
Abstract
BACKGROUND:
Haskap (Lonicera L.) is as a new perspective berry species for growing in temperate region climate. According to nowadays knowledge haskap is absolutely self-sterile species hence the studies on pollination mode are required.
OBJECTIVE:
The aim was to evaluate new haskap cultivars of Canadian and Russian origin in terms of their matching for cross-pollination.
METHODS:
The overlapping of flowering time of cultivars for mutual cross pollination was selected. The effectiveness of pollination was assessed: in terms of pollen tube overgrowth through the pistil tissue and the quality of set fruit.
RESULTS:
The Russian cultivars bloomed much earlier than the Canadian cultivars. The stigma is most receptive in the freshly open flower stage and directly after the anther burst. The minimum qualitatively acceptable weight of a berry is 1 g, which corresponds to the formation of about 6 seeds in the fruit. The most compatible pairs of cultivars were: ‘Aurora’בJugana’, ‘Aurora’בSinij Utes’ and ‘Aurora’בVostorg’.
CONCLUSION:
The Russian cultivars bloomed much earlier than the Canadian cultivars, the stigma is most receptive in the freshly open flower, minimum qualitatively acceptable weight of berry is 1 g (what represents 6 seeds in fruit), best mutual pollinating cultivars are the cultivars within the same breeding group (Russian x Russian and Canadian x Canadian)
Introduction
The genus Lonicera L. (Caprifoliaceae Juss.) (haskap or honeysuckle) includes a large number of plant species occurring in the Northern hemisphere from temperate to subarctic climate zones [1]. The cultivation of honeysuckle is carried out in several countries, including Poland, Estonia, Romania, but the leaders are Russia and Canada [2–9]. Breeding programs often concern cultivar selection in terms of such traits as fruit production, fruit size, low fruit shedding tendency, no bitterness, taste, sugar-acid ratio or resistance to gray mould [10–13]. Japanese, Kuril and Russian genes are used mainly in Russia in breeding work on new haskap cultivars. Fertility of Russian breeding cultivars, cross-pollination and the possibility of effective pollination of cultivars are very important issue [9, 14–16]. In Canada, in breeding works Lonicera spp. forms originated from four geographical centres: Russia, Japan, Kuril Islands and Canada are used [7]. It was found that the origin from individual centres determined the features of the descendant forms, such as: higher plants, good fruit production –Russian and Japanese genotypes, early harvest –Russian and Canadian genotypes, late harvest, round fruit shape, fruit sticking too tightly to the peduncle –Japanese and Kuril genotypes, sweet fruit taste –Kuril and Canadian genotypes, unequal fruit maturation –Japanese genotypes, low yield –Kuril genotypes, elongated fruit shape, fruit tartness –Russian genotypes, small fruits –Canadian genotypes. Haskap cultivars inherited the pollen diversity and quality of pollen was influenced by the origin of the cultivar [17]. Depending on its geographical origin, pollen differed in fertility, size and anatomical features. It was found that pollination with own pollen between closely related cultivars resulted in poor fruit setting and its low quality. Origin genes can affect suitability for cultivation of Lonicera in various climatic conditions. It was reported that haskap genetic material shows a wide range of genetic variation, which affects phenological adaptation to temperate climate conditions. Cultivars originating from species with high winter frost resistance may show physiological disturbances in temperate climate due to inadequacy to high air temperatures in winter and early spring [18, 19]. Breeding of new cultivars using genetic material from all three groups (Russian, Japanese and Kuril) requires selection to avoid forms with early phenology. Cultivars with a high proportion of genes from Kamchatka and Siberia, where the growing season is very short, show phenological disorders in temperate climates, manifested by early summer dormancy and possibility of another flowering in the late autumn months. If positive temperature occurs in the late autumn for several days, absolute dormancy is interrupted and the plants begin to bloom. Similarly, early periodic warming, alternating with temperature drops, can cause haskap dormancy interruption, which in turn leads to damage to plants that have been awakened from dormancy [19–21]. Some researchers have attempted to demonstrate the influence of moderate climate on the correctness of micro- and macrosporogenesis and early stages of embryogenesis based on microscopic cross-sections [22].
Due to the great interest in planting this species, there is a need for agrotechnical research on cultivar distribution in the plantation for an effective cross-pollination. Many authors reported testing different systems of distribution and number of shrubs of cross-pollinating cultivars [7, 18]. In our earlier study [16] we reported investigations on flowering biology of 40 haskap cultivars in order to assess the biological characteristics of flowers that affect cultivar fertility. The current research is a continuation of previous experiments and includes mainly new cultivars from Canadian and Russian breeding.
The aim of the present research was to evaluate nine new haskap cultivars of Canadian and Russian origin in terms of their matching for cross-pollination. The results will help to answer several important questions: (i) Do the flowering dates of these cultivars overlap? (ii) How does stigma receptivity change at different stages of flower development? (iii) Which of the crossed haskap pairs are the most compatible? (iv) How can compatible pollination of crossed cultivar pairs improve fruit quality?
Material and methods
The experiments have been carried out in haskap (Lonicera spp.) commodity plantation located in the southern Poland (coordinates: altitude 251 meters above sea level; latitude 50°17’31”N, longitude 20°06’59”E). The shrubs were planted on sandy loam calcareous soils with a pH of 5.6, in a 1.2×4.0 m row system, mulched in the rows with black polyurethane fabric, turf was mowed in the inter-rows, the shrubs were fertigated as needed. The experiment included haskap berry cultivars originated from Russian selection: ‘Jugana’, ‘Sinij Utes’, ‘Vostorg’ and from Canadian selection: ‘Aurora’, ‘Boreal Beast’, ‘Boreal Beauty’, ‘Boreal Blizzard’, ‘Honey Bee’, ‘Indigo Gem’. Most of the cultivars have only recently been introduced to the commercial market.
Flowering periods, flower biometric measurements, number of pollen grains in flower, viability of pollen
Detailed observations of flower phenology were carried out during flowering of haskap cultivars. The stages of flowering of each cultivar were distinguished and the cultivars were segregated in terms of beginning of flowering. Biometric measurements of flowers were conducted for each of the haskap cultivars (in four replicates, of 30 flowers each, 4×30 = 120 flowers), including the following parameters: length and diameter of the sympetalous corolla [mm] and stamen and pistil length [mm]. Based on these measurements, calculations were made of the difference between the length of the style and the stamen, and the length of the perianth and the style in order to determine the best adaptation of flowers to effective pollination by insects. Osmia rufa and Bombus terrestris as insect pollinators were introduced to the plantation at the beginning of flowering. The numbers of pollen grains in the anther and flower [pcs] of haskap cultivars were evaluated using a Bürker’s hemocytometer. The sample consisted of ten anthers randomly collected from ten flowers of a given cultivar; the calculations were performed in four replicates. Observations were made using a Carl Zeiss Image M2 AXIO microscope at 10X magnification in white light. Directly before pollination, the viability of pollen grains [%] was determined using the Alexander colouring method. This dye stains viable pollen grains in a fuchsia color and dead pollen grains in green [16].
Stigma receptivity
Stigma receptivity of haskap cultivars was determined [23, 24]. Flowers were collected from each cultivar and classified according to the following developmental stages: stage 1 –closed flower, stage 2 –closed flower with protruding stigma, stage 3 –freshly opened flower, stage 4 –open anthers, stage 5 –brown anthers. In laboratory, in order to assess the receptivity of the stigma, a 3%H2O2 solution was applied to the surface stigma and stigma was observed under a Carl Zeiss Discovery 2.0 binocular magnifier. The receptive stigma actively releases oxygen in the form of gas bubbles, and a non-receptive stigma does not show this capacity. The percentage of receptive stigmas in each developmental stage of the flower was calculated (ten flowers were assessed at each stage).
Cross-pollination of haskap cultivars
Based on the earlier studies [16] it was found, that Lonicera is cross-pollinating genus and in the presented experiment the authors focused on the cross-pollination in order to select the best pairs of cultivars in terms of mutual compatible pollination. Twenty nine targeted manual crosses with pollen of a specific cultivar were performed. The control for these crosses was combination of open-pollination of flowers of each cultivar by insects with a pollen mixture. Cultivar combinations have been selected to overlap the flowering times. On the plantation experimental shoots with flowers in the balloon stage (four replicates, of about 100 flowers each) were isolated with agrotextile insulators. The anthers collected from flowers of pollinator cultivar were dried at 20°C for 24 hours, and then the pollen obtained from open anthers was poured into glass vials and stored at 6°C until pollination. Each flower was pollinated 2 or 3 times every two days during full blooming. Manual crossing was performed with a silicone plug dipped in glass vials with dried pollen. The pollen was applied to the flower stigmas and the presence of pollen grains on the stigmas was checked at using the handheld magnifier.
Microscopic observations of pollen tube growth through the style
At the end of flowering, some flowers after cross pollination were collected for microscopic analysis of pollen tube growth through the style, and some flowers were left for fruit development. This analysis has been used to determine the effect of pollen origin on the process of pollen tube overgrowth in the style tissue. The dissected flower pistils were fixed in FAA (formalin: ethyl alcohol: acetic acid, 8:1:1) for 10–12 h. The pistils were macerated in 30%NaOH solution for 2–3 h, and then the plant tissue was cleared with a 6%H2O2 solution. In the next step, after washing with water, the pistils were stained with aniline blue for 3 h. In the final stage, the preparations were closed with glycerol, and observations were conducted using a Carl Zeiss Image M2 Axio fluorescence microscope. Germination of pollen grains on the stigma and the growth of pollen tubes through style tissue to the base of the pistil were observed in a microscope under UV light. The numbers of overgrowing pollen tubes were assessed at three levels of the style: 1/3, 1/2 of length and the base of the pistil and expressed as percentages in relation to the number of germinating pollen grains on the stigma. A sample consisted of about 30 flowers from a combination.
Pollen germination index
The pollen germination index (PGI) was calculated for crossing pairs of cultivars and enabled to trace pollen grains germination on the stigma and pollen tube growth throughout the length of the style. It was possible to define the style region where the disturbance in pollen tubes growth occurred and the tubes were rejected. PGI was calculated according to the following formula [25].
where:
a –number of styles with pollen grains
b –number of styles, which pollen grains do not germinate
c –number of styles, which pollen grains germinate on the stigma
d –number of styles, which pollen tube grows into the style
e –number of styles, which pollen tubes growth into base of the pistil
Based on detailed microscopic observations, a graphical model of pollen tube growth through the style was elaborated.
Evaluation of fruit quality obtained after cross- and open-pollination
Mature fruits, from the experimental shoots left for development were harvested approximately 60 days after flower pollination. It was assessed in which combinations the fruits were obtained; moreover, biometric measurements of all harvested fruits from individual cross- and open-pollination combinations were made. The measurements concerned 520 fruits and included the following parameters: berry weight [g], berry length and width [mm], number of seeds in a berry [pcs]. Based on biometric measurements, the percentage of fruits with well-developed seeds was calculated, as well as the fruit shape ratio (berry length: berry width) and correlation between fruit weight and the number of well-formed seeds.
Statistical analysis
The results were statistically calculated using STATISTICA 13.3 (StatSoft Inc., Tulsa, OK, USA) and one-way analysis of variance and Tukey’s honest significant difference (HSD) test were performed at the probability level of α= 0.05. The correlation between the weight of one berry and the number of seeds in a berry was calculated on the basis of all fruits (N = 520).
Results
Flowering periods, flower biometric measurements, number of pollen grains in flower, viability of pollen
The cultivars differed in the time of individual flowering stages, the time of flowering was from 8 to 10 days for each cultivar. Cultivars of Russian origin (‘Jugana’, ‘Sinij Utes’ and ‘Vostorg’, and the Canadian cultivar ‘Aurora’ (which has Russian cultivar ‘Soloviej’ in its pedigree) flowered the earliest. Afterwards the Canadian cultivars ‘Honey Bee’ and ‘Indigo Gem’ flowered, and ‘Boreal Beast’, ‘Boreal Beauty’ ‘Boreal Blizzard’ flowered at the latest. The flowering periods were shifted 5–6 days in relation to the full flowering date between cultivars.
The flowers of the cultivars differed significantly in several biometric parameters (Table 1). The cultivar ‘Jugana’ had the shortest perianth length, while the remaining cultivars did not differ significantly in this trait. Perianth width varied from 4.33 mm (‘Boreal Blizzard’) and 4.40 mm (‘Jugana’) to 6.79 mm (‘Sinij Utes’). The cultivars ‘Boreal Beast’ and ‘Sinij Utes’ were characterized by very long pistils. The cultivar ‘Sinij Utes’ had the longest stamen. The differences between the length of the pistil and the stamen as well as the perianth and the pistil were calculated. The obtained numerical values can be used to assess the effectiveness of pollen grains depositing on the stigma of haskap flowers by pollinating insects, which affects one of the most important factors of pollination efficiency. In the case of the cultivar ‘Vostorg’, the pistil was hidden in the perianth because it was shorter than the perianth by 2 mm. The cultivars differed significantly in the number of pollen grains in the flower. Two groups could be distinguished among the assessed cultivars, the group of cultivars with the highest number of pollen grains in the flower (‘Aurora’, ‘Boreal Beast’ and ‘Sinij Utes’) and the group with a significantly smaller number of them. Pollen viability was at a high level (from 89.6 to 97.1%) and did not differ significantly between cultivars, except for the cultivar ‘Jugana’, which was characterized by the lowest pollen viability among the cultivars (78.0%).
Morphology and quality of haskap flowers
Morphology and quality of haskap flowers
*There is always five stamens in haskap flowers.
Stigma receptivity, defined as the ability of the flower to accept pollen, changes during its life. Five stages of flower development were distinguished for haskap flowers. Table 2 shows that stigma receptivity varied depending on the stage of flower development.
Receptivity of haskap flower stigmas
Receptivity of haskap flower stigmas
*1–5 stages of flower development, see detail description in Material and methods.
Stages 1 and 2 correspond to closed flower buds, but in stage 2, the pistil clearly grows above the closed syntepalous corolla. The highest percentage of receptive stigma was recorded in stages 3 and 4. Haskap flowers were characterized by a lower stigma receptivity at the early stage of development (stage 1) as well as at the end of flowering (stage 5) (Fig. 1a-c).

a-f. Receptivity of haskap flower stigmas and overgrowth of pollen tubes through pistil tissue. a. Non- receptive young stigma. b. Receptive stigma - visible oxygen bubbles. c. Non- receptive old stigma. d. Large population of germinating compatible pollen grains on the stigma and growing pollen tubes in pistil tissue. e. Inhibition of pollen tube growth on a stigma after incompatible pollination. f. Numerous pollen tubes growing to the base of the pistil after compatible pollination.
Table 3 presents the results of manual cross-pollination (29 pairs of cultivars) and open pollination by insects of 9 cultivars (pollen mixture).
Overgrowth of pollen tubes through the pistil tissue and pollen germination index (PGI) depending on the pollinated cultivars
Overgrowth of pollen tubes through the pistil tissue and pollen germination index (PGI) depending on the pollinated cultivars
It was found that the number of pollen grains on the stigma differed depending on the pair of crossed cultivars. The first inhibition of pollen tubes growth in the pistil occurred on the section from the stigma to 1/3 of the style. The percentage of pollen tubes observed in 1/3 of style length was calculated in the relation to the number of germinating pollen grains on the stigma. Three groups of cultivars could be distinguished. The first group included those crossings for which the percentage of pollen tubes was about 80–100%, the second group consisted of crossings where the percentage of pollen tubes was at the level of about 50–79%, and the third group consisted of crossings where the percentage of pollen tubes was below 49%.
In the middle of the style, there was a second reduction in the number of pollen tubes in relation to the number of germinating pollen grains on the stigma, and similarly, three groups of crosses were distinguished. The first group included crosses for which the percentage of pollen tubes was at a level of 40–70%, the second group from 20–39%and the third group below 20%. In the base of the pistil, the percentage of pollen tubes was low, two groups of crossings were distinguished, the first from 10 to 26%of growing pollen tubes in relation to the germinating pollen grains on the stigma and the second group with a very low percentage of pollen tubes (less than 10%) (Fig. 1d-f).
Pollen germination index (PGI) proved that the cultivars were well matched in terms of pollination efficiency (Table 3). If the value of this index equals at least 2.0, it proves good compatibility of cultivars. Of pollination combinations (29 pairs of crossed cultivars and open pollination of 9 cultivars), the value of this coefficient was 2.0 in 8 combinations. On this basis of analysing the growth of pollen tubes in successive zones of the style models showing a growth of pollen tube through the style were elaborating (Fig. 2).

Model of pollen overgrowth through the pistil of haskap flowers depending on the origin of the cultivars. 1. combination of pollination: Russian cultivars×Canadian cultivars. 2. combination of pollination: Canadian cultivars×Russian cultivars. 3. combination of pollination: Russian cultivars×Russian cultivars. a - average number of germinated pollen grains on stigma. b - average number of pollen tubes in 1/3 length of pistil. c - average number of pollen tubes in 1/2 length of pistil. d - average number of pollen tubes at the base of pistil.
Figure 2 presents three models of pistils with combinations of crossed cultivars according to their origin: 1 –Russian×Canadian cultivars, 2 –Canadian×Russian cultivars, 3 –Russian×Russian cultivars. The models show that the first difference in the average number of deposited and germinating pollen between the distinguished groups of cultivars already appeared on the stigma, despite the fact that a similar number of pollen grains were manually applied to the stigma of each combination. Combination Russian×Russian cultivars was most favourable (the average number of pollen tubes was the highest both on the stigma and in individual length zones of the style). In this combination, an average of 11.1 pollen tubes grew to the base of the pistil, which allowed good quality fruit setting. Contrary to that group of crosses, combination Canadian×Russian cultivars was the worst, in terms of both the average number of pollen grains present on the stigma and the average number of pollen tubes in the subsequent length zones of the style.
The fruits obtained as a result of individual crossing pairs and open-pollination by insects with a pollen mixture were assessed (Table 4).
Biometric measurements of haskap fruits obtained after open- and cross-pollination
Biometric measurements of haskap fruits obtained after open- and cross-pollination
Fruits were obtained in 28 combinations of crosses. However, no fruit was obtained in the following 10 pairs of crossed cultivars: ‘Boreal Beast’בJugana’, ‘Boreal Beast’בVostorg’, ‘Boreal Beauty’בJugana’, ‘Boreal Beauty’בSinij Utes’, ‘Boreal Beauty’בVostorg’, ‘Honey Bee’בJugana’, ‘Honey Bee’בSinij Utes’, ‘Honey Bee’בVostorg’, ‘Indigo Gem’בSinij Utes’ and ‘Vostorg’בJugana’. The average fruit weight ranged from 0.6 to 1.8 g (Table 4). The length and width of the fruits were measured and the data was used to calculate the fruit shape factor. The resulting shape factor proved that most of the obtained fruits had an elongated shape, with the exception of fruits obtained from open pollination of ‘Boreal Beauty’ and ‘Indigo Gem’ cultivars and in fruits resulting from the cross between the cultivars ‘Jugana’ and ‘Indigo Gem’. But in the case of the latter cross, only 3 fruits were obtained due to the large incompatibility between the crossed cultivars. The obtained fruits differed in the presence of seeds. Only in the case of 17 pollination combinations, 100%of fruits developed seeds (Table 4). The average number of seeds in a berry ranged from 0.9 to 19.3. After compatible pollination, large, well-formed fruits formed, with numerous well-developed seeds, while fruits resulting from incompatible pollination did not contain seeds, fruits were often deformed, or did not grow to normal size or remained green for a long time. From all pollination combinations, 520 fruits were obtained, and correlation between the number of seeds in a berry and the weight of a berry was calculated for each berry on the basis of individual measurements. The correlation was found to be significant, which means that the number of seeds in a berry significantly influenced fruit weight (Fig. 3).
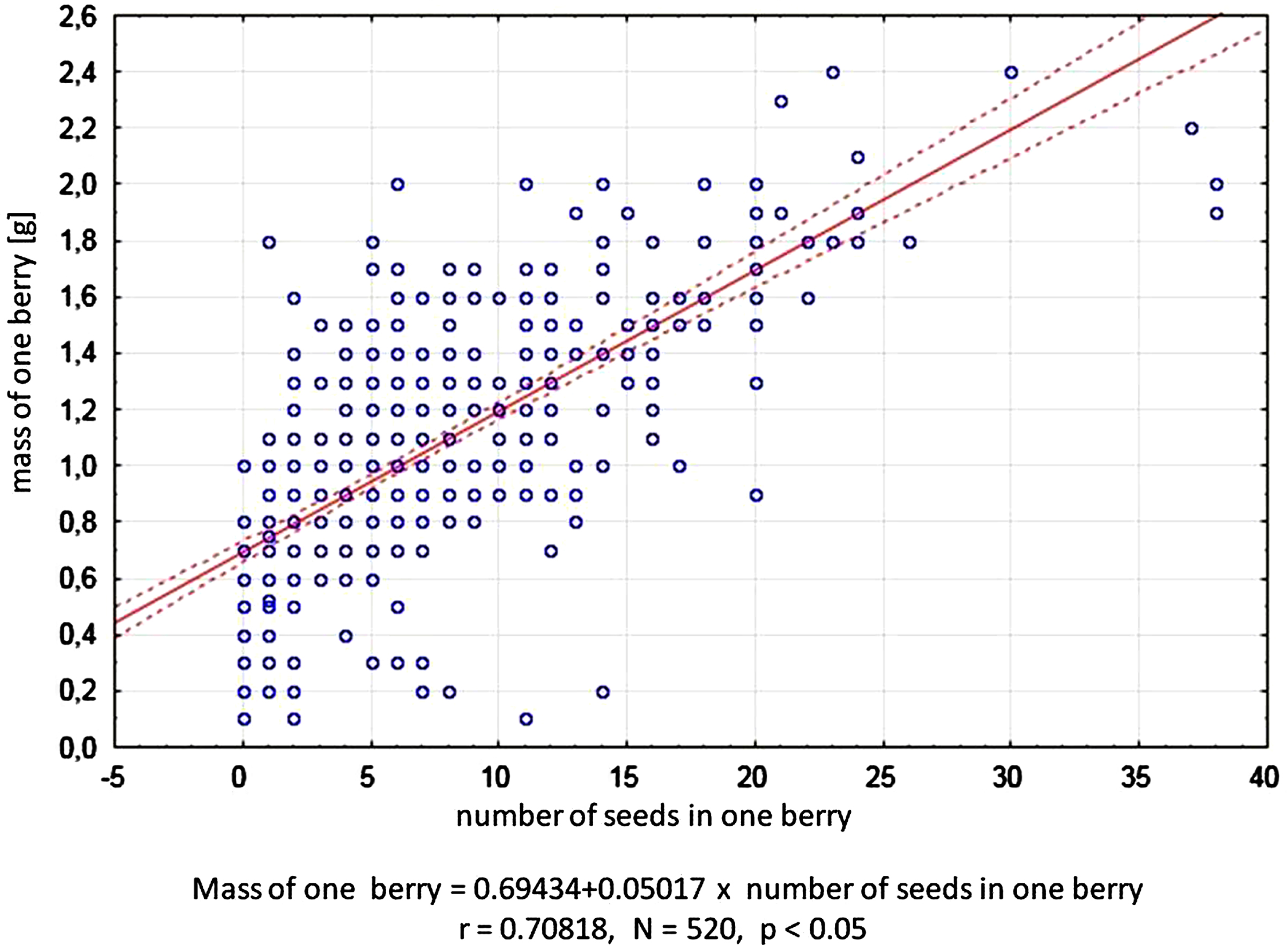
Correlation between the number of seeds in a fruit and the weight of the haskap fruit.
The flowers of cultivars of the genus Lonicera develop on the border of one-year shoots. They always appear in pairs of two flowers for one ovary [16, 27]. Haskap is a cross-fertile species, which means that it requires pollination of flowers with pollen of a different cultivar for effective fruit setting [15, 16]. The compatibility of cultivars with regard to cross-pollination depends on genetic, physiological and phenological mechanisms that prevent fertilization with their own pollen or with pollen of closely related cultivars. Hence, the first condition for the correct selection of cultivars in terms of ovules fertilization, fruit setting and obtaining a good yield is matching them in respect of flowering time. The flowering times of individual cultivars were shown to differ depending on weather conditions prevailing in a given year [7]. In the discussed experiment, the flowering periods differed depending on the cultivar. Each of the cultivars bloomed from 8 to 10 days, with a mutual shift in flowering of 5–6 days. Differences in phenology of cultivars are observed depending on the percentage of individual genotypes (Kuril, Russian or Japanese) which were use in breeding cultivars. The cultivars with the highest proportion of Kuril and Russian genes begin vegetation as the first ones [7, 19]. In the discussed experiment, the cultivars ‘Aurora’, ‘Jugana’, ‘Sinij Utes’ and ‘Vostorg’ of Russian origin started flowering first, which indicated the dominance of Russian genes determining the early start of flowering. The presence of Japanese genotypes in the cultivars ‘Boreal Beast’ and ‘Boreal Beauty’ causes the late start of flowering, which was recorded in the current experiment. When designing the experiment, the overlapping flowering time of each cultivars pairs was taken into account. At overlapping of flowering time of cultivar pairs might be assumed that effective pollination period (EPP) for these pairs will be approximate, what promotes mutual pollination of cultivars. The EPP is determined by the longevity of the ovule minus the time required by the pollen tube to reach the ovule [28]. The viability of the ovule and pollen, stigma receptivity and the kinetics of the pollen tube in the style tissue affect ovule fertilization, and the interaction and synchronization of these factors results in the correct fertilization effect.
Research on the number of pollen grains in anthers is not conducted frequently. This feature is very important, especially in the case of large-scale crops, where the cultivar treated as pollinating should be characterized by a large number of pollen grains in anthers. The number of pollen grains for cultivars is genetically diverse [11] but it is also influenced by weather conditions during microsporogenesis [16, 22]. Cultivars characterized by a large number of pollen grains are especially recommended as pollinating cultivars; in our experiment, the cultivars: ‘Aurora’, ‘Boreal Beast’ and ‘Sinij Utes’ exhibited this feature.
The factor determining the fertility of a flower is the ability of the stigma to accept a pollen grain, defined as stigma receptivity. The basis of stigma receptivity is the presence of active peroxidase, which is closely related to the stage of flower development [29]. The average lifespan of a haskap flower is about 5 days [26, 30]. The surface of the stigma is covered with unicellular or multicellular thin-walled papillae formed from the epidermis and the receptivity of the stigma corresponds to the integrity of papillae [31]. The stigma is capable of accepting pollen in the strictly species-defined flower development stage, hence determining stigma receptivity allows assessing the period when the stigma is ready to accept a pollen grain. Stigma receptivity changes with different stages of flower development, which was presented in our experiment. The flowers of the species Lonicera kamchatica are protogenic (protogynia), which means that the pistil develops earlier than the anthers and is able to accept pollen grains. In our experiment we distinguished five stages of flower development. Stages 1 and 2 refer to a closed bud, where in stage 2 (a closed flower with a protruding stigma), the flower begins to produce nectar and is visited by bumblebees, which are long-proboscis insects and therefore can collect nectar from the closed flowers [32]. Relating this information to different flowering times of individual cultivars, so that the stigmas of the pollinated cultivar are receptive, is an important issue. The receptivity is not active in the too early stage of flower development (stage 1) or is already disappearing in the late stage (stage 5). Therefore, the flowering time of cross-pollinating cultivars must be similar so that pollen (pollinating cultivar) and receptive stigmas (pollinated cultivar) could be available.
From pollen grains on the receptive stigma the pollen tubes germinate and grow through the style tissues, taking nutrients from them, because the materials obtained from the pollen grains are insufficient [33, 34]. Usually, the number of pollen grains on the stigma is significantly higher than the number of ovules in the ovary. In our experiment, the recorded average number of pollen grains per stigma ranged from several dozen to over 100. However, according to own unpublished data, the average number of ovules in the ovaries of the cultivars ‘Aurora’, ‘Honey Bee’, ‘Jugana’ and ‘Vostorg’ was 25.9, 28.3, 28.5 and 31.9 ovules, respectively. During microscopic observations, we found regularity in the decreasing percentage of pollen tubes overgrowing the style. The first, evident reduction in the percentage of pollen tubes occurred in 1/3 of the length of the style, then in 1/2 of the length of the style, and more than 10%of compatible pollen tubes grew to the base of pistil. According to the available literature, there are several reasons for the reduction in the number of pollen tubes growing through the style. These include the mechanical barrier, i.e. narrowing of the transmission tissue, competition between the pollen tubes and genetic barriers [35–37]. Pollen tubes differ in genetic traits and only the fastest growing pollen tubes reach the ovaries and take part in the fertilization of the ovules; this phenomenon is called pre-zygotic pollen tube selection [38]. In the sub-stigmatic part of the pistil, pollen tubes, which had the same haplotype and were recognized as incompatible, were inhibited [35]. Only genetically compatible pollen tubes grow into ovaries. In our experiment, cultivar compatibility was confirmed by the pollen germination (PGI) coefficient, which in the case of compatible pollen tubes reached a value of 2.0 or very close to it. The combinations of ‘Aurora’בJugana’, ‘Aurora’בSinij Utes’ and ‘Jugana’בAurora’ reached a PGI coefficient equal to 2, which could indicate a high compatibility of these cultivars in terms of cross pollination.
It was noted that the cultivars from the same breeding groups were the most compatible. The cultivars of Russian breeding were compatible with each other. In the case of crosses of Russian and Canadian cultivars, and vice versa, incompatibilities were observed on the stigma and in 1/3 of the length of the style. Despite a significant amount of pollen deposited on the stigma by manual deposition of pollen, only a few germinating pollen grains were observed on the stigma, in the crosses of ‘Boreal Beast’בJugana’ and ‘Boreal Beast’בSinij Utes’. In light of the experiment described above, we can conclude that there was a incompatibility between the groups of Russian and Canadian cultivars. Therefore, cultivars from these two breeding groups should not be recommended for planting for reciprocal cross-pollination on the plantation. The calculations based on microscopic observations, allowed the authors to develop a graphic model of pollen tube overgrowth through the style for three groups of crossed cultivars. For the combination - the cultivars of Russian origin crossed with the cultivars of Russian origin - the base of the pistil was overgrown by about 15%of pollen tubes in relation to the number of germinating pollen grains on the stigma. This percentage equalled on average 11 pollen tubes in the ovary, thus there was a chance to produce 11 seeds. Assuming the fact, that there are about 20–30 ovules in the ovary of haskap flowers that can potentially turn into seeds, and thus affect the fruit weight, the need for a large number of compatible pollen grains on the stigma is evident. The model shows a pollen tube outgrowth scheme for Canadian cultivars pollinated with Russian cultivars. In this combination, with a small number of pollen tubes germinating on the stigma, only 2%of pollen tubes grew to the base of the pistil (0.4 of pollen tubes on average), which meant that one pollen tube grew into the ovary only in every second flower, which did not guarantee fruit setting.
In the case of haskap the fruit should reach a weight of about 1 g for Russian cultivars and from 2.8 to 3.9 g for large-fruited cultivars from the Boreal series [7]. The minimum acceptable weight of haskap fruit is approx. 1 g, which corresponds to the number of about 6 well-formed seeds. Arising the fruit of about 2.4 g average weight requires the formation of about 25–30 well-formed seeds. The statement that fertilized ovules transform into seeds, whose number significantly affects fruit weight can be find in researches of many berry plants [39–42] as well as haskap berry [15, 44]. In our research, we calculated for the first time for haskap fruits the correlation between the number of seeds and the weight of the fruits which allowed to establish the minimum acceptable fruit weight and the minimum number of seeds to obtain this weight. We have found the occurrence of fruit without seeds in 11 combinations, especially in the crosses of cultivars of Russian and Canadian origin. The fruits were very small, of poor quality and irregular, their weight ranged from 0.2 to 0.6 g. Similar observations were recorded by [14], who obtained fruits without seeds with a weight three times smaller compared to fruits with seeds [26].
Conclusion
In the presented research, we assessed flower pollination effectiveness of nine haskap cultivars from Russian and Canadian breeding. We found that the Russian cultivars bloomed much earlier than the Canadian cultivars, and therefore the flowering time of these two cultivar groups did not fully overlap, thus most cultivars in these groups could not pollinate each other. In several cases, where the flowering time for cultivars from both breeding groups overlapped, microscopic observations revealed difficulties in the pollen tube growth through the style, which could indicate barriers of genetic incompatibility between the pollinated and pollinating cultivars. We have found that the stigma is most receptive in the freshly open flower stage and directly after the anther burst. Closed flowers with protruding stigmata were receptive in a small percentage. The quality of the forming fruits was to a large extent physiologically related to the number of seeds. We have found that the minimum qualitatively acceptable weight of a berry is 1 g, which corresponds to the formation of about 6 seeds in the fruit. The most compatible pairs of cultivars in the presented experiment, based on all the studied parameters (number of pollen grains per stigma, compatible pollen tube overgrowth through the style, PGI value of 2, large number of high-quality fruits with a large number of seeds), were: ‘Aurora’בJugana’, ‘Aurora’בSinij Utes’ and ‘Aurora’בVostorg’.
Footnotes
Acknowledgments
This work was supported by The Ministry of Science and Higher Education of Poland (project number DS-3500/KO/2019).
Funding
The authors report no funding.
Conflict of interests
The authors have declared that no conflict of interests exist.