
Abstract
BACKGROUND:
Increasing evidence has established neuroinflammation as the hallmark of neurodegenerative disorders such as Alzheimer’s disease (AD). However, despite the underlying immunological mechanisms are far from being understood, the involvement of excessive activation of microglia is attracting more and more attention.
OBJECTIVE:
In the present study, we investigated the protective effect of black raspberry (BRB) anthocyanins on LPS-induced neuroinflammation in BV2 microglia.
METHODS:
LPS-induced mouse BV2 microglia were treated with black raspberry anthocyanins and the levels of NO, ROS, IL-1β and IL-18 produced by the cells were measured to determine the extent of oxidative stress and inflammatory response.
RESULTS:
The results showed that BRB anthocyanins reduced the production of ROS in LPS-induced BV2 microglia by down-regulating the level of NOX2 and its downstream factors, including thioredoxin-interacting protein (TXNIP) and NOD-like receptor protein 3 (NLRP3) inflammasome. Furthermore, BRB anthocyanins inhibited the secretion of Interleukin-18 (IL-18) and Interleukin-1β (IL-1β), eventually attenuating the LPS-induced inflammatory response of BV2 microglia.
CONCLUSIONS:
BRB anthocyanins might play an important neuroprotective role in inflammation-related neurodegenerative disease, potentially, by down-regulating the NOX2 /TXNIP/ NLRP3 signaling axis in brain microglia.
Introduction
Neuroinflammation is observed in many brain disorders, especially in those with neurodegenerative conditions such as Alzheimer’s disease, Multiple Sclerosis and Parkinson’s disease [1, 2]. Accumulations of misfolded and aggregated proteins are a key feature of the pathology of all of the major neurodegenerative diseases. The misfolded and aggregated proteins bind to pattern recognition receptors on microglia or astroglia and trigger an innate immune response, characterized by the release of inflammatory mediators, which contribute to the progression and severity of the disease [3]. With the aging population on the rise globally, elderly-related diseases are gradually receiving more public concern [4, 5]. Among these diseases, neurodegenerative diseases might incur the heaviest burden on both the health care system and the family members of the patients. Thus, exploring effective means of therapeutics that are less expensive and with fewer side effects is a matter of priority. Small-molecule compounds targeting neuroinflammation arising from neurodegenerative disorders are likely to lead to future preventive or therapeutic strategies for neurodegenerative diseases [6].
Microglia, located in the central nervous system (CNS), play a critical role in the immune regulation of the CNS [7, 8]. Microglia are mononuclear macrophages derived from bone marrow hematopoietic stem cells that enter the CNS through a blood-brain barrier-deficient region of the brain [9]. During the development of the nervous system, microglia phagocytose non-viable neurons and oligodendrocytes, thereby regulating cell differentiation and apoptosis. Persistent stimulation of microglia by external signals can lead to their over-activation, resulting in the secretion of excessive amounts of ROS, cytokines and neurotoxic factors. This can eventually lead to neuronal damage and multiple pathological changes associated with inflammation. Indeed, excessive activation of microglia is a hallmark of neuroinflammation [10].
Oxidative stress caused by the overproduction of ROS can result in intracellular oxidation, leading to extensive cellular and molecular damage. It is a major factor in the development of multiple diseases. In microglia, NOX2 (NADPH oxidase 2), is a significant source of ROS [11, 12]. NOX2 also plays a crucial role in regulating the formation of ROS [13]. It contains six subunits: two catalytic subunits (gp91phox and p22phox) on the membrane and four regulatory subunits in the cytoplasm (p47phox, p67phox, p40phox, and Rac1/2). When macrophages are exposed to external stimuli (such as LPS), all the six subunits of NOX2 are activated, one after the other, and the enzyme functions as a whole complex in the regulation of ROS production [14, 15]. ROS production is also associated with the assembly of the NLRP3 inflammasomes. NLRP3 inflammasome is a multiprotein complex composed of the proteins NLRP3, ASC, and pro-caspase-1. It is located in the cytoplasm and is responsible for the maturation and secretion of the cytokines IL-18 and IL-1β [16, 17]. The NLRP3 inflammasome is associated with the development of several inflammatory-related diseases, such as cancer, cardiovascular disease, and neurodegeneration. In neuroinflammation, the NLRP3 inflammasome in microglia can be activated by a large spectrum of pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), such as LPS [18, 19].
Previous studies have shown that the consumption of berries can lead to a reduction in blood sugar and cholesterol levels [20, 21] and prevent the development of cardiovascular diseases [22, 23]. Recently, more research has focused on the neuroprotective effects provided to the CNS by berries or their major components such as anthocyanins [24, 25]. Black raspberry fruit is moderately sweet, juicy, and nutritious, and it contains minerals, vitamins, dietary fiber, and polyphenols such as quercetin, ferulic and chlorogenic acids, ellagitannins, phytohormones, and anthocyanins [26]. Black raspberry (BRB) anthocyanins, which have strong free radical scavenging activity, have been shown to exhibit anti-oxidative, anti-inflammatory, and anti-tumor effects [27]. Our previous studies have shown that BRB anthocyanins can exert chemoprevention effects in a mouse model of colorectal cancer through regulating gut microbiota and epigenetic alterations like up-regulating miRNA 24-1-5p, miRNA 483-5p, and demethylating the promoter of SFRP2 [28–30]. The impact of chronic intestinal inflammation and dysbiosis on the development of neurological disorders has aroused great interest in recent years [31], and dietary polyphenols have been proposed as a novel strategy to modulate microbiota-gut-brainaxis [32].
In the present study, we investigated the effects of BRB anthocyanins on the fate of LPS-induced microglia by monitoring the changes in inflammatory factors and critical proteins induced in these cells by LPS in the absence or presence of BRB anthocyanins.
Materials and methods
Reagents
FBS (Cat.no.900408) and DMEM (Cat.no.SH30022.01) were purchased from Hyclone (Logan, UT, USA). LPS was obtained from Sigma Aldrich (Cat.no.L6529; St. Louis, MO, USA). GSK2795039 (NOX2 inhibitor, GSK for short) was obtained from MedChemExpress (Cat.no.#HY-18950/CS-5988; MCE, Europe) [33]. Primary antibodies against NOX2 (Cat.no.ab80508), NLRP3 (Cat.no.ab214185), caspase-1 (Cat.no.ab138483), TXNIP (Cat.no.ab210826) and TRX (Cat.no. ab273877) were obtained from Abcam (Bristol, UK). CCK-8 kit (a cell counting kit) was purchased from Sagecreation (Cat.no.Q06014; Shanghai, China). Griess reagent kits for nitrite (Cat.no.S0021), DCFH-DA (Cat.no.S0033) and membrane protein extraction (Cat.no. P0033) were purchased from Beyotime Institute of Biotechnology (Jiangsu, China). ELISA kits for IL-18 (Cat.no.MM-0169M1) and IL-1β (Cat.no.MM-0040M2) were obtained from Meimian Biotechnology (Jiangsu, China). PrimeScript RT Master Mix real-time kit was purchased from Takara (Cat. No. RR036Q; Dalian, China).
Preparation of BRB anthocyanins
BRB anthocyanins (purity > 90%, HPLC), which were extracted from the mature fruits of black raspberry, were supplied by Professor Yuqing Zhao from Shenyang Pharmaceutical University. BRB anthocyanins are composed of four ingredients: cyanidin-3-O-rutinoside (16.91 mg/g), cyanidin-3-O-glucoside (2.63 mg/g), cyanidin-3-O-xylosylrutinoside (0.73 mg/g), and the less abundant cyanidin-3-O-sambubioside [34].
Cell culture
The murine microglial cell line BV2 was a gift from the College of Life Sciences, Northeastern University, China. BV2 cells were cultured in DMEM supplemented with 10% FBS, 100 U/ml penicillin, and 100 mg/ml streptomycin at 37oC in the presence of 5% CO2. BV2 cells were seeded in 25 cm2 cell-culture flasks, and the medium was replaced with fresh medium every other day.
Cell viability assay
The potential toxic effect of LPS on BV2 microglia was evaluated by incubating the cells in medium containing different concentrations of LPS (0.01, 0.1, 1 and 10 μg/ml) for 24 h followed by cell viability assay using a CCK8 assay kit according to the manufacturer’s instructions. In this assay, the BV2 cells were incubated with 10 μl CCK-8 solution for 2 h. The formazan dye in the cells was then dissolved with DMSO and the optical density of the sample was measured at 450 nm wavelength with a microplate reader.
Measurement of NO production
BV2 microglia were incubated with medium containing LPS (0.01, 0.1, 1 and 10 μg/ml) for 24 h. After that, 50 μl of the culture medium was taken and the concentration of nitrite (NO) in the sample was determined according to the Griess reaction [35] and with reference to a standard curve of sodium nitrite (NaNO2).
LPS-induced BV2 inflammatory cell model
The concentration of LPS that induced a high level of nitrite production but a minimum loss of cell viability was chosen to establish the LPS-induced BV2 inflammatory cell model. BV2 cells were cultured in 25 cm2 cell-culture flasks under the same conditions as described above until they reached 70–80% confluence. The monolayer in the flask was then digested with 1 ml of 0.25% trypsin (without phenol red and EDTA) and the digestion was terminated by the addition of 2 ml of DMEM medium containing 10% FBS. The cell suspension was transferred to a 5 ml round-bottom centrifuge tube and centrifuged at 225×g for 5 min. The cell pellet was then resuspended in 1 ml of DMEM medium and seeded in a 96-well plate at a density of about 104 cells per well. After attachment, the cells were exposed to the desired concentration of LPS for 24 h, yielding LPS-induced BV2 inflammatory microglia. To evaluate the effect of BRB anthocyanins on the nitrite production and viability of these LPS-induced BV2 inflammatory microglia, the culture medium was replaced with fresh medium containing different concentrations (3, 10, 30 and 100 μg/ml) of BRB anthocyanins followed by 24 h of incubation. After that, the level of nitrite in the culture supernatant and the loss of cell viability were determined as described above and compared with those of LPS-induced cells only and the control cells (no LPS induction).
From these experiments, suitable concentrations of BRB anthocyanins were chosen for further evaluation of the effect of BRB anthocyanins on LPS-induced inflammation. The LPS-induced microglia were incubated without or with the desired concentrations of BRB anthocyanins or 5 μM GSK2795039 (NOX2 inhibitor) for 24 h and then subjected to different assays as described below. BV2 microglia not induced with LPS were used as control cells.
Determination of ROS generation by DCHA-DA
The level of ROS in the cells was measured with a DCFH-DA kit. In the presence of ROS, DCFH is oxidized to form DCF, a fluorescent substance. The intensity of the fluorescence is directly proportional to the level of intracellular ROS, which can be determined by measuring the fluorescence of DCF. The fluorescence intensity of the cells was measured by a TECAN fluorescence microplate reader with excitation and emission wavelengths set at 488 nm and 525 nm, respectively.
RNA extraction and quantitative PCR
Total RNA was extracted from the cells using Trizol Reagen. The RNA obtained was quantified with a NanoDrop spectrophotometer and used to synthesize the cDNA by reverse transcription. The cDNA was then amplified by quantitative PCR (qPCR) with primers specific for IL-1β, IL-18, iNOS, NOX2 (gp91phox, p47 phox), NLRP3, ASC, caspase-1, TXNIP, TRX, or GAPDH. The primer sequences used in qPCR are listed in Table 2.
List of abbreviations used in this study
List of abbreviations used in this study
List of primers used in quantitative PCR
Cells were lysed with ice-cold RIPA buffer (RIPA:PMSF = 99:1). The total protein concentration in the cell extract was quantified with a BCA protein assay kit. A total of 20 μg protein from each sample was resolved in 10% SDS-polyacrylamide gel and the protein bands in the gel were then transferred to a nitrocellulose membrane. The membrane was blocked with 5% BSA in TBS-Tween 20 for 1 h, and then incubated with a primary antibody directed against NOX2 (gp91phox) (1:1000), NLRP3 (1:1000), caspase-1 (1:2000), TXNIP (1:1000), TRX (1:1000) or β-actin (1:1000) for overnight at 4 °C. After that, the membrane was washed in TBST and then incubated with a 1:5000 dilution of HRP-conjugated anti-rabbit IgG for 2 h at room temperature followed by washing in TBST. Detection of positive signals in the blot was carried out using Image J software.
Confocal microscopy assay
Cell slides were washed several times with pre-cold PBS, fixed in 4% paraformaldehyde for 20 mins, and then permeabilized with 0.5% TritonX-100 for another 20 min. The slides were incubated overnight with a 1:1000 dilution of anti-NOX2 (gp91phox) antibody and then with a green fluorescent protein-labeled secondary antibody for 2 h. After that, the slides were mounted in a DAPI-mounting medium and observed with a confocal microscope.
Measurement of IL-18 and IL-1β Levels by ELISA
BV2 microglia were cultured in a 96-well plate at a density of 104 cells/well and treated as described above. The levels of IL-18 and IL-1β in the culture supernatant were measured using an ELISA kit according to the manufacturer’s instructions. The absorbance of the plate was measured at 450 nm using a microplate reader.
Statistical analysis
All graphical data were plotted using GraphPad Prism 7 (GraphPad InStat Software, USA). Statistical significance of differences between control and treated samples were calculated using analysis of variance (ANOVA) and values of P < 0.05 were considered statistically different.
Results
BRB anthocyanins reduce the release of nitrite in LPS-induced BV2 microglia
A cellular inflammatory model of BV2 microglia was established via activation by LPS. BV2 microglia were activated by incubation with 0.01, 0.1, 1 and 10 μg/ml LPS. All concentrations of LPS were able to cause a significant (P < 0.001) increase in the level of nitrite released compared with the control (no LPS treatment) (Fig. 1A). Furthermore, the extent of increase was dependent on the concentration of LPS, with the concentration of nitrite in the cultures treated with 1 and 10 μg/ml being more than 4-fold the concentration in the control culture. Considering that 10 μg/ml LPS exerted a stronger effect on the viability of the microglia than 1 μg/ml LPS (Fig. 1B), 1 μg/ml LPS was used to establish the LPS-induced inflammatory cell model for all subsequent experiments. In addition to the change in the level of nitrite released, the effect of LPS on the morphology of BV2 microglia was also observed. Larger cells indicative of inflammatory cells were obviously present among the LPS-induced cells, whereas these cells were absent in the control cells (Fig. 1C). LPS-induced cells treated with 3, 10, 30 and 100 μg/ml BRB anthocyanins produced significantly lower concentrations of nitrite in the culture supernatant (P < 0.001) compared with those not treated with BRB anthocyanins, although these concentrations of nitrite remained significantly higher than that of the control cells (Fig. 1D). The influence of BRB anthocyanins on the viability of LPS-induced cells was relatively mild at the 3–30 μg/ml range, although it was obviously stronger at the 100 μg/ml concentration (Fig. 1E). Based on these results, the effects of 3 and 30 μg/ml BRB anthocyanins on the mRNA level of inducible nitric oxide synthase (iNOS) were examined by qPCR. The result showed that both concentrations of BRB anthocyanins significantly (P < 0.001) reduced the mRNA level of iNOS, with the 30 μg/ml concentration yielding more than 50% reduction compared with no BRB anthocyanins treatment (Fig. 1F) Taken together, the results clearly demonstrated that the success of the LPS-induced inflammatory cell model and established the protective effect of BRB-anthocyanins against LPS-induced inflammation.

Effects of BRB anthocyanins on the inflammatory status of LPS-induced BV2 microglia. Nitrite production (A), Cell viability (B) and cell morphology (C) of untreated BV2 microglia (control) or BV2 microglia treated with LPS. Nitrite level (D), cell viability (E), and iNOS mRNA level of untreated BV2 microglia, and LPS-induced microglia treated without or with different concentrations (3, 10, 30,100 μg/ml) of BRB anthocyanins (BRBA) for 24 h. Relative mRNA abundance in untreated cells was assumed to be “1” (control). The red arrow in (C) indicates activated microglia. All graphical data are the means±SDs from three independent experiments. ‘##’ and ###’ indicate significantly different from the control group at the P < 0.01 and P < 0.001 levels, respectively. ‘***’ indicates significantly different from the LPS-induced group without additional treatment at the P < 0.001 level.
To further demonstrate the protective effect of BRB anthocyanins against LPS-induced inflammation in BV2 microglia, the effect of BRB anthocyanins on ROS production in these cells was determined using the fluorescence probe DCFH-DA. The level of fluorescence in LPS-induced BV2 microglia was greatly enhanced compared with the control (no LPS induction) cells as revealed by fluorescence microscopy (Fig. 2A), consistent with a significant (P < 0.001) increase in ROS production (Fig. 2B). However, LPS-induced BV2 microglia treated with 3 or 30 μg/ml BRB anthocyanins all exhibited greatly reduced fluorescence, consistent with a significant reduction in ROS production. Furthermore, the level of ROS produced by the cells treated with 30 μg/ml BRB anthocyanins being comparable with that produced by the cells treated with the NOX2 inhibitor GSK2795039, indicating that BRB anthocyanins could suppress the production of ROS in LPS-induced BV2 microglia.
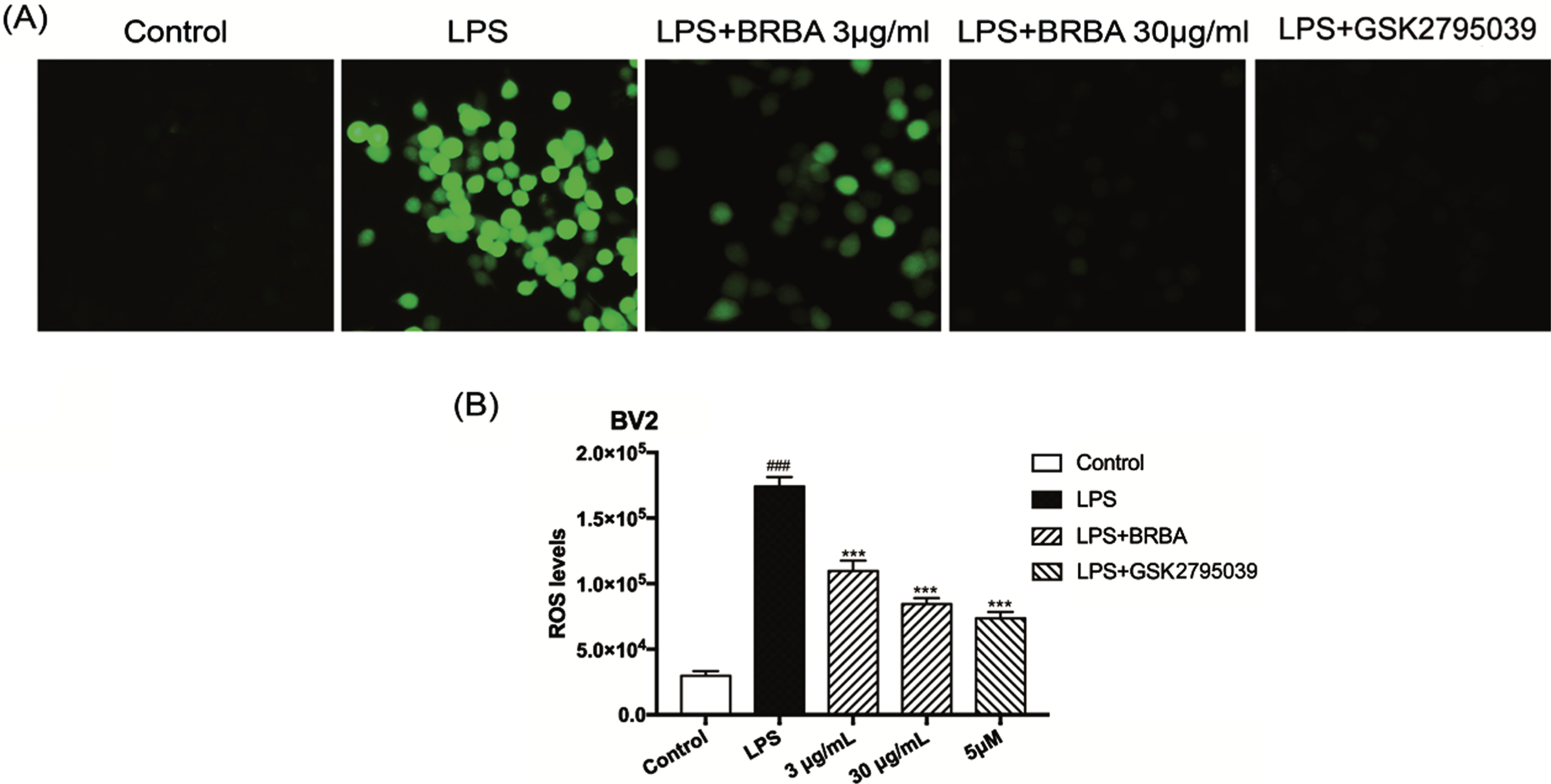
Effect of BRB anthocyanins on the release of ROS in LPS-induced BV2 microglia. BV2 microglia were either not treated (control) or induced with 1 μg/ml LPS followed by treatment with BRB anthocyanins (3 and 30 μg/ml) or 5 μM GSK2795039 for 24 h. The level of ROS in the cells was then quantitatively determined by measuring the fluorescence intensity of the probe DCFH-DA. (A) Representative images from fluorescence microscopy. (B) Quantitative comparison of ROS levels from different groups. Data are the means±SDs from three independent experiments. ‘###’ and ‘***’ indicate significantly different from the control group and LPS-induced group without additional treatment, respectively, at the P < 0.001 level.
ROS are derived from superoxide anions that are generated by NOX2 in the microglia. According to the above results, BRB anthocyanins decreased the release of LPS-induced ROS, and therefore, the effect of BRB anthocyanins on NOX2 expression was investigated and measuring the changes in protein level of the catalytic subunit (gp91phox) of NOX2. The result demonstrated that BRB anthocyanins significantly reduced the protein level of gp91phox, and the effect of 30 μg/ml was close to that exerted by GSK2795039 (Fig. 3A). Next, the effect of BRB anthocyanins on gp91phox as well as the regulatory subunit (p47 phox) of NOX2 was investigated at the mRNA level. BRB anthocyanins were found to significantly reduce the mRNA level of gp91phox while having no effect on the mRNA level of p47 phox (Fig. 3B). Furthermore, in situ expression of gp91phox in BV2 cells was observed under confocal microscopy following the labeling of gp91phox with a fluorescence-tagged secondary antibody. LPS-induced BV2 microglia that were treated with BRB anthocyanins or GSK2795039 showed reduced fluorescence intensity compared with the untreated but LPS-induced cells, indicating a loss of gp91phox in the treated cells (Fig. 3C). BRB anthocyanins, therefore, appeared to suppress the production of ROS through down-regulating the expression of NOX2, both at the transcription and translation levels.

Effect of BRB anthocyanins on the expression of gp91phox subunit in LPS-induced microglia. BV2 microglia induced with 1 μg/ml LPS were treated without or with BRB anthocyanins (3 and 30 μg/ml) or 5 μM GSK2795039, while control BV2 microglia were not given LPS or other treatments. The expression of the two subunits (gp91box and p47phox) of NOX2 in the cells was then measured by western blot and qPCR. (A) The expression of gp91phox as determined by western blot. Only a representative blot is shown. The plot beside the blot shows the expression of gp91phox as determined from the intensity of the bands in the blot. (B) The expression of gp91phox and p47phox as determined by qPCR. All graphical data are expressed as means±SDs from three independent experiments. squo###′ indicates significantly different from the control at the P < 0.001 level. ‘*’ and ‘***’ indicate significantly different from the LPS-induced group without additional treatment at the P < 0.05 and P < 0.001 levels, respectively. (C) The influence of BRB anthocyanins on gp91phox in situ as observed by confocal microscopy. A representative image from each group is shown.
BRB anthocyanins were found to have a significant inhibitory effect on the cellular indices of oxidative stress in the above-mentioned experiments. Oxidative stress is often accompanied by the development of inflammation. Therefore, the effect of BRB anthocyanins on the biomarkers associated with inflammation was investigated. Four such biomarkers were chosen, and these were NLRP3, caspase-1, TXNIP and TRX. The expression of NLRP3, caspase-1 and TXNIP (all are pro-inflammatory factors) in LPS-induced BV2 microglia was decreased at both the mRNA (Figs. 4A-B) and protein (Fig. 4C) levels after treatment with BRB anthocyanins. However, the expression of thioredoxin (TRX) was enhanced at both the mRNA and protein levels, consistent with its role as an anti-inflammatory factor. The results of these experiments strongly suggested that BRB anthocyanins protected BV2 microglia against LPS-induced inflammation by inhibiting the expression of pro-inflammatory proteins while enhancing the expression of anti-inflammatory proteins.
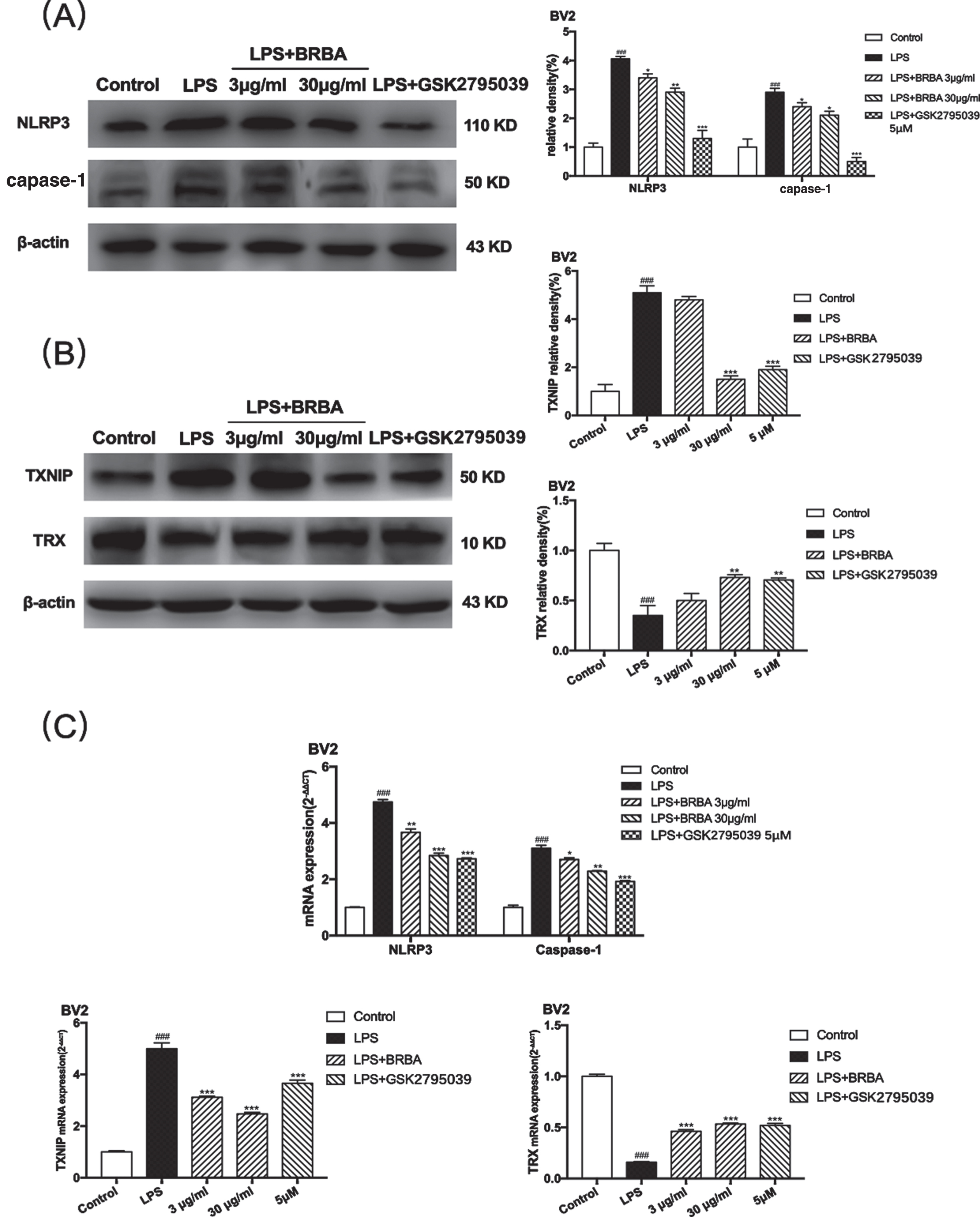
Effects of BRB anthocyanins on the factors associated with NLRP3 signaling pathway in LPS-induced BV2 microglia. BV2 microglia induced with 1 μg/ml LPS were treated without or with BRB anthocyanins (3 and 30 μg/ml) or 5 μM GSK2795039, while control BV2 microglia were not given LPS or other treatments. The levels of four inflammation biomarkers produced by these cells were then measured by western blot or qPCR. (A) NLRP3 and caspase-1 and (B) TXNIP and TRX protein levels in the cells as determined by western blot. Only representative blots are shown. The plots beside the blots compare the protein levels of NLRP3, caspase-1, TXNIP and TRX based on the intensities of the corresponding bands in the blots. (C) NLRP3, TXNIP, caspase-1 and TRX mRNA levels in the cells were determined by qPCR. Relative mRNA abundance in the control cells was assumed to be 1. All graphical data are the means±SDs from three independent experiments. ‘###’ indicates significantly different from the control at the P < 0.001 level. ‘*’, ‘**’ and ‘***’ indicate significantly different from the LPS-induced group without additional treatment at the P < 0.05, P < 0.01 and P < 0.001 levels, respectively.
In order to explore the effect of BRB anthocyanins on inflammation in LPS-treated BV2 microglia, changes in the levels of two pro-inflammatory cytokines, IL-18 and IL-1β, were measured. Compared with the control cells, LPS-induced BV2 microglia exhibited a significant increase in IL-18 and IL-1β levels in the culture supernatant as shown by ELISA (Figs. 5A & 5B). The extent of increase was more than three-fold for IL-18 and more than two-fold in the case of IL-1β. However, when LPS-induced BV2 microglia were treated with BRB anthocyanins, the levels of IL-18 and IL-1β decreased significantly (P < 0.01) compared with those not treated with BRB anthocyanins. The extent of reduction was as much as 40% for the 30 μg/ml treatment and about 15% for the 3 μg/ml treatment. Similarly, treating LPS-induced BV2 microglia with GSK also led to a significant reduction in both IL-18 (∼70%) and IL-1β (∼50%) levels relative to the untreated cells. The effect of BRB anthocyanins on IL-18 and IL-1β in LPS-induced BV2 microglia was also investigated by measuring the mRNA levels of both cytokines in the cells. The result was consistent with the ELISA data, whereby BRB anthocyanins treatment significantly reduced the mRNA levels of these two cytokines, with 30 μg/ml being more potent than 3 μg/ml (Figs. 5C-D). Taken together, the result strongly suggested that BRB-anthocyanins could ameliorate the extent of LPS-induced inflammation in BV2 microglia by suppressing the production of proinflammatory cytokines at both the transcription and translation levels.
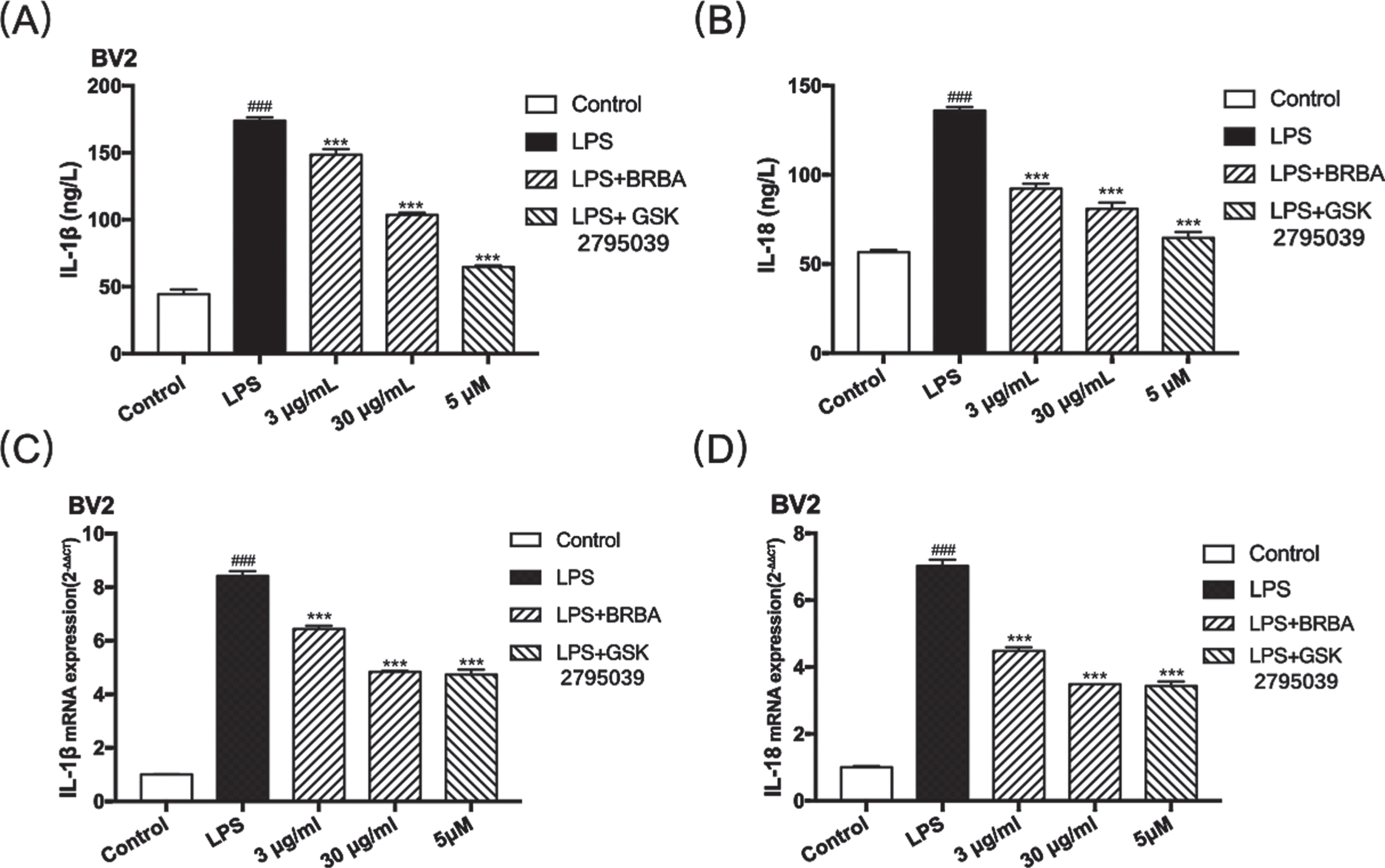
Effect of BRB anthocyanins on the secretion of IL-18 and IL-1β by LPS-induced BV2 microglia. BV2 microglia induced with 1 μg/ml LPS were treated without or with BRB anthocyanins (3 and 30 μg/ml) or 5 μM GSK2795039, while control BV2 microglia were not given LPS or other treatments. IL-1β (A) and IL-18 (B) levels in the culture medium were determined by ELISA, while IL-1β (C) and IL-18 (D) transcript levels in the cells were determined by qPCR and compared with those of the control cells. Data are the means±SDs from three independent experiments. ‘###’ indicates significantly different from the control at the P < 0.001 level, whereas ‘***’ indicates significantly different from the LPS-induced group without additional treatment at the P < 0.001 level.
AD is associated with the activation of microglia, and excessive activation will cause these cells to produce large amounts of pro-inflammatory and neurotoxic factors, resulting in neuronal loss and accumulation of amyloid Aβ [36, 37]. Therefore, identifying compounds that have anti-inflammatory activity and can inhibit the activation of microglia may be an alternative therapeutic approach for the treatment of neurodegenerative diseases. Several studies have found that anthocyanins are unique flavonoid compounds that can modulate several of the factors contributing to neuronal death, and interest in their use as therapeutics for neurodegeneration has grown in recent years [38–40].
Experiments conducted in the present study have led to the following observations: The participation of NOX2 in the inflammatory response against LPS-induced inflammation in BV2 microglia, which involved the up-regulation of NOX2, TXNIP, and NLRP3 inflammasome expression levels, and increased production of ROS and pro-inflammatory cytokines (IL-18 and IL-1β). The release of IL-18 and IL-1β from the cells was related to the assembly of the NLRP3 inflammasome. More importantly, a neuroprotective role for BRB anthocyanins in LPS-induced microglia was found and an underlying mechanism that could link AD with inflammation through the NOX2-TXNIP-NLRP3 pathway was discovered. These observations indicated the potential application of BRB anthocyanins in neuroinflammation-associated diseases exemplified by AD.
Recently, an increasing number of studies have focused on the role of NOX2 in CNS inflammation, and the potential value of NOX2 inhibitors for the treatment of diseases of the nervous system [41, 42]. ROS are highly active molecules produced during the normal cellular respiratory metabolism and they play a vital role in signaling pathways [12, 43]. There has been mounting evidence suggesting that neuroinflammation is often accompanied by the production of oxidative stress [44]. Moreover, the presence of ROS in microglia may serve as an early signal that triggers the production of cytokines, thereby participating in the endogenous inflammatory signaling. Both the mRNA and protein levels of NOX2 in LPS-induced BV2 microglia were significantly up-regulated (Fig. 1). Moreover, the level of ROS detected by DCFH-DA was also markedly increased in these cells (Fig. 2). BRB anthocyanins significantly decreased the expression level of NOX2 and reduced the production of ROS. These findings indicated that NOX2 might play a critical role in the neuroprotective effect of BRB anthocyanins, and by regulating the expression of NOX2, BRB anthocyanins could effectively regulate the production of ROS.
TXNIP and TRX are crucial components that regulate the cellular redox state and defense against oxidative stress. Oxidative damage caused by ROS can lead to the assembly of intracellular inflammasomes, mediated by TRX and its endogenous negative regulator TXNIP [45]. When cells undergo oxidative stress, the ROS produced promotes the dissociation of TXNIP from TRX and allows it to bind NLRP3 to activate the NLRP3 inflammasome. In the current study, we found that LPS treatment significantly up-regulated the protein levels of NLRP3 and TXNIP in microglia and the release of IL-18 and IL-1β by these cells. When LPS-induced BV2 microglia were exposed to BRB anthocyanins, oxidative indexes such as nitrite, ROS and the pro-inflammatory cytokines such as IL-1β and IL-18 were down-regulated. The mRNA levels of gp91phox, NLRP3, caspase-1, and TXNIP were also downgraded by BRB anthocyanins. The data indicated that regulation of the NLRP3 signaling pathway might play a critical role in the neuroprotective effect of BRB anthocyanins.
In summary, BRB anthocyanins might play a neuroprotective role by suppressing the production of pro-oxidative/pro-inflammation factors in LPS- induced BV2 microglia, such as NOX2, NLRP3 and TXNIP (Fig. 6). The current finding might shed more light on the potential of BRB anthocyanins as a future preventive or therapeutic strategy for neurodegenerative disease.

Schematic representation showing a possible neuroprotective role provided by BRB anthocyanins through the downregulation of NOX2/TXNIP/NLRP3 signaling.
Credit authorship contribution statement
Teng Mu: Data development, methodology, Writing-original draft. Yang Guan: Data development, software, validation. Tianqiao Chen: Software, methodology, resource. Shuning Wang: Data development, software. Mei Li: Data development, software. Alan K Chang: Revising the original draft and proofreading the final draft. Zhe Yang: Data development. Xiuli Bi: Project administration, supervision, conceptualization, methodology and writing the original draft.
Conflicts of interest
We confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Compliance with ethical standards
No statement.
Footnotes
Acknowledgments
This work did not receive any specific funding from public, commercial or non-profit sector funding agencies.
Funding
The research was partially financially supported by LiaoNing Revitalization Talents Program (#XLYC 1807058).