
Abstract
BACKGROUND:
Strawberry (Fragaria×ananassa) production is hampered by anthracnose caused by Colletotrichum spp. Cinnamyl alcohol dehydrogenase (CAD) catalyzes lignin biosynthesis to fortify plant cell wall upon various stresses, but their involvement in strawberry resistance is elusive.
OBJECTIVE:
Current work aims to uncover the involvement of CAD genes in both woodland and cultivated strawberry responses to Colletotrichum spp.
METHODS:
Genome-wide identification of CAD genes was updated in cultivated strawberry. Comparative molecular characterization integrated with pathogenesis phenotyping after C. fructicola inoculation, were followed by RT-qPCR analysis and RNAseq data retrieval.
RESULTS:
Cultivated strawberry comprises 32 CAD genes of three classes. Thirteen syntenic relationships were observed between cultivated and woodland strawberry, with high homology in protein coding regions while promoters being greatly diversified. F. vesca ‘Hawaii4’ was least susceptible, followed by octoploid cultivar ‘ShenQi’, and ‘Benihoppe’ the most susceptible to C. fructicola. The class I CAD4 and CAD5, preferentially expressed in leaves and crowns, respectively, were up-regulated in susceptible varieties at 72 h post inoculation (hpi). Of class II, CAD8 was up-regulated in three genotypes at distinct timepoints, while CAD12 was quickly down-regulated in ‘ShenQi’ upon inoculation. CAD1 and CAD2 of class III were down-regulated in ‘ShenQi’ and ‘Hawaii4’ at distinct stages. The strong involvement of CAD genes in strawberry response to Colletotrichum spp. sets a basis for future functional study.
CONCLUSION:
F. vesca is less susceptible to Colletotrichum spp. than cultivated strawberries but the expression of CAD family in F. vesca and F.×ananassa were differentially provoked in timing and magnitude hinting no correlation with strawberry resistance to this pathogen.
Keywords
Introduction
Plant cell wall is the first barrier that pathogens must overcome to invade plant cells to obtain nutrients, which means that cell wall is critical for plant disease resistance. The cell wall of plants is composed of cellulose, lignin, hemicellulose, protein and pectin. Lignin is the biopolymer second to cellulose in abundance in plant [1]. Lignin is of great significance for mechanical support, water conduction and barrier to exogenous stresses. It is one of the important substances for plants to evolve from aquatic to terrestrial and adapt to terrestrial ecological environment [2, 3]. In addition, lignin is important for plant resistance to both biotic and abiotic stresses [2]. For example, lignin metabolism plays a central role in cotton disease resistance to Verticillium dahliae [4]. Lignin-based fortification of the cell wall is important for stopping Botrytis cinerea growth in strawberry fruits [5]. The lignin composition is flexible and differs among species [6]. The biosynthesis of lignin involves in a variety of enzymes, and these enzyme-coding genes are differentially expressed within the context of normal development and in responses to various stresses [6]. There exists a positive feedback regulation of lignin biosynthesis. Overexpression of wheat Cinnamyl alcohol dehydrogenase (CAD) TaCAD12 enhanced resistance via up-regulating several genes involved in monolignol biosynthesis [7].
Cinnamyl alcohol dehydrogenase (CAD) enzyme functions in the last step of synthesizing monomeric precursors of lignin, catalyzing the conversion of cinnamaldehyde (coumaryl aldehyde, coniferyl aldehyde, sinapyl aldehyde) to cinnamyl alcohol (coumaryl alcohol, coniferyl alcohol, sinapyl alcohol) through redox reaction [2, 9]. CAD protein could form heterodimers with other upstream enzymes in lignin synthesis to increase activity and efficiency [10]. CAD is encoded by a gene family composed of multiple members widely found in vascular plants [9, 11]. This family was also identified in the ancestral plants such as streptophyte algae [12] and moss Physcomitrium patens [13]. There exists a very complicated evolutionary history for CAD genes and some members are functionally versatile [15].
In Arabidopsis, there are nine CAD genes, but only AtCAD4 and AtCAD5 are considered to be critical for lignin synthesis, because double mutants Atcad4cad5 resulted in softer stems with 40% decrease in lignin content [16]. The rest seven members were thought to have a minor or no contribution to lignification [17]. AtCAD7 and AtCAD8 (previously called ELI3-1 and ELI3-2) are strongly induced by pathogens and pathogen-derived elicitors [18]. CAD genes have been studied in many agronomic plants such as sorghum [19], wheat [20], rice [21], tea [22], Populus [11, 23], melon [24], cotton [25]. The involvement of CADs in plant resistance to pathogens can be very complex. AtCAD4 and AtCAD5 were found to defend plants against Pseudomonas syringae through the salicylic acid pathway [26]. CAD7 s in Arabidopsis and tobacco were revealed as negative regulators of plant immunity induced by Phytophthora spp. infection, which were exploited by multiple Avr3a-like effectors to promote infection [27]. Silencing GhCAD35, -43, and -45 in cotton consistently damaged stem lignification and salicylate synthesis, reducing resistance to the fungal pathogen Verticillium dahliae [25].
Strawberry is a perennial herb of the Rosaceae family. It is widely grown around the world because of its unique flavor and nutritional value. The modern cultivated strawberry (Fragaria×ananassa) was an allo-octoploid interspecific hybrid between F. virginiana from North America and F. chiloensis from South America, spontaneously created in France about 300 years ago [28]. As compared with other diploid ancestors, woodland strawberry F. vesca is accepted as the single dominant subgenome provider with significantly greater gene content in cultivated strawberry [29]. Information of the improved genome sequences and annotations for F. vesca and F.×ananassa have already been available [30]. These remarkable findings provide us valuable chance to identify genes relevant to agricultural traits such as disease resistance. Virtually, strawberry is highly vulnerable to various diseases due to its short stature and soft berry fruits without protection. Anthracnose caused by Colletotrichum spp. has posed the most destructive threat to strawberry worldwide. Infection with Colletotrichum spp. can lead to the occurrence of leaf spot, crown rot, stolon spot, petiole spot, and fruit rot in strawberry [32]. Currently, C. siamense and C. fructicola of C. gloeosporioides complex are the most prevailing species threating strawberry worldwide [34]. Anthracnose management in strawberry field relies heavily on fungicides, but drug resistance has widely evolved in these pathogenic fungi [38]. Therefore, it is of great significance to identify strawberry genes relevant to defense for future resistance breeding.
Previously, we had performed a genome-wide identification of CAD genes in F. vesca and F.×ananassa [39] prior to the release of the improved genome annotation for cultivated strawberry [31]. There is still some uncertainty regarding the CAD gene family in cultivated strawberry. Furthermore, it is short of an analysis of this versatile family in response to the most destructive pathogen Colletotrichum spp. of strawberry. Thus, current work aims to i) update CAD family in cultivated strawberry based on the recently improved genome annotation, ii) comparatively characterize this family in cultivated strawberry and its dominant ancestor F. vesca, iii) identify the differentially regulated CAD genes in both woodland and cultivated strawberry infected with Colletotrichum spp. This study provides a systematic characterization of strawberry CAD gene family, which would set a basis for future functional study of strawberry CAD genes and anthracnose pathogen-response mechanisms in strawberry.
Materials and methods
Plant materials, growth conditions, and inoculation with Colletotrichum fructicola
The woodland strawberry ‘Hawaii4’ (Fragaria vesca) and cultivated strawberry (F.×ananassa) varieties ‘Benihoppe’ and ‘ShenQi’ were grown in an incubator at constant 25°C under a 12h-dark/ 12h-light rhythm. Healthy plants with 6-8 fully expanded leaved were used for inoculation with a hemibiotrophic fungal pathogen C. fructicola (CGMCC3.17371, a strain from strawberry field in Qingpu District, Shanghai, 2007). The preparation of inoculum and spraying-inoculation method for whole plant was previously reported [40]. To observe the disease symptoms, 10 plants for each variety were used in one time, and 20 ml conidia solution (2×106 cfu) with 0.01% Tween-20 was sprayed for one plant. The experiment was repeated twice independently in 2019. Anthracnose symptoms on whole plants were scored at 4 days post inoculation (dpi) and 7 dpi. Leaves with anthracnose lesions were rated in a zero to nine scale as previously reported [41]. Disease index (%) was calculated following the same publication.
For expression analysis, thirty-six plants of each variety at the same developmental status were used for inoculation in 2021. A half of plants were sprayed with conidia solution and the rest were sprayed with sterile water containing 0.01% Tween 20 for mock-treatment. Eighteen plants were divided into three biological replicates. Six leaves (the third or the fourth) collected from six independent strawberry plants at 0, 6, 24, 48 or 96 hours post inoculation (hpi) were pooled as a biological replicate. It was noteworthy that 0 hpi was actually corresponding to less than 30 min post inoculation, because sampling was performed after spraying all 18 plants of one genotype with 20 ml conidia solution per plant. Sampled leaves were wrapped in tin foil and stored immediately at -80°C for RNA purification in one month. The experiment was repeated twice independently.
Identification and phylogenic analysis of CAD genes in Fragaria×ananassa
To identify CAD genes in octoploid cultivated strawberry, we tried two approaches. First, the mRNA sequences of 14 FveCAD genes of Fragaria vesca [39] were used for BLASTN analysis against the F.×ananassa CDS database v1.0.a2 at https://www.strawberryblast.ml:8080/strawberry/viroblast.php (SGD) [31]. Alternately, the conserved ADH_N (Alcohol dehydrogenase GroES-like domain) and ADH_ZINC_N (Zinc-binding dehydrogenase) domains of Arabidopsis CAD proteins were used for BLASTN search against the GDR database (Fragaria×ananassa genome v1.0 a2 transcripts). Candidate sequences with a E value < 1E-5 were selected and further validated at PFAM website (https://www.ebi.ac.uk/Tools/pfa/pfamscan/) for the occurrence of both ADH_N and ADH_ZINC_N domains. The same results were obtained in two approaches and a total of 32 CAD loci was identified in the genome of cultivated strawberry.
Phylogeny analysis of CAD proteins from woodland and cultivated strawberry was achieved in the software MEGA7.0 using the neighbor-joining (NJ) method with 1000 bootstrap replicates. The resulted evolutionary tree was improved using the iTOL online tool (https://itol.embl.de). Finally, the F.×ananassa CAD genes were named according to their phylogenetic relationship with homoeologous allele in F. vesca.
In silico prediction of the biochemical features for the deduced F.×ananassa CAD proteins were accomplished online: subcellular localization predicted at the WoLF PSORT website (https://wolfpsort.hgc.jp/), and isoelectric points(pI) and molecular weights (Mw) analyzed at the ExPASy website (https://web.expasy.org/).
Exon-intron structure, protein module and sequence alignment
The exon-intron structure of cultivated strawberry CADs was obtained via comparing CDS and genomic DNA sequence for each member using the GSDS online tool (https://gsds.cbi.pku.edu.cn/). The conserved ADH_N and ADH_zinC_N domains in CAD protein sequences was analyzed in PFAM and visualized using IBS software. In addition, conserved motif information for strawberry CADs was identified at MEME website (https://meme-suite.org/) and displayed via using TBtools [42].
For the alignment of protein sequences, ClustalW online tool (https://www.genome.jp/tools-bin/clustalw) was used for the CAD members from F.×ananassa, F. vesca and Arabidopsis thaliana. The result of sequence comparison from ClustalW saved as.aln format was further mapped via using the online tool ENDscript/ESPript (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi) [43]. The final output image was improved in Photoshop.
Chromosomal localization and synteny analysis
The chromosomal position information of strawberry CAD genes was obtained from genome databases at GDR [29]. The physical map showing all chromosomes of F.×ananassa and the distribution of CADs was visualized by MapChart1.0 [44].
To find synteny between F.×ananassa and F. vesca, genome databases with annotations were downloaded from SGD in.gff3 format, and the synteny was calculated using the MCscan pipeline [45] in the software J. Craig Venter Institute (JCVI)-Syn 1.2.7 (https://github.com/tanghaibao/jcvi) (Python-version). In JCVI the synteny between two genomes were calculated with a hit score reaching 0.99, which was used to effectively screen out the best hits. The resulted synteny relationships were drawn and simplified with the synteny for CAD loci highlighted. The synteny for CADs between F. vesca and F.×ananassa was also summarized into a table, which was improved in Photoshop.
Identification of cis-elements for promoter sequence
The genome databases of F. vesca and F×ananassa in .gff3 format were downloaded from SGD, which were uploaded into TBtools and the 2500 bp upstream fragment before the start codon (ATG) was extracted as the promoter sequence for each CAD gene. The promoter sequence was submitted to plantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html) to identify cis-elements. The prediction information was classified and visualized via using TBtools package.
Primer design, RNA extraction and RT-qPCR
For RT-qPCR analysis, specific primerset matching each FveCAD in F. vesca and its homoeologous alleles in F.×ananassa was designed using Primer 5.0. Selected primers (supplementary Table S1) had also been examined via BLASTN analysis in strawberry genome database [31] (SGD, https://www.strawberryblast.ml:8080/strawberry/viroblast.php) to ensure the specificity. In addition, a mixed cDNA template of ‘Benihoppe’ leaves inoculated with C. fructicola and mock treated at different stages, was used for primer efficiency analysis as diluted templates (3×, 9×, 27×and 81×).
RNA isolation and RT-qPCR were performed as previously described [46]. Briefly, the ChamQTM Universal SYBR qPCR Master Mix (Vazyme, Lot#Q711, Nanjing, China) was used in a total reaction volume 12μl run in Light Cycler 480 (Roche, USA). RT-qPCR analysis was performed using the 2-ΔΔCT method combined with a calibration in geNorm software based on the Ct values of two reference genes EF1a and GAPDH2 [47].
Retrieval of RNA-seq data for strawberry responses to Colletotrichum spp.
To further dissect the expression profiles of strawberry CAD genes upon infection with Colletotrichum spp., relevant RNA-seq data in previous reports [48] were mined. The conditions for strawberry-Colletotrichum spp. interaction and reference databases used in those previous works were summarized hereafter to enhance comparing the expression data obtained.
In Wang et al. [48], strawberry varieties ‘Yanli’ (resistant) and ‘Benihoppe’ (susceptible) were inoculated with C. gloeosporioides; RNA sequencing was performed for leaves sampled at 0 h, 72 h untreated and 72 hpi in two varieties, where the database Fragaria×ananassa (FAN_r1.1) (https://strawberry-garden.kazusa.or.jp/) was used as the reference for annotation. In Zhang et al. [40] of current lab, a susceptible strawberry cultivar ‘JiuXiang’ was inoculated with C. fructicola and leaves at 0, 24, 72 and 96 hpi were sampled for RNA-seq, where the genome of woodland strawberry (https://www.rosaceae.org/species/fragaria/fragaria_vesca/genome_v1.0) were used as reference.
In Chandra et al. [49], two cultivated strawberry cultivars ‘Florida Elyana’ (resistant) and ‘Festival’ (susceptible) were inoculated with C. gloeosporioides on crowns, where crowns were sampled at 72 hpi, and the GDR database (Fxa_v1.2_makerStandard_transcripts) was used as reference for annotating RNA-seq data. In Adhikari et al. [50], an expression study was performed in the leaves of a susceptible genotype ‘Chandler’ as compared with in a resistant genotype NCS 10-147 at 48 hpi with C. gloeosporioides, where the reference genome database was the recently updated [31].
Statistical analysis
The average of three biological replicates (each examined in three technical repeats, a total of 9 raw data) was used for expression values. For transcriptional differences larger than 2-fold between average values, statistical analysis was further performed. The independent sample T-test method in SPSS software was used to detect whether there was significant difference between the mock-treated and the inoculated material at each timepoint, with P≤0.05 (*), 0.01 (**) or 0.001 (***). The Waller-Duncan method in one-way ANOVA test was used to detect significantly dynamic changes in the transcript levels of CADs during certain biological process. Different lowercase and uppercase letters were used to indicate the significant dynamic changes in mock and inoculation-treatment, respectively. Finally, diagrams were produced in Origin2022 software.
Results
Characterization and phylogeny of CAD genes in cultivated strawberry
Based on BLAST search and PFAM analysis, a total of 32 CAD genes were identified in the updated database of F×ananassa CDS v1.0 a2 (Table 1). It was worth noting that two loci FxaC_6g04900 and FxaC_6g04990 shared completely identical sequences in 2500 bp upstream promoter, gene and protein. The possibility of incorrect sequence assemble cannot be ruled out. Thus, FxaC_6g04990 was not included for further characterization in current work. The whole CAD family genes of cultivated strawberry were clearly grouped into three classes through phylogenetic analysis, with 9 loci of Class I, 19 of Class II and 4 of Class III (Fig. 1). Accordingly, these genes were named after their homologs in Arabidopsis and F. vesca. There were one to three FxaCAD homoalleles corresponding to one FveCAD of Class II and Class III, while four to five FxaCAD homoalleles were identified for FveCAD of Class I, the bona fide CAD. The preferential subcellular locations for FxaCADs were predicted firstly to be cytoplasm (Ch), secondly cytoskeleton (Ck). However, FxaCAD3a/b, FxaCAD13a and FxaCAD4d were predicted to localize at peroxisome (Pe), chloroplast (Ch) and nucleus/extracellular matrix (Nu/Ex), respectively. The molecular masses of FxaCADs ranged from 30.08 kDa (FxaCAD14a, 275aa) to 47.58 kDa (FxaCAD13a, 437aa). The isoelectric point ranged from 5.22 (FxaCAD4a) to 8.0 (FxaCAD13a), and most members of Class I with isoelectric points lower than 6.0 (Table 1).
The genomic and biochemical information for CAD genes in F.×ananassa genome
The genomic and biochemical information for CAD genes in F.×ananassa genome
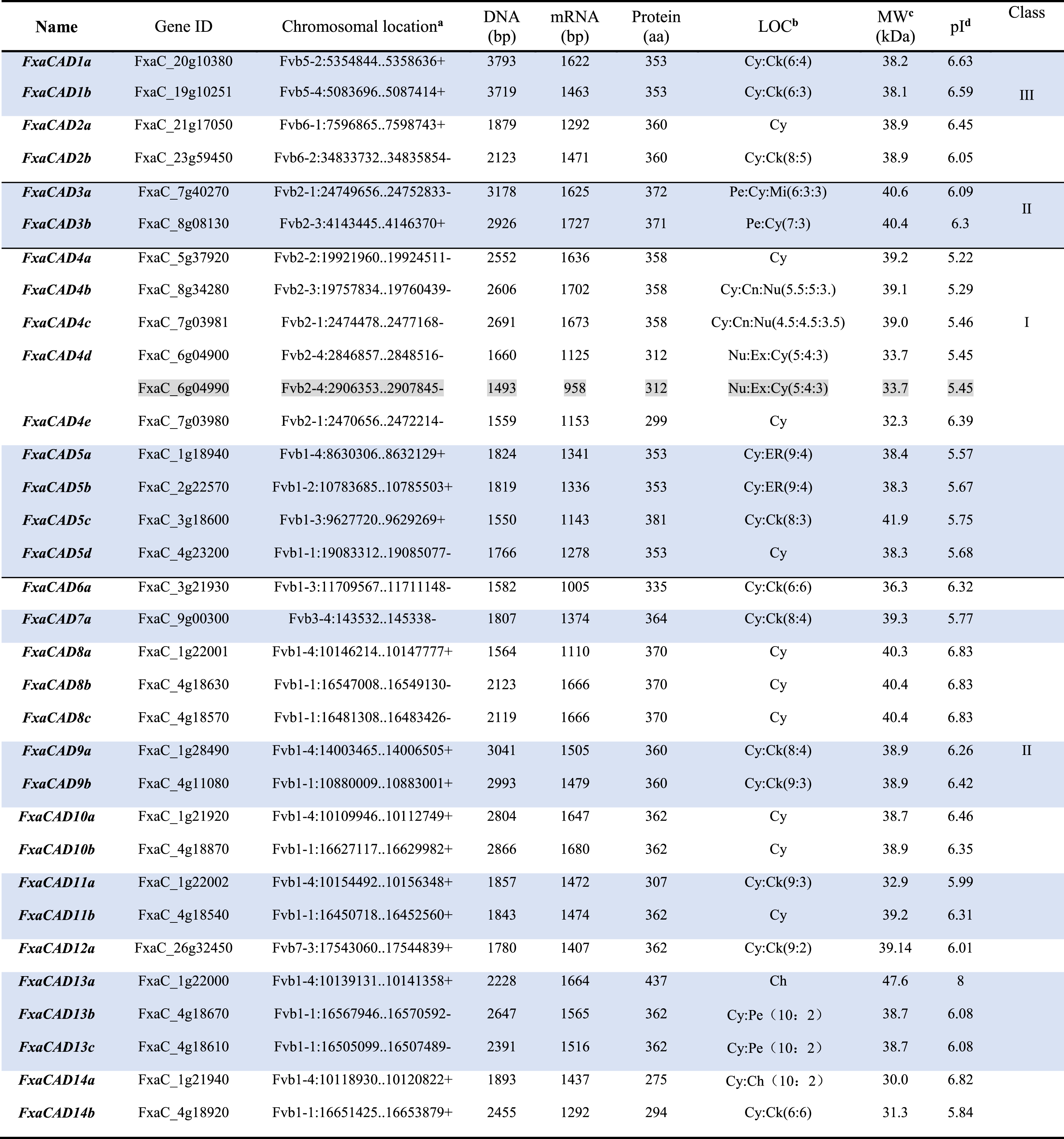
a“+, - ” for transcription orientation. bSubcellular locations predicted at the WoLF PSORT website: Ch (chloroplast); Ck (cytoskeleton); Cn (shuttling between the cytoplasm and nucleus); Cy (cytoplasm); ER (endoplasmic reticulum); Ex (extracellular matrix); Mi (mitochondria); Nu (nucleus); Pe (peroxisome). cMolecular weight of the amino acid sequence, Da is Daltons. dIsoelectric point.

Phylogenetic relationships of cinnamyl alcohol dehydrogenase (CAD) proteins in strawberry. A set of 55 CAD proteins including 32 FxaCADs (highlighted in bold) from https://www.strawberryblast.ml:8080/strawberry/viroblast.php (SGD), nine Arabidopsis AtCADs from www.arabidopsis.org (TAIR), as well as 14 FveCADs (gray-shaded) from https://www.rosaceae.org/ (GDR) were included for constructing the neighbor-joining (NJ) tree using MEGA7 software with 1000 bootstrap re-samplings and visualized using the interactive Tree Of Life (iTOL). Different colored rings indicate distinct classes.
Exploring gene structure revealed that the intron-exon organization of CAD family genes was largely conserved among Arabidopsis, F. vesca and F.×ananassa (supplementary Fig. S1). Genes of Class I possessed 5 exons with the exception of FxaCAD4d and -4e missing one exon. Most genes of Class II had 5 exons except for FxaCAD14a and -14b being composed of 3 exons. As compared with FveCAD11 and FxaCAD11b, FxaCAD11a also missed one exon. Comparative analysis of the protein module structure indicated that most FxaCADs possessed the conserved ADH_N (Alcohol dehydrogenase GroES-like domain) and ADH_ZINC_N domains. Most FxaCADs showed identical module organization as their homoalleles in F. vesca (supplementary Fig. S2). However, truncated ADH_N domains were found in FxaCAD11a and -4e. Also, incomplete ADH_ZINC_N domains were observed in FxaCAD6 and -14a. These differences could be further evidenced in motif composition revealed via using MEME online tool (supplementary Fig. S3).
Sequence alignment of the CAD proteins from Arabidopsis, F. vesca and F.×ananassa revealed that their ADH_N and ADH_zinc_N domains were composed of four highly conserved functional motifs (supplementary Fig. S4): Zn1 catalytic center (C47, H69, and C163, referring to the sites in FxaCAD4a), the Zn-binding signature GHEXXGXXXXXGXXV (starting with G68 in FxaCAD4a), the Zn2 structural motif (C100, C103, C106, and C114 in FxaCAD4a), and the NADPH-binding domain [GLGGV(L)G] motif (starting with G189 in FxaCAD4a), similarly organized in Arabidopsis [51]. All these observations indicated that FxaCAD members identified in current study belong to plant CAD family.
Two contrasting insights emerged from analyzing the chromosomal localizations of CAD genes in F.×ananassa. FxaCAD4 s (on Fvb2) and FxaCAD5 s (on Fvb1) of the bona fide Class I were evenly distributed on four subgenomes (Fig. 2a), which reinforced the evolutionary conservation and importance of these suggested authentic genes for lignin biosynthesis [9]. However, the rest FxaCAD genes were unevenly distributed. Total 32 CAD genes identified in F.×an anassa were composed of 10, 3, 7 and 12 loci in the A, B, C and D subgenomes, respectively. A tandem duplication event involving six FveCADs of Class II (FveCAD10, -14, -13, -8, -11 and -6) on LG1 of F. vesca genome [39] largely preserved and duplicated in A and D subgenomes of F.×ananassa, with the exception of CAD6 experienced an interchromosomal translocation to Fvb1-3 of C subgenome (Fig. 2a). The rest five genes in this tandem repeat were intactly maintained on Fvb1-4 of A subgenome (FxaCAD10a, -14a, -13a, -8a and -11a) and on Fvb1-1 of D subgenome (FxaCAD14b, -10b, -13b, -8b, -13c, -8c and -11b). In the latter, both chiasmata (the relative locations of FxaCAD14 and -10) and biased duplication (FxaCAD8b/c and -13b/c) might have contributed to the expansion of this tandem repeat in cultivated strawberry.
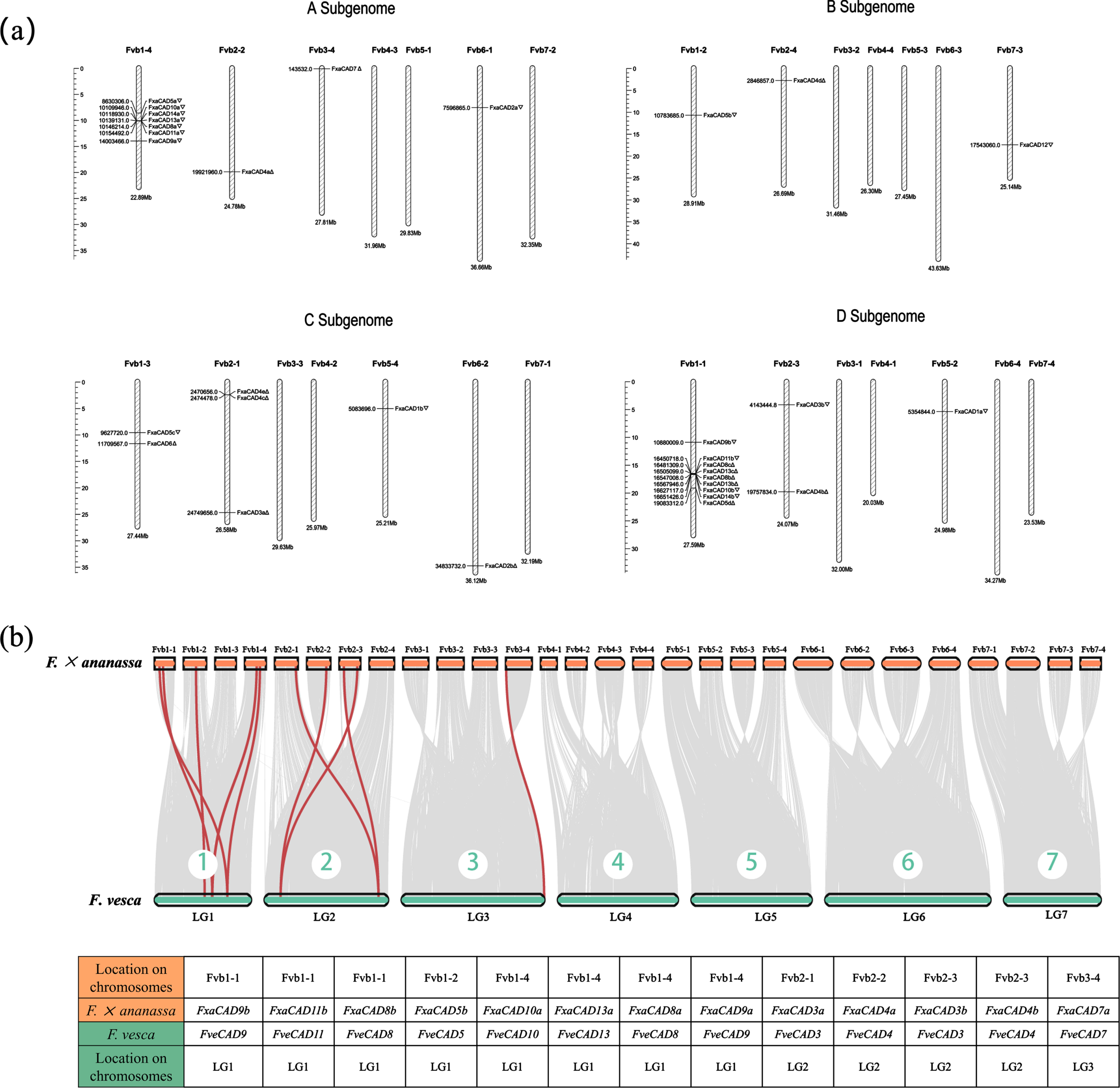
Chromosomal distribution and synteny of FxaCADs with FveCADs.
To explore the orthologous relationships of CAD genes between cultivated strawberry and its ancestor woodland strawberry, the synteny between the whole genomes of F.×ananassa and F. vesca was calculated. With a hit score set to 0.99, in current work 13 syntenic gene pairs were found between the CAD families of F.×ananassa and F. vesca, with eight on LG1, four on LG2 and one on LG3 (Fig. 2b). Moreover, there were four FveCADs (FveCAD3, -4, -8 and -9) each corresponding to two syntenic genes in F.×ananassa chromosomes with 99% or above identity. These syntenic genes might share high similarity in both molecular structure and biological function. In sum, nine of 14 FveCADs including the bona fide Class I have intactly persisted and even been duplicated in cultivated strawberry through multispecies merge and domestication.
The occurrence of cis-elements in promoter often sheds light on the biological process certain gene involved in. A total of 458 and 683 cis-elements were identified in the 2500 bp upstream before the starting code ATG of CAD family genes in F. vesca and F.×ananassa, respectively (supplementary Table S2). The elements related with plant growth accounted for the lowest proportion than those related with plant responses to hormones and stresses. The growth-related cis-elements of FveCAD were often lost in the promoters of some homoeologous members in F.×ananassa (Fig. 3).
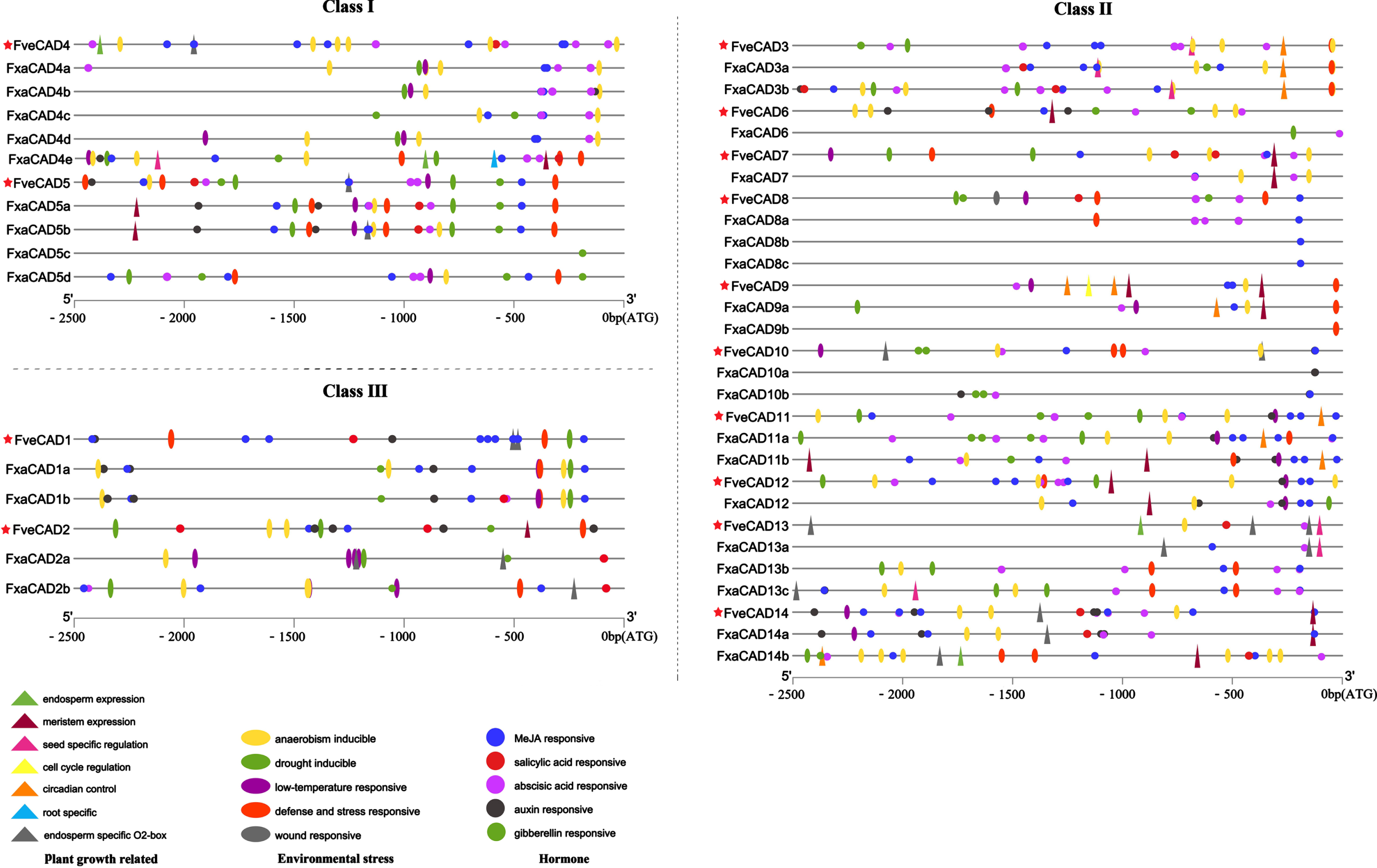
Cis-elements in the promoter regions of CAD genes in F. vesca and F.×ananassa predicted at PlantCare. Different colors and shapes were used to distinguish different cis-elements, with triangles for plant growth-associated, ovals for environmental stresses, and circles for hormone responsiveness. Information for the CAD genes from F. vesca was marked with red star. The promoters of F.×ananassa and F. vesca CAD genes harbor too many light responsive elements to be included here, since which might influence the visuality of other cis-elements.
In both F. vesca and F.×ananassa, the promoters of CAD genes had above 40% cis-elements related with hormonal responses. The most abundant and ubiquitous cis-elements were MeJA responsive motifs (blue circle). There were 9 copies of this motif identified in the promoter of FveCAD1, and a total of 7 copies were observed in the promoters of its cultivated strawberry homoalleles FxaCAD1a and -b. The abundance of Abscisic acid (ABA) responsive element ranked the second in strawberry CAD promoters. Interestingly, this element was largely absent in the promoters of CAD genes belonging to Class III. By contrast, the least identified element involved in hormone responses was salicylic acid (SA) response-related (red circle). Several members contained one copy of SA-responsive element, and only the promoters of FveCAD2, -7, and FxaCAD3b contained 2 copies.
Environmental stress responsive elements were clearly enriched in the promoters of CAD genes of cultivated strawberry than those of F. vesca. Notably, the promoters of FveCAD5 together with its homoalleles FxaCAD5a and -b similarly contained 3 copies of the defense and stress responsive element (TC-rich repeats, red oval). Although none of this type element was identified in FveCAD11 promoter, FxaCAD11a and -b each possessed one copy of TC-rich repeats in their promoter. Generally, in terms of the composition of cis-elements in CAD promoters, variations were widely observed between two strawberry species F. vesca and F.×ananassa.
Pathogenesis following spraying-inoculated C. fructicola challenge in three genotypes was monitored with the occurrence of necrotic lesions on leaf blades. Disease lesions were hardly observed in ‘Hawaii4’ at 4 dpi, when lots of diseased spots were clearly observed on the inner young leaflets of two octoploid varieties (Fig. 4a). In the leaflets of the cultivar ‘Benihoppe’, the formation of necrotic lesions was higher than that in ‘ShenQi’ at 4 dpi. The variation in the susceptibility was further distinguished at 7 dpi (Fig. 4b). Typical symptoms occurred on leaves, varying with the phyllotaxy. The necrosis in the first leaf was more serious than that of older leaves. The first and second leaves of ‘Benihoppe’ even were withered at leaf edge, while the disease symptom in ‘ShenQi’ was lighter with necrotic lesions largely limited to two inner leaves. At this timepoint, very small necrotic lesions could also be observed on the first to third compound leaves of ‘Hawaii4’.

Differences in the susceptibility to Colletotrichum fructicola among F. vesca var. ‘Hawaii4’, F.×ananassa var. ‘Benihoppe’ and ‘ShenQi’.
The percentage of necrotic leaf area was calculated to compare their susceptibility to C. fructicola in three strawberry varieties (Fig. 4c). Low necrotic lesions developed in ‘Hawaii4’, and the disease index was highest in cultivated variety ‘Benihoppe’ at any timepoint. Although F. vesca ‘Hawaii4’ plant was smaller in stature with thinner leaf blades than F.×ananassa, it was less susceptible than herein two cultivated genotypes. ‘Benihoppe’ was highly susceptible, and ‘ShenQi’ was moderately susceptible.
RT-qPCR based transcription assay was used to identify defense-related CAD genes of strawberry in response to C. fructicola infection. The expression responses during 96 hpi were investigated in aforementioned three genotypes with varying susceptibility. PCR amplification with some primers specific to several genes resulted in low or unevaluable PCR efficiency [52]. Those unevaluable members included FveCAD5-7/FxaCAD5-7, which were completely not detected in current work, as well as FveCAD14/FxaCAD14, whose transcripts had been detected but the level was too low to be reliably quantified. In addition, primer PCR efficiency for FveCAD2/FxaCAD2 was lower than 80% (Table S1), whose expression data displayed in Fig. 5 should be refereed with cautious.

RT-qPCR analysis of strawberry CAD genes upon C. fructicola infection in three strawberry varieties. The relative expression of CAD genes in F.×ananassa and F. vesca was normalized with two reference genes GAPDH2 and EF1 α (Amil-Ruiz et al. 2013) [47]. CK for mock-treated; Cf for C. fructicola infection. Significant differences between CK and Cf at certain time point were indicated at the 0.05 (*) or 0.01 (**) probability level (Student t-test). Uppercase and lowercase letters marked over the columns indicate significant changes (p < 0.05) during the dynamic process for Cf and CK, respectively. When there was no significant difference, letters were omitted.
As shown in Fig. 5abc, FveCAD4/FxaCAD4 was the only member of Class I bona fide CAD detected. The transcriptional level of this gene was not differentially altered during 96 hours with C. fructicola infection in the less susceptible F. vesca ‘Hawaii4’ and the moderately susceptible F.×ananassa ‘ShenQi’, contrasting with a clear upregulation in the highly susceptible genotype ‘Benihoppe’ at 96 hpi. Of Class III CADs, FxaCAD1 was only significantly downregulated at 48 hpi in ‘ShenQi’, and it might be transiently induced at 6 hpi in two octoploid genotypes although not to a significant level. FveCAD2 was markedly down-regulated in F. vesca at 24 h and 96 h post C. fructicola inoculation, whereas its homoallele FxaCAD2 was not significantly changed in two octoploid genotypes.
Among the seven members of Class II CADs, the transcription levels of FveCAD9-10/FxaCAD9-10 and FveCAD13/FxaCAD13 were not significantly altered in all three genotypes during 96 h post C. fructicola inoculation. In each genotype, FveCAD12/FxaCAD12 showed an expression pattern reminiscent with that of FveCAD11/FxaCAD11. In F. vesca, although not to a significant level, FveCAD11-12 might be transiently upregulated immediately after C. fructicola inoculation (indicated as 0 hpi, less than 30 min post inoculation). In the highly susceptible ‘Benihoppe’, FxaCAD11 was significantly upregulated at 6 hpi, and FxaCAD12 might be also induced at this timepoint although not to a significant level. Contrastingly, in the moderately susceptible ‘ShenQi’, FxaCAD12 was markedly down-regulated immediately after C. fructicola inoculation at 0 hpi. Suppression of FxaCAD12 lasted till 6 hpi although not to a significant level, and there might also be a weak downregulation of FxaCAD11 at the early stages post inoculation with C. fructicola. In most cases, FveCAD3/FxaCAD3 showed a pattern similar with that of Class I FveCAD4/FxaCAD4, i.e., not significantly altered in ‘ShenQi’, downregulated at 96 hpi in ‘Benihoppe’. But only FveCAD3 was markedly up-regulated immediately upon C. fructicola inoculation within 30 min. It was noteworthy that FveCAD8/FxaCAD8 was the unique member differentially fluctuated during 96 h post C. fructicola inoculation in all three genotypes, significantly upregulated at distinct timepoints. It was markedly induced at 0 hpi and 96 hpi in ‘Hawaii4’, while induced at 6 hpi and 48 hpi in ‘Benihoppe’ and ‘ShenQi’, respectively.
Generally, half members of strawberry CAD family (7/14) involved in response to C. fructicola infection. All these seven members were differentially regulated in three genotypes with varying susceptibility. Temporally, significant expressional overlap during 96 h post C. fructicola inoculation was observed for the CAD homologues. For example, FveCAD3 and FveCAD8 were immediately induced at 0 hpi in F. vesca. FxaCAD8 and -11 were induced at 6 hpi as well as FxaCAD3 and -4 were up-regulated at 96 hpi in ‘Benihoppe’. Significant early down-regulation of CAD gene was only observed for FxaCAD12 in ‘ShenQi’ upon C. fructicola inoculation.
To expand our understanding of strawberry CAD family involved in defense responses, RNAseq data in cultivated strawberry-Colletotrichum spp. were retrieved from previous publications [40]. BLAST search against the nucleotide acid sequence databases previous reports used in strawberry Garden and GDR was performed to match members sharing higher than 90% identity with current CADs. If the previously reported sequence matched several members of current work, the expression value was assigned to the member(s) with the highest sequence identity. Still some sequence obtained in RNA sequencing could be completely identical with one partial fragment conserved in several homologues.
In a comparative transcriptomic assay [48], the differentially expressed CADs between the highly susceptible ‘Benihoppe’ and resistant ‘Yanli’ at 72 hpi with C. gloeosporioides were obtained (Fig. 6a). A transcript fragment matching FxaCAD4abc showed transcription increase in the resistant ‘Yanli’ as compared with no change in the susceptible ‘Benihoppe’ at 72 hpi with C. gloeosporioides, while transcripts matching FxaCAD4de were similarly down-regulated in two genotypes with varying susceptibility. Transcripts matching FxaCAD1ab and FxaCAD2ab were upregulated in the resistant genotype ‘Yanli’, and the later was also downregulated in the susceptible ‘Benihoppe’ at 72 hpi. With the exception of FxaCAD9, several members of Class II CADs were down-regulated in the resistant ‘Yanli’, i.e., FxaCAD3ab, -7a, -10ab, -11ab/12a and -13abc at 72 hpi, while FxaCAD10ab, -11ab/12a and -13abc were also down regulated in the susceptible ‘Benihoppe’ to varying degree.
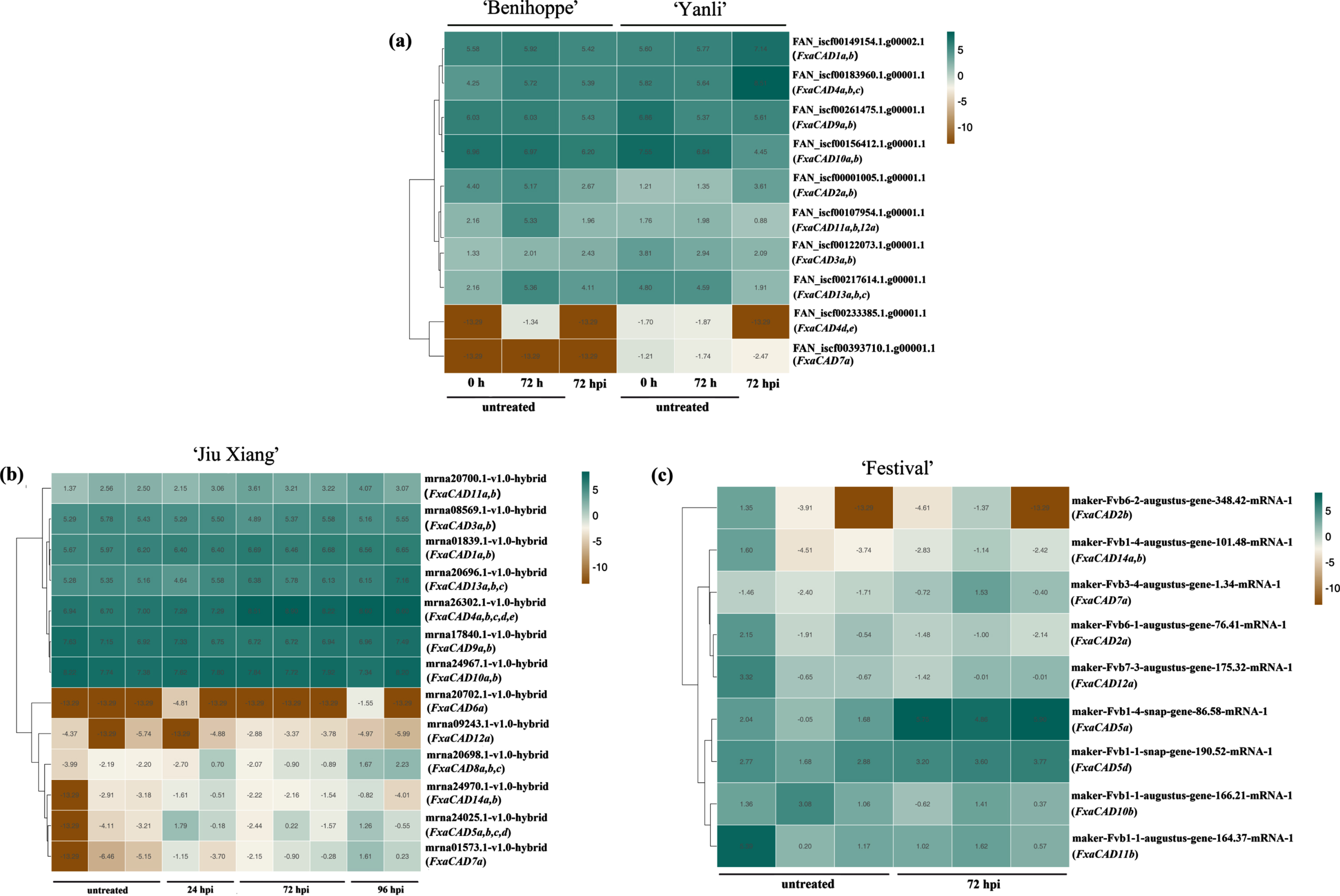
RNA-seq data retrieved from previous studies for CAD family genes in cultivated strawberries upon Colletotrichum spp. infection. The RPKM (Reads Per Kilobase per Million mapped reads) values were normalized after Log2-transformation for plotting. Heatmaps were generated via using the Pasenol Gene Cloud online tool (https://www.genescloud.cn/chart/HeatMap). Strawberry materials and fungal pathogen inoculation conditions for three panels were reported previously:
In our previous work [40] aiming at revealing the temporal transcription changes in a moderate susceptible genotype ‘Jiuxiang’ infected with the same C. fructicola strain, seven CADs including FxaCAD4, -5, -7, -8, -11, -12 and -13 displayed transcriptional increase upon infection at different timepoints than in mock-treated plants at 0 hpi (Fig. 6b). An additional comparative study of the defense responses in the crown tissues of susceptible ‘Festival’ and resistant ‘Florida Elyana’ was performed with C. gloeosporioides infection [49]. That work revealed a clear upregulation of FxaCAD5a, -5d and -7a in the crowns of ‘Festival’ at 72 hpi, while no differentially expressed CAD members were identified in ‘Florida Elyana’ (Fig. 6c). In an additional work, FxaCAD13a (strawberryCLC_DN25_c9461_g9461) was significantly down regulated in the leaves of a susceptible genotype ‘Chandler’ as compared with in a resistant genotype NCS 10-147 at 48 h post infection with C. gloeosporioides, which was not shown in Fig. 6.
The above high-throughput sequencing from RNAseq analysis largely disagreed with our RT-qPCR results, highlighting the complexity of strawberry CAD family responses to the infection with Colletotrichum spp. The probable reasons might include differences in strawberry genotypes, strawberry tissues, the inoculation conditions and the fungal strains. It was worth mentioning that CAD4/5 members belonging to the Class I branch were the most highly expressed members at 72 hpi post Colletotrichum spp. infection, with FxaCAD4 and FxaCAD5 markedly up-regulated in leaves and crowns, respectively.
The formation of the octoploid strawberry was proposed approximately one million years ago, whose chromosomes are indeed mosaics of DNA from four different diploid ancestors following widespread homoeologous exchange, biased fractionation and selective sweep [28]. As the spontaneous hybrids of wild octoploid species F. chiloensis and F. virginiana about 300 years ago, cultivated strawberries worldwide have experienced repeated expansion and reshaping in genetic diversity through introgression of alleles from wild allo-octoploid founder ecotypes during domestication [53].
Our previous work revealed that F. vesca genome holds 14 CAD loci and most FveCADs have at least one homoallele sharing ∼97% or above amino acid identity in cultivated strawberry [39]. In this study, 32 CAD genes were identified in the genome of cultivated strawberry F.×ananassa. The unevenly distribution of CADs in the genome of F.×an anassa with more members (22/32, about 69%) on A and D subgenomes is agreeing with a previous report that the unique dominant diploid ancestor F. vesca contributed 74.2-81.4% and 52.7-66.2% of the genic DNA on A and D subgenomes, respectively [28]. Clearly, CADs from the ancestor F. vesca are largely maintained and even duplicated in cultivated strawberry. The high homology relationship among these CADs implies similar functions between two species. However, the occurrence of cis-elements in the promoters of FveCAD and its FxaCAD homoalleles is largely distinct, with the exception of that between FveCAD14 and FxaCAD14a, which is quite different from the conservation observed for the promoters of NPR-like families from cultivated strawberry and F. vesca [46]. Upon inoculation with Colletotrichum spp., transcription of several CAD members was significantly altered in F. vesca, F.×ananassa or both. Notably, none of these differentially expressed genes displayed a similar pattern in three genotypes including F. vesca and two F.×ananassa varieties varying with susceptibility. Indeed, it is hard to correlate expression of any CAD gene with strawberry resistance to Colletotrichum spp. at present. In addition, our results showed that F. vesca var. ‘Hawaii4’ is less susceptible than two octoploid cultivars, although with thinnest leaves and smallest stature. This is consistent with a previous report [54]. Clearly, wild strawberry species comprise many natural sources of disease resistance and are valuable resources for breeding more resistant cultivated strawberry [55].
Lignin is an amorphous polymer component of plant secondary cell wall, and there exist at least 35 natural lignin monomers [6]. Cinnamyl alcohol dehydrogenase (CAD) catalyzes the reduction of multiple substrates to generate the diverse lignin monomers in the last step [56]. Naturally, lignin amount and composition vary greatly, depending on plant species, tissues, cell types, cell wall layers, as well as developmental status, environmental factors and stress types. This might be the same scene for CAD gene expression. In Arabidopsis, AtCAD5 was specifically expressed in the more lignified roots at a high level [57]. Although we did not detect the expression of FveCAD5 and its homoalleles in fruits [39] and leaves (current), RNAseq data indicated that woodland strawberry FveCAD5 was preferentially expressed in root and remarkably up-regulated upon phosphate starvation (4.8-fold increase) [30]. Interestingly, FxaCAD5 transcripts were dramatically enhanced in the susceptible strawberries upon infection with Colletotrichum spp. [49]. In flax (Linum usitatissimum), the class I LuCAD1B was markedly induced in root tips at 48 h post infection with Fusarium oxysporum [58]. It seems that the class I bona fide CADs not only are capable of responding to various stresses, but have tissue-specific expression patterns conserved in some dicot species. The observations in strawberry reinforced that CAD4 and CAD5, the most highly expressed CAD member in the aboveground and underground parts, respectively, might cooperate but not function redundantly with each other in lignin synthesis and against stresses at distinct parts.
It was suggested that AtCAD7 subfamily has lost the enzymatic activity of canonical CAD members, but gained a novel role as a negative regulator of plant defense against Phytophthora capsici [27]. This might be partially controversial. Indeed, AtCAD7 does not completely loses enzymatic potential but still has a low activity in catalyzing the biosynthesis of monolignol [51]. The enzymatic activity and responsiveness to stresses might be largely conserved in CAD genes. Although there is short of solid evidence for the occurrence of lignin in the ancestral moss plant Physcomitrium patens, overexpression of PpCAD1 resulted in more giant moss colony and enhanced resistance to Botrytis cinerea; PpCAD1 ectopic overexpression in Arabidopsis increased lignin content and resistance against pathogens [13]. In upland cotton, there were three genes including GhCAD35 (belonging to class I bona fide CAD), GhCAD45 (within the same branch of AtCAD7 of class II), and GhCAD43 (CAD-like class V) consistently induced by infection with the soil-borne fungus Verticillium dahlia. Silencing each of these three CADs reduced defense-induced lignification and impaired resistance to V. dahlia [25]. Knocking out the AtCAD8 paralog OsCAD8B in rice enhanced susceptibility to Rhizoctonia solani, while inhibiting the paralog ZmCAD degradation in maize reduced plant lignin accumulation and susceptibility to R. solani. Thus, these class II CAD genes are important for lignin accumulation to restrict the infection of R. solani [59].
The present study indicates that strawberry CAD genes were differentially provoked in response to Colletotrichum spp. Whether these genes contribute to resistance or susceptibility in the studied diploid and octoploid genotypes is questionable. Notable differences in the timing and magnitude of strawberry CADs expression responses to C. fructicola were revealed. Concerning the metabolic complexity of lignin [60], the complicated evolution of CAD family [15], and the fact that lignin content and CAD expression are not always positively correlated with plant disease resistance [61], it is hard to infer the function of CAD genes responsive to Colletotrichum spp. Previously, significant positive correlation was only observed between the aimed agricultural trait such as stem strength and CAD enzymatic activity, whereas no correlation was found between CAD gene expression and stem strength [64]. There is a great need of functional characterization of strawberry CAD genes via overexpression and CRISPR/Cas9-mediated genome editing methods. The corresponding responsive gene in F. vesca and their dominant allele in F.×ananassa could be edited by CRISPR-Cas9 technology and overexpressed to reveal their functions, and to convert a susceptible variety into a resistant one in the future.
Future studies on the molecular mechanisms underlying these responsive CADs in strawberry immunity have great potentials for improving strawberry resistance breeding. It is worthwhile to explore the upstream and downstream events in CAD-dependent resistance. AtCAD1 not only functions in lignification, but regulates the programmed cell death via inhibiting the expression of PR genes and NPR1 of the SA-dependent defense pathway [65]. Similarly, silencing cotton class I GhCAD35 and Class II GhCAD45 led to the decrease of SA level and the transcript level of PR1, ICS1 in cotton at 24 hpi with V. dahliae inoculation, indicating that SA-dependent defense pathway at the early infection stage was altered [25]. In Arabidopsis over-expression of AtCAD7 led to a significant down-regulation of the transcript level of WRKY33, a key transcriptional factor crucial for resistance to necrotrophic pathogens [27]. Undoubtedly, changes in SA-dependent resistance pathway and relevant transcriptional factors in the transgenic materials with certain CAD-knockout or -overexpressed are most promising. Such work is underway in our group.
Author contributions
K. D., Q-H. G. and C-N. G. conceived this work. Z-Y. L. carried out experiments and data analysis. J-J. Z. contributed to collinearity analysis; B. L. performed inoculation and phenotyping; X-Y. W, S-Y C and X. L. undertaken RNA extraction. Z-Y. L. prepared tables and figures. Z-Y. L. and K. D. wrote the paper. All authors contributed to revising and approved the manuscript.
Footnotes
Acknowledgments
This work was funded by Shanghai Agriculture Applied Technology Development Program, China (Grants No. G2016060104 to KD and No. X2019-02-08-00-08-F01108 to QHG) and by funds from Shanghai Academy of Agricultural Sciences (Grants No. JCYJ232201). We are grateful to Ms Yun Bai for valuable guidance in bioinformatic analysis. Thanks are due to anonymous reviewers for valuable comments which improved this manuscript.
Conflict of interest
The authors have no conflict of interest to report.
Supplementary data comprising 2 tables and 4 figures are available in the electronic version of this article.![]() .
.
Supplementary Table S1. The primers used for RT-qPCR analysis of strawberry CADs responses to C. fructicola.
Supplementary Table S2. The cis-elements information for the 2500bp promoter regions of CAD family genes in F. vesca and F.×ananassa.
Supplementary Figure S1. The intron-exon structure of the CAD genes from Arabidopsis, F. vesca and F.×ananassa. The CDS and DNA sequences of each CAD member were analyzed with the gene structure display server (GSDS) (![]() ), accessed on 10 June 2022.
), accessed on 10 June 2022.
Supplementary Figure S2. The comparative protein domain modules of CAD members from F.×ananassa and F. vesca. The locations of conserved ADH_N domain and ADH_ZINC_N domain (identified at PFAM on 7 November 2022) were indicated. Visualization was improved with IBS software.
Supplementary Figure S3. The motif compositions of the CAD proteins from Arabidopsis, F. vesca and F.×ananassa. Distinct motifs varying in colors were identified at MEME (![]() , accessed on 21 August 2021).
, accessed on 21 August 2021).
Supplementary Figure S4. The alignment of CAD protein sequences from Arabidopsis and strawberry. Present were 9, 14 and 32 CAD members from Arabidopsis, F. vesca, and F.×ananassa, respectively. The conserved Zn1 catalytic center is marked with red triangles. The Zn, Zn2, and NADPH binding sites are highlighted with red boxes.