 30%, 40%). Cells were fixed for 1 h after treatment with 100μL at 50%(v/v) trichloroacetic acid (TCA) at 4°C. The plate was washed with water to remove the TCA and it was let to get dried overnight; the cells were stained for 30 min with 100μL SRB solution (0.4%). The unbound dye was rinsed with 1%v/v acetic acid and then the plate was let to get dried overnight. Finally, 200μL 10 mM Tris solution (pH 10.5) was added to every well and the plate was incubated while agitated for 30 min at 55 rpm. The OD value was measured at 490 in a microplate reader (Biorad iMARK, Berkeley, CA, US). Percentage of inhibition of viability was calculated as follow = [1 - (ODt / ODc)]×100, where, ODt is optical density of treated cells and ODc is optical density of control (untreated cells). A linear regression on these data was used to determine the concentration at which the extract exerts half of its maximal inhibitory effect (IC50).
Cell growth rate
The effect on growth rate of aqueous extract in colon cancer cell lines was determined by using the SRB assay as described earlier with modifications. In brief, 3000 viable cells from each cell line were exposed to different concentrations of extract 24 h after seeding and incubated for different times. Control cells were untreated. Culture media were replaced every 48 h. Cell culture was stopped by the addition of trichloroacetic acid (50%v/v), and cell proteins were determined by staining with 200 mL SRB 0.4%w/v (Sigma-Aldrich, San Louis, Missouri, USA). The absorbance of SRB is proportional to the number of adherent and live cells were determined by measuring the optical density (OD) at 490 nm using a microplate ELISA reader (iMark Biorad, California, USA). All experiments were performed in triplicate.
Cell cycle analysis
The cell cycle distribution of cells was analyzed by using Propidium Iodide (PI) according to Nicoletti et al. (1991) method [24]. Briefly, 1×106 cells were maintained at 37°C and CO2 5%incubation, after 24 h the cells were treated at 30%; after 48 h cells were collected by trypsinization (0.5%trypsin/2.6 mM ethylenediaminetetra acetic acid tetrasodium salt), later they were fixed, washed and resuspended in PBS with PI and RNAsa for 30 min, at 37°C. The fluorescence of at least 10,000 events was analyzed in flow cytometer.
Apoptosis analysis
The cells SW480 and SW620 were exposed to the extract for 24 h and then the cells were collected by trypsinization and washed twice with PBS (phosphate buffered saline). Apoptosis was quantified by measurement of phosphatidylserine externalization by flow cytometry. The assays were carried out with the commercial annexin-V-FLUOS staining kit (Sigma-Aldrich), following the manufacturers’ instructions. The cell pellet was resuspended and incubated in solution containing the staining buffer, annexin-V-FITC and IP for 20 min at room temperature. Double stained cells were collected to analyze 10,000 events per sample by flow cytometer (FACSCanto™ II, BD Biosciences, Franklin Lakes, NJ, US) at 488 nm excitation, 515 nm for annexin-V and 617 nm for IP, in cell populations from which cell debris were excluded.
Mitochondrial membrane potential
SW480 and SW620 cells were analyzed as described by García-Gutiérrez et al (2017) [25]. In brief, cells were seeded at a final density of 1×106 cells /well in 6-well tissue culture plates and allowed to grow for 24 h. Then, they were treated with respective IC50 of Andean Berry extract at 37°C in 5%CO2 for 24 h. At the end of treatment, cells were collected using trypsin as described before and washed; then, cells were stained and incubated with 50μM DiOC6 (3,3-dihexyloxacarbocyanine iodide) (Thermo Fisher Scientific, Waltham, MA, USA) in darkness at 37°C for 15 min, washed twice with 2 mL of PBS, and then stained at room temperature in the dark with 10μg/mL propidium iodide (PI) for 10 min. Double stained cells were collected to analyze 10,000 events per sample by flow cytometer (FACSCanto™ II, BD Biosciences, Franklin Lakes, NJ, US) at 488 nm excitation and detection of emission with green filters (530/515 nm). This method allowed us quantifying cells with depolarized mitochondrial membrane.
Detection of intracellular reactive oxygen species
The 2,7-Dicholorodihydrofluorescein diacetate (DCFH-DA) was used as cell-permeable probe for detect the intracellular production of ROS [22]. This probe is metabolized inside mitochondrial subparticles, where it is deacetylated and oxidized to form the fluorescent compound DCF (Dicholorofluorescein), due to deacetylase enzyme activation in response to the increase of intracellular ROS. Thus 10.000 cells per well were seeded on a 96-well plate using maintenance medium. After 24 h without treatment, the cells were exposed to aqueous extract of Andean Berry for 24 h at 37°C and 5%CO2. Medium was rinsed off and a solution of 100μL of 8μM DCFH-DA with PBS was added for 60 min. Fluorescence was measured at 520 nm excitation/485 nm in a microplate reader (GlomaxTM Promega). Data are shown as the mean of relative fluorescence units (RFU) with standard deviation (RFU±SEM).
Statistical analysis
The data are presented as mean± standard error of mean (SEM) from three independent experiments. Linear regression analysis was used to calculate IC50. Statistical differences between treated and untreated cells (control) were evaluated by one-way Anova and specific differences were identified using Tukeys’ test. Graph Pad Prism V8.0 statistical software was used. For all cases a p < 0.05 was considered significant.
Results
Chemical characterization and antioxidant capacity of aqueous Andean Berry extract
The total contents of phenols, anthocyanins and FRAP, ORAC and DPPH values corresponding to antioxidant methods used are presented in Table 1. The R2 of the curves constructed with cyanidin-3-glycoside and gallic acid were 0.99 and 0.98, respectively; the R2 values for the calibration curves constructed with gallic acid for FRAP, ORAC and DPPH methods were 0.99, 0.99 and 0.93, respectively. In the aqueous extract, the anthocyanin content corresponds to 2%of the total phenolic compounds present in the extract. In relation to the antioxidant activity, the ORAC value showed that the extract contains compounds capable of better neutralizing reactive oxygen species, followed by the FRAP value that shows a lower iron reducing capacity and finally the DPPH value indicated the low capacity of the compounds present in the aqueous extract to interact with the DPPH molecule (Table 1).
Phytochemicals and antioxidant activity of Andean Berry extract
Phytochemicals and antioxidant activity of Andean Berry extract
The results are expressed as the mean±SD of three independent experiments, in triplicates. 1mg Gallic Acid Equivalents/100 mL; 2mg Gallic Acid Equivalents/100 g Fresh Weight; 3mg Cyanidin-3-Glycoside Equivalents/100 mL; 4mg Cyanidin-3-Glycoside Equivalents/100 g Fresh Weight; 5 μmoles Trolox Equivalents/100 mL; 6 μmoles Trolox Equivalents/100 g Fresh Weight.
Cytotoxic activity was evaluated on SW480 and SW620 cells by Sulforhodamine B staining after 24 h of treatment with different concentrations of Andean Berry extract. The extract induced a decrease in cell viability of SW480 and SW620 cells in a dose-dependent manner and significant differences were observed between cell viability in all concentrations evaluated compared to control (Fig. 1). The highest reduction on cell viability on both cell lines was observed at 40%concentration of the extract. The IC50 of the extract was lower against SW480 cells compared to SW620 cells, corresponding to 32 and 38.4 %v/v, respectively (Fig. 2).
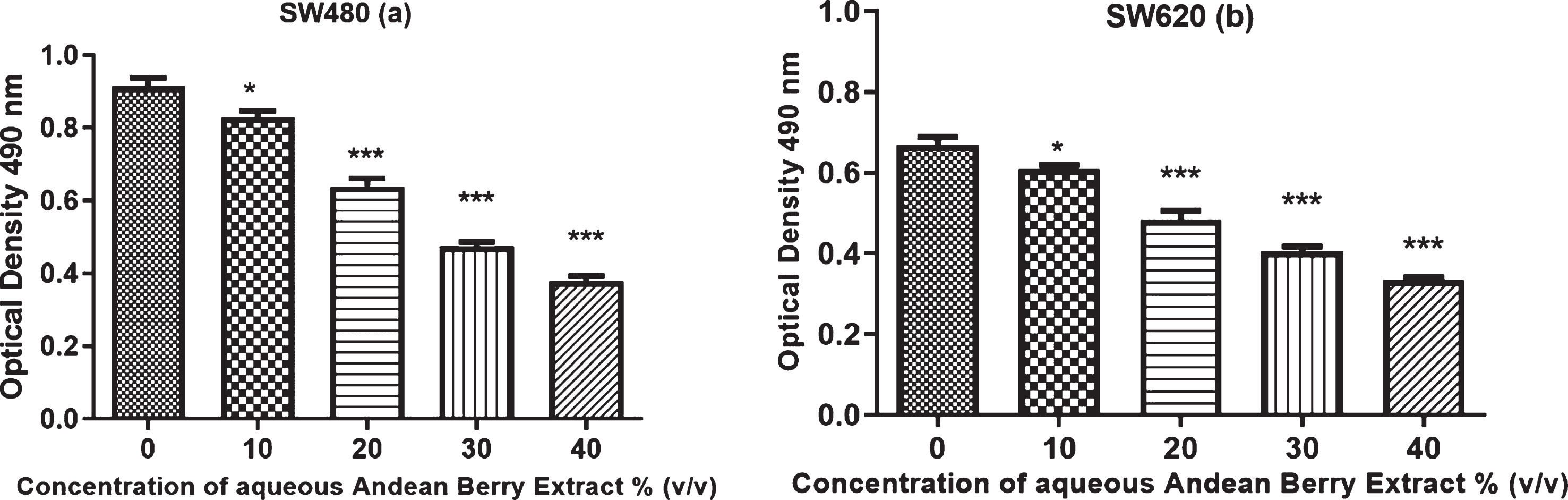
Effect of aqueous Andean Berry extract on cell viability of SW480 (a) and its metastatic derived cells SW620 (b). The cells were exposed to different concentrations for 24 h. Data expressed as mean± standard error from three independent experiments. (a) SW480: *p = 0.028, *** p < 0.0001; (b) SW620: *p = 0.041, ***p < 0.0001.

Calculation of IC50 of aqueous Andean Berry extract for SW480 (a) and SW620 (b) after treatment at different concentrations for 24 h. Data expressed as the average of three independent experiments.
The effect of the extract at several concentrations on SW480 and SW620 proliferation rate was measured at different times. The highest reduction in the proliferation rate on both cell lines was observed after 72 h of treatment. The proliferation rate of SW480 cells decreased from 24 h to 72 h after the treatment with the extract at 30 %and 40 %concentration. In contrast, Andean Berry extract inhibited SW620 proliferation only at the concentration of 40%after the different times of treatment (Fig. 3).
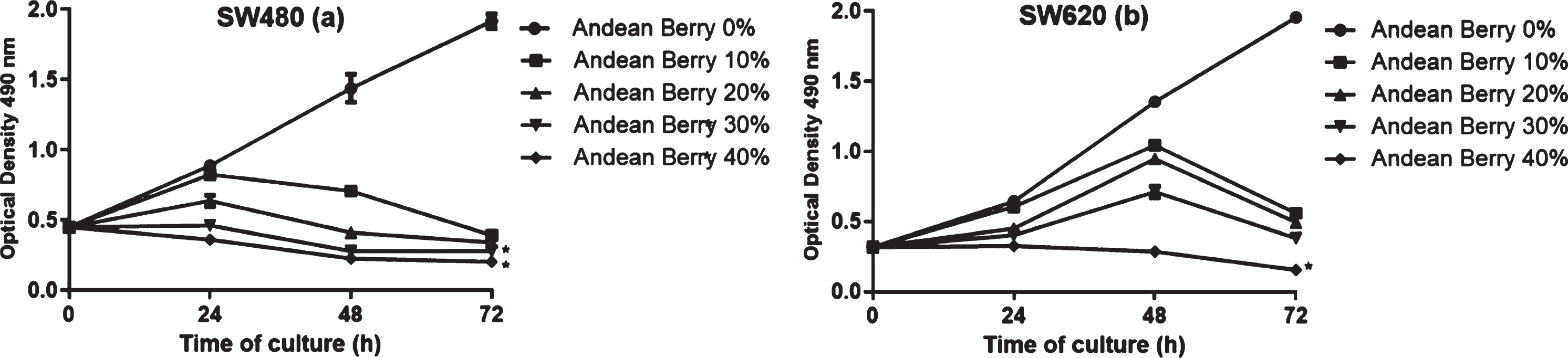
Antiproliferative effect on cells SW480 (a) and SW620 (b) of the aqueous Andean berry extract. Data expressed as mean± standard error from three independent experiments. * Significant difference between Andean Berry treatments compared to the control (p < 0.05). (a) SW480: *p = 0.047, *p = 0.038; (b) SW620: *p = 0.034.
Considering berries have shown to induce cell cycle arrest on several cancer cell lines, the effect of Andean Berry extract on cell cycle distribution was evaluated. The extract induced an arrest of SW480 cells at S (30.2 %) and G2/M (23.8 %) phases and decreased the percentage of cells at G0/G1 (32.9 %) and SubG0/G1 (0.7 %) compared to control which showed 2.6 %, 54.3 %, 22.4 %and 16.2 %at SubG0/G1, G0/G1, S and M phases. In contrast, SW620 cells showed an increase at G0/G1 phases (31 %) and a decrease at G2/M phase (13.7 %) after extract treatment (30%). Also, the extract induced and increase in the percentage of SW620 cells at SubG0/G1 phase (37.01 %) which could indicate apoptosis (Fig. 4).
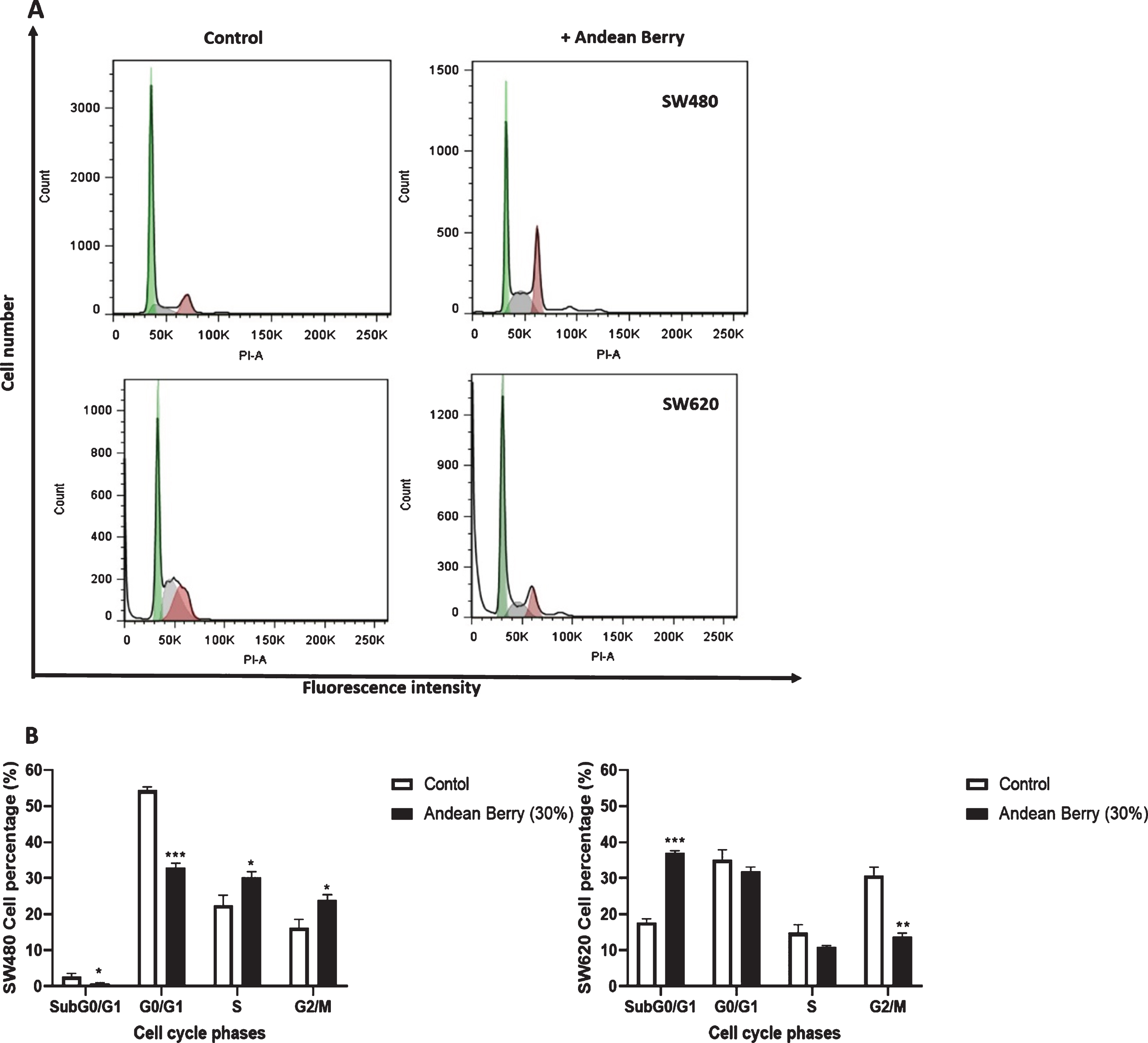
Effects of aqueous Andean Berry extract (30%) on SW480 and SW620 cell cycle. (A) Flow cytometry analysis was done using propidium iodide after 24 h of treatment. (B) SW480 and SW620 cells percentage distributed in cell cycle phases. Data are presented as the mean value± SEM of at least three separate experiments. Andean Berry treated cells versus controls:
In order to evaluate if Andean Berry extract (IC50: 30 %v/v) induce apoptosis, it was evaluated the exposure of phosphatidylserine as a marker of early apoptosis using annexin V and IP to identify cells in late apoptosis or necrotic cells (Table 2). Figure 5 corresponds to representative dot plots of the effect of Andean Berry aqueous extract on SW480 and SW620 cells after double staining with Annexin-V/PI. This double staining is useful to discriminate non-apoptotic/necrotic cells (Q1: Annexin-V/PI+); late apoptosis (Q2: Annexin-V+/PI); early apoptosis (Q3: Annexin-V+/PI); and viable cells (Q4: Annexin-V/PI). SW480 and SW620 cells increase the percentage of cells positive for early and late apoptosis after the treatment with the extract during 24 h compared to the respective untreated cells, being SW480 cell line more sensitive than SW620 cells that showed more apoptotic cells after treatment in the same conditions. Although, the extract induced a significant increase of necrotic SW480 and SW620 cells compared to control group, the percentage of cells in apoptosis were higher. The percentage of viable cells decreased after the treatment with the extract which is in agreement with the results of the assay of cytotoxicity and antiproliferation.
Effect of Andean Berry extract on SW480 and SW620 cells cell cycle distribution
Effect of Andean Berry extract on SW480 and SW620 cells cell cycle distribution
Data in the table are the mean value± SEM of at least three separate experiments after 24 h of treatment. Q: Quadrant. Andean Berry treated cells versus controls:
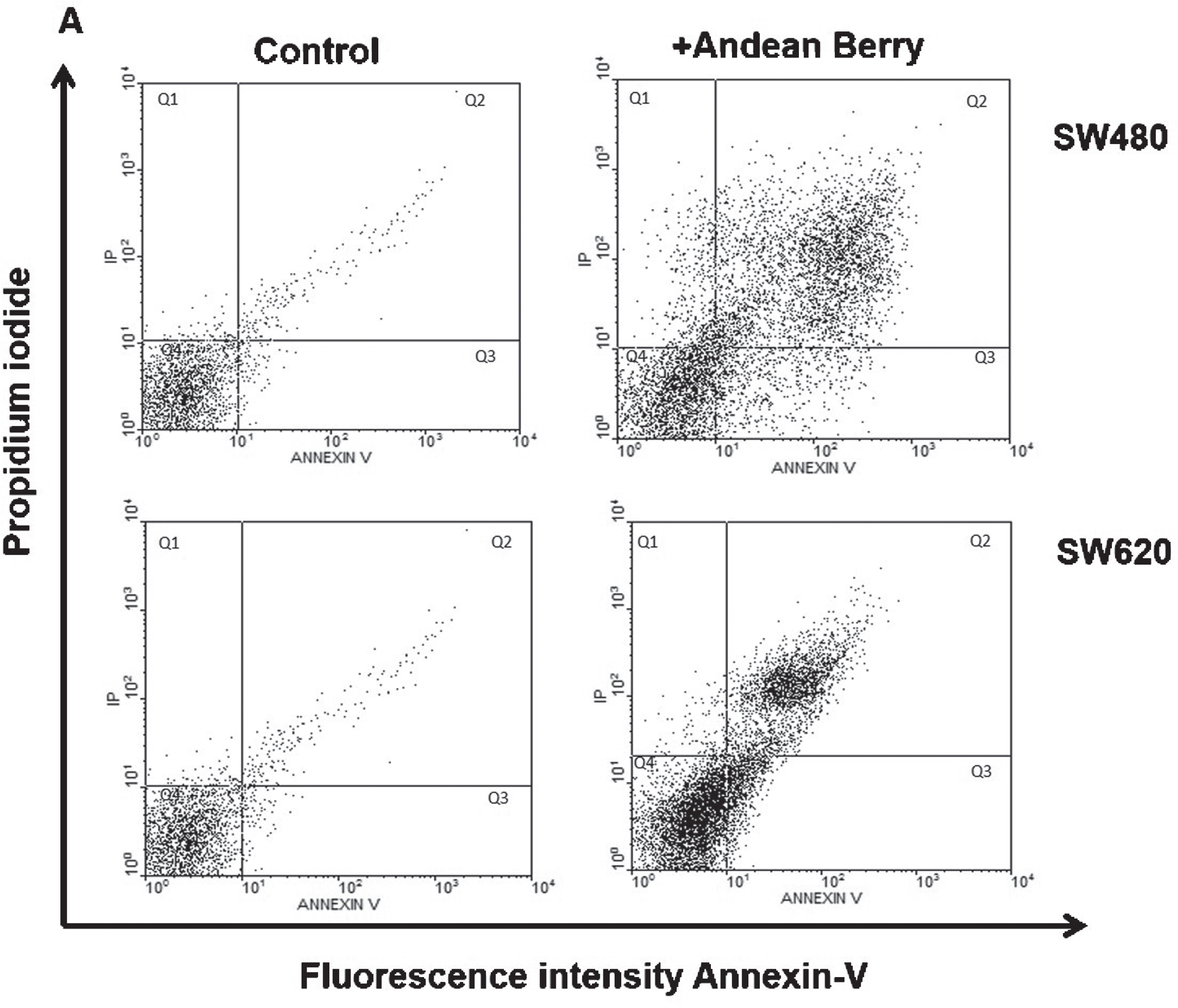
Cytometric analyses of apoptotic and necrotic cells determined using Annexin-V and PI staining after Andean Berry extract exposure. Representative dot plots of showing SW480 and SW620 cells untreated (control) or after 24 h treatment with 30%aqueous Andean Berry extract.
In addition, we evaluate the mitochondrial potential of the cells and integrity of plasma membrane integrity using DiOC6 and PI, respectively (Table 3). Figure 6 correspond to representative dot plots of the effect of Andean Berry aqueous extract (IC50: 30 %v/v) on SW480 and SW620 cells after double staining with DIOC6/PI. This double staining is useful to discriminate dying cells, non-apoptotic/necrotic cells, low ΔΨ m, and low membrane integrity (Q1: DiOC6–/PI+); late apoptosis, high ΔΨ m, and low membrane integrity (Q2: DiOC6+/PI+); high ΔΨ m and good membrane integrity (Q3: DiOC6+/PI–); and early apoptosis, low ΔΨ m, and good membrane integrity (Q4: DiOC6–/PI–). The Andean Berry aqueous extract increased late apoptotic cells without mitochondrial depolarization, suggesting a non-mitochondrial apoptotic cell death like the extrinsic apoptosis.
Effect of Andean Berry extract on SW480 and SW620 cells mitochondrial potential
Data in the table are the mean value± SEM of at least three separate experiments after 24 h of treatment. Q: Quadrant. Andean Berry treated cells versus controls:

Cytometric analyses of mitochondrial transmembrane potential (DIOC6) and plasma membrane integrity (PI) after Andean Berry extract exposure. Representative dot plots of showing SW480 and SW620 cells untreated (control) or after 24 h treatment with 30%aqueous Andean Berry extract.
Taking into account that an increase in production of ROS can induce apoptosis, we evaluated the levels of intracellular ROS using CM-H2DCFDA which emits green fluorescence in the presence of ROS. The amount of ROS did not change after the treatment with the extract compared to the control on both cells, SW480 y SW620 (Fig. 7).
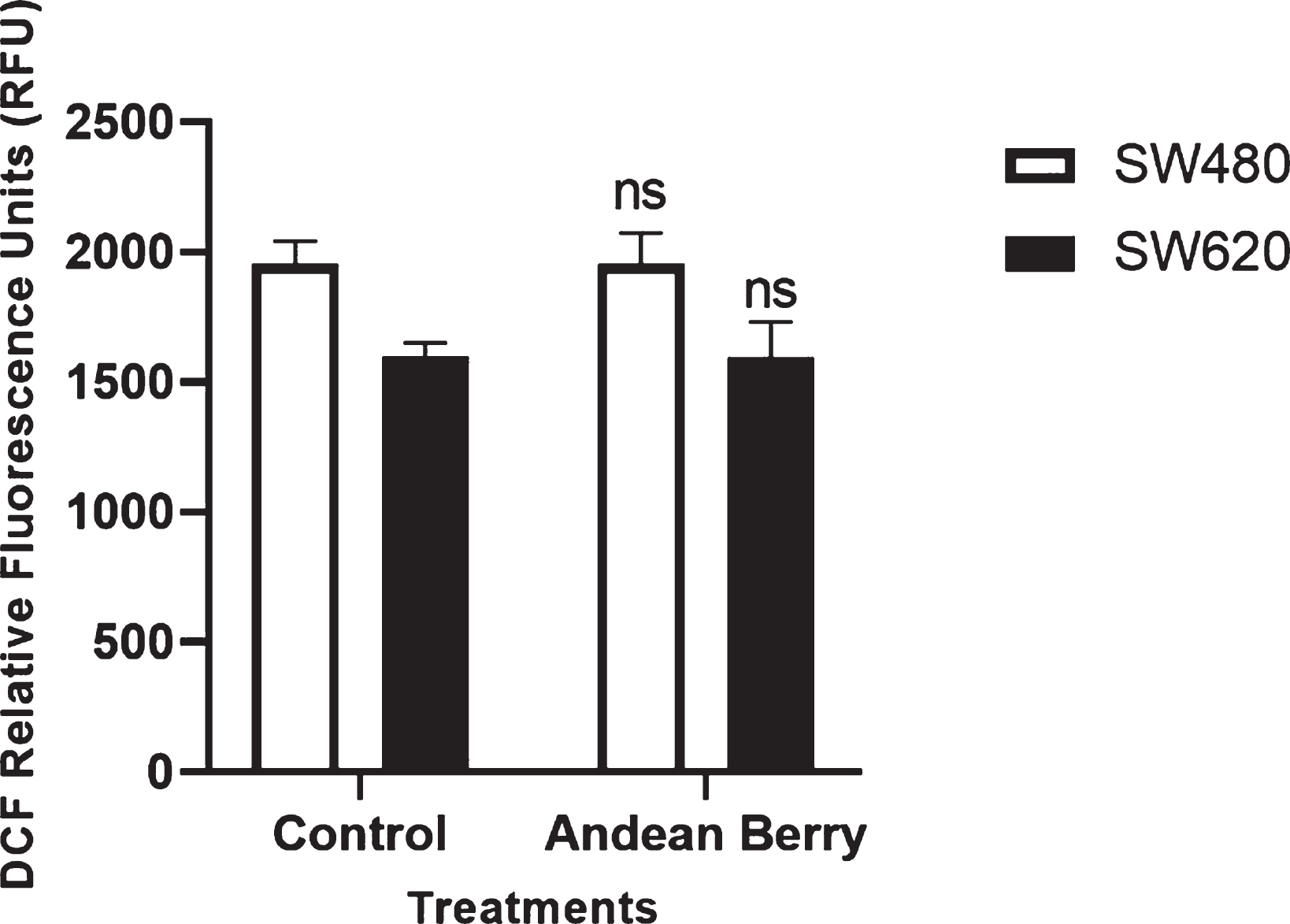
Intracellular reactive oxygen species production induced by Andean Berry in SW480 and SW620 cells exposed or not to extract for 24 h. The RFU values are presented as the mean value±SEM of at the least three separate experiments. ns: no significant (Andean Berry treatment versus controls.
Due to the side effects and toxicity generated by the agents currently available for the treatment of colon cancer, new therapeutic alternatives have been investigated including natural products from human diet among them fruits such as berries of the genus Vaccinium. Taking the above into account, the phenolic and total anthocyanin content, the antiproliferative and antioxidant capacity and the possible mechanism of induction of cell death of an aqueous extract of Andean Berry on human colon adenocarcinoma cells SW480 and their derived-metastatic SW620 cells were evaluated.
In our study, the aqueous extract of Andean Berry compared to the fresh fruit showed a significantly higher content of total phenols and anthocyanins, but compared with other studies using Vaccinium meridionale for Juice and nectar, the total phenolic content ranged 2.2 to 20.2-fold times [18, 28]. The total anthocyanins content in the extract was comparable to other studies for juice and nectar prepared with this fruit, which showed values between 0.418 –0.564 mg of cyanidin-3 glucoside equivalents /mL of sample [18, 27], respectively.
Regarding the total phenol content in the fruit (mg GAE/ 100 g FW) was comparable to that found in Northern Highbush blueberry (181–473) [21] and Vitis vinifera L. grape (151–246) [29], but lower than Rabbiteye blueberry (230–457) [21], Lowbush blueberry (290–495) [21], Andean blackberry (Rubus glaucus Benth) (266), and Andean blueberry (Vaccinium floribundum Kunth) (882–925) [30, 31]. The total anthocyanin content in Vaccinium meridionale berry (mg Cyanidin-3-Glycoside Equivalents/ 100 g FW) was also lower than those reported for berries of other species. In example, Prior et al. [21] found that the total anthocyanin content was for Northern Highbush blueberry (Vaccinium corymbosum) 92–235, Rabbiteye blueberry (Vaccinium ashei) 60–187, and Lowbush blueberry (Vaccinium angustifolium) 290–300.
These differences in the phenol and the anthocyanin content between the aqueous extract and fresh fruit can be explained based on the method to obtain and prepare the aqueous extract that led to the recovery of these bioactive compounds. The analysis of bioactive compounds and antioxidant capacity for fresh fruit was done from fruit pulp, a process that separates the pulp from the seeds and skin, but not as efficiently as ultrasound used here to obtain the aqueous extract, an oscillating sound pressure wave with a frequency over 20 kHz and non-thermal processing that facilitate extraction of phenolic compounds such as anthocyanins by increasing the mass transfer between the plant material and the solvent [32] such as water in our case, because anthocyanins are water-soluble flavonoids. Its use in anthocyanin extraction has already been demonstrated yielding 51.76± 3.70 mg anthocyanins/g extract of Hibiscus sabdariffa calyces [33] and 12.49 mg/g dry weight in 70%ethanol from blueberries (Vaccinium sp.) [34] and 4.27 mg C3G/g dry weight total anthocyanins and 16.41 mg GAE/g dry weight total phenols [35].
The aqueous extract of Andean Berry and the fruit showed the highest antioxidant capacity by ORAC, followed by FRAP and DPPH methods. If it is considered that in each method the reaction and conditions are different, these results suggest that in the extract and the fruit could be present several active compounds in a wide range of its metabolic profile, of different polarity, which means that there is a major source of metabolites with antioxidant potential that may be modified according to the extraction method such as phenolic acids, anthocyanins, and other (poly)phenols compounds [36].
The methods used here to measure the antioxidant capacity of the aqueous extract and the fruit were FRAP, ORAC and DPPH, which are used to evaluate the capacity of a compound to quench free radicals in organic and/or aqueous solutions.
Regarding the FRAP value, there are few reports that express it as μmol Trolox equivalent per sample for the genus Vaccinium; in some cases it has been presented as mg of ascorbic acid per sample, however, Alarcón-Barrera et al. [30] reported the FRAP and DPPH value for crude extracts of Andean blackberry (Rubus glaucus Benth) and Andean blueberry (Vaccinium floribundum Kunth) from Ecuador (FRAP: 100 –250μmol TE/g FW) and DPPH: 150 –250μmol TE/g FW). Our results agree with the FRAP values reported by these authors, but not the DPPH value, this can be attributed to the fact that the DPPH method evaluates the antioxidant capacity in organic media, and the evaluated fractions of the extract and fruit were aqueous and acidified methanolic extracts, respectively, with a high concentration of polar compounds that perform better in aqueous media, where the FRAP and ORAC assays were performed.
By other hand, the ORAC assay measures the capacity of an antioxidant to absorbs peroxyl free radical that is commonly found in the body and makes this reaction biological relevant (Prior et al., 1998). The ORAC value observed in this study was 17.3 fold times higher than the value reported for Andean Berry nectar [18], as well as the ORAC value of fresh fruit is comparatively higher than reported for different species and cultivars of Vaccinium (13.9 to 45.9μmol Trolox equivalents/g of fresh Berry) and is correlated to the anthocyanin (r2 = 0.77) or total phenolic (r2 = 92) content [21].
Anthocyanins as other phenolic compounds act as free radical scavengers against ROS because anthocyanins phenolic hydroxyl groups can quench reactive radical species by single electron transfer reaction and through hydrogen atom abstraction from phenolic grou, thus plant phenolic extract and matrices such as red wine and blueberry present important antioxidant activities attributed to the anthocyanin content, which suggests a high-impact biological and nutraceutical value of these compounds in plants and foods [37].
Concerning the antiproliferative activity of the Andean berry aqueous extract, it was effective for up to 72 h in SW480 and SW620 cells, resulting in IC50 values was 32%(19.2 mg/mL) and 38. 4%(23 mg/mL) at 48 h, values lower than those reported by Franco-Tobon et al. (2016) [18] with two nectar of Andean berries sweetened with sucrose (IC50 = 1.12 g/mL) and aspartame (IC50 = .4 g/mL) at 48 h of treatment in SW480 cells. In other studies, such as that of Seeram et al. (2006) [38] also observed that an aqueous cranberry extract inhibited 78%and 35%of HT29 and SW620 colon cancer cell growth, respectively after 48 h of treatment, but using a lower concentration (200μg/mL) than ours. These differences in antiproliferative ability against HT29 colon adenocarcinoma cells have been observed from higher to lower with anthocyanin-rich extracts of purple corn, chokeberry, bilberry, purple carrot, grape, radish and elderberry, respectively [39], this is attributed to the characteristics and content aglycones, sugars, and acylated acids, and the position and degree of glycosylation and acylation in the anthocyanin molecules considered the main factors influencing the anticancer property, however the structure-activity relationship of anthocyanins as chemoprotective agents remains to be further elucidated [40]. The major anthocyanin of Vaccinium meridionale Swartz berry is Cyanidin 3-galactoside, followed by Cyanidin 3-arabinoside, Delphinidin 3-hexoside, Delphinidin 3-pentoside, and Cyanidin 3-glucoside [41]. On the other hand, the product evaluated here can be considered as concentrated fruit juice, whose IC50 value is in the minimum range of fruit content for this type of beverage (7 to 25%) according to resolution 3929 of 2013 published by the Colombian Ministry of Health for the preparation and consumption of fruit-based beverages [42] suggesting that Andean Berry Juice at this concentration has a potential antiproliferative capacity against human colon cancer.
The effect of the Andean Berry extract on cell cycle distribution was also evaluated, and a cell cycle arrest was observed in the S and G2/M phases (SW480) and the G0/G1 phases (SW620). The Sub G0/G1 peak observed in SW620 cells after treatment with Andean berry extract represents stained cells with DNA reduced that was cleaved into smaller fragments during apoptosis. In other hand, SW480 cells did not appear the Sub G0/G1 peak and enter to apoptosis from the S phase by increasing in cell number which indicates that SW480 cells respond to a growth-promoting signal to duplicate DNA, but due to DNA-damage or replicative stress during exposure to the extract, SW480 cells get arrested in S-phase, in relation to cells in G2/M phases this mean that the Andean Berry extract interfere with the mitotic division. Shangguan et al. (2014) [43] found that the ginsenoside Rf extracted from the traditional Chinese herb ginseng, induced G2/M phase arrest in the human osteosarcoma cell line MG-63 cells by the downregulation of Cdk1 and cyclin B1. In addition, p53 activated is able to induce a G2 arrest that inhibits entry into mitosis, this event is activated when DNA synthesis is blocked or incompletely synthesized, may be p53 decreases cyclin B1 transcription and consequently its synthesis [44].
In order to confirm that the antiproliferative effect of the extract involve apoptotic events such as loss of membrane asymmetry by phosphatidylserine exposure and mitochondrial depolarization. We observed that the aqueous extract of Andean Berry was capable of inducing phosphatidylserine exposure on SW480 and SW620 cell membrane, indicating triggering apoptosis without mitochondrial damage because reduction in mitochondrial membrane potential did not change after exposition of SW480 and SW620 cells to the aqueous extract. Other berries of the genus Vaccinium have been shown to be able to induce apoptosis, including blueberry and black raspberry but require concentrations higher than those required by the Andean Berry extract. Black Raspberry extract induced internucleosomal degradation of HT-29 cells DNA at concentration of 200μg/mL after treatment during 48 h [45]. Also, blueberry hydroalcoholic extract at 0.474 mg/mL induced apoptosis on HT-29 cells after 24 h of treatment, and this was evidenced by the presence of apoptotic bodies [45].
Finally, we measured the intracellular ROS production in both cell lines because oxidative stress plays an important role activating intrinsic (mitochondrial) apoptosis in colon cancer cells by releasing of cytochrome c [46]. However, the extract did not induce mitochondrial depolarization and it also did not affect the basal level of intracellular ROS in both cell lines which suggest the extract exhibit protective effect on mitochondria, acts as antioxidant at the concentration evaluated taking into account the results of ORAC, DPPH and FRAP methods. A similar result was observed with vitamin C as antioxidant on SW480 cells that reduced significantly ROS production induced by the flavonoid lupulone [46]. Thus, these observations suggest that a mitochondrial-independent apoptosis on SW480 and SW620 cells is activated by the aqueous extract of Andean Berry which seemed to be not be related with the levels of intracellular ROS in this study.
In contrast, there are reports that some components of the berries induce an increase in ROS levels, such as proanthocyanidins obtained from cranberry (Vaccinium macrocarpon), which induce apoptosis in neuroblastoma cells in part due to the increase in the production of ROS [47]; it has also been observed that an extract rich in cranberry proanthocyanidins induces an increase in ROS production in esophageal adenocarcinoma cells at a concentration of 100μg /mL [48]. However, there are also reports of antioxidant activity of the extract in powder and the extract of the juice of this berry in hepatocarcinoma cells (HepG2), suggesting that these fruits can have both an antioxidant and pro-oxidant role [49].
In conclusion, the aqueous extract of Andean Berry displays an effective antiproliferative activity on SW480 colon adenocarcinoma cell line and their metastatic-derived SW620 cell line both in time- and concentration-dependent manner. The mechanisms studies showed that the Andean Berry extract triggered SW480 and SW620 apoptosis through the cell cycle arrest in phases S, G2/M and SubG0/G1, respectively, without promoting the mitochondrial membrane damage and oxidative stress suggesting the activation of intrinsic apoptotic pathway. These effects could be attributed to the bioactivity of the phenolic and anthocyanin content and the antioxidant capacity demonstrated by FRAP, ORAC and DPPH values. These observations highlighting the potential use of Andean Berry as source of bioactive compounds in colon cancer chemoprevention.
Footnotes
Acknowledgments
Author Sandra Arango-Varela was supported by a scholarship from the Department of Science, Tecnolohy and Innovation of Colombia (MinCiencias). Authors David Torres-Camargo and Camilo Reyes-Dieck were supported by the program Jóvenes Investigadores (Young Researchers) from Vicerrectoría de Investigación (Universidad de Antioquia) between 2019-2020. Author Maria Bibiana Zapata-Londoño was supported by program Pasantías Postdoctorales 848 from MinCiencias [grant number: 143–2020]. This work was also supported by MinCiencias Project number 58580 through the Program Ecosistema Científico [grant number FP44842-211-2018].
Funding
The authors report that this study was funded by MinCiencias Project number 58580 through the Program Ecosistema Científico [grant number FP44842-211-2018].
Conflict of interest
The authors have no conflict of interest to report.