
Abstract
BACKGROUND:
The fig (Ficus carica L.) tree known for its tasty and nutritious fruits, is typically propagated by cutting. While previous studies have focused on the effects of different treatments and environmental conditions on fig cutting propagation, little attention has been paid to the specific role and association of biochemical properties in leaves, stem bark and fruit on the rooting process.
OBJECTIVE:
This research explores the complex relationship between 40 biochemical traits and the rooting ability of fig cuttings. To achieve this objective, various machine learning techniques were employed, such as a random forest model, feature importance analysis, linear regression, and principal component analysis (PCA).
RESULTS:
The random forest model showed significant predictive ability with a classification accuracy of 100%, supported by a high kappa statistic. Feature importance analysis identified a* (a colorimetric parameter in fruit), fruit trans-ferulic acid and leaf total flavonoids as the most influential traits in determining the rooting ability of cuttings. The robustness of these findings is supported by the high R-squared value (0.9002) and low error metrics (MAE 0.7554 and MSE 0.6980) of the linear regression model built on these important traits. In parallel, PCA indicated that a*, leaf total flavonoids and fruit trans-ferulic acid were the dominant traits in samples with lower rooting percentage.
CONCLUSIONS:
These identified biomarkers can be effectively used by fig breeders and growers to select and introduce fig cultivars with improved rooting ability.
Highlights
The random forest model achieved a classification accuracy of 100% and thus showed an excellent prediction of the rooting ability of fig cuttings. Feature importance analysis revealed that the fruit colorimetric parameter a*, fruit trans-ferulic acid content and leaf total flavonoid content are associated with the rooting ability of fig cuttings. The linear regression model, with an R-squared value of 0.9002 and low error metrics (MAE 0.7554, MSE 0.6980) underlines the strong predictive relationship between selected biochemical traits and rooting ability. The principal component analysis confirmed that total leaf flavonoids, fruit a* and trans-ferulic acid were the dominant traits in the samples with lower rooting percentages. The study identifies biomarkers useful for selecting fig varieties with better rooting ability for breeding. The study shows the potential of data-driven approaches to help growers improve their fig-growing practices.
Introduction
The fig (Ficus carica L.), a fruit tree of great economic and ecological importance, shows remarkable adaptability to diverse environmental conditions [1]. Consequently, it has become a valuable crop in many regions. For the expansion of fig orchards, the propagation of fig trees through hardwood stem cuttings is a widely used practice. The production of fig nursery stock is mainly based on cuttings obtained from the mother tree and placed in a rooting medium where they develop roots and shoots [2–4]. The traditional method of fig cultivation involved planting cuttings taken from one- or two year old shoots directly into the soil to establish new trees [2].
The rooting ability of cuttings is influenced by a combination of genetic, environmental and physiological factors [5, 6]. Several studies have focused on the effects of different treatments and environmental conditions on fig cutting propagation [7, 8]. Plant growth regulators, particularly auxins, play a significant role in stimulating and initiating root formation in fig cuttings. Auxins, such as IBA and NAA, have been found to improve the rooting ability of fig cuttings by inducing root formation [9, 10]. Furthermore, the concentration of endogenous and exogenous phytohormones, as well as the physiological and biochemical characteristics of the cuttings, can impact their ability to successfully root [10]. In addition, the anatomical structure of the stem cuttings and the duration of shading of the mother plants were found to influence the rooting performance of different fig cultivars [11].
In our previous study [12], we examined the correlation between rooting percentage and biochemical traits specifically in stem bark using univariate statistical methods. However, the specific role of biochemical traits in leaves, stem bark and fruit in the rooting process of cuttings has received relatively little attention and previous research has not identified specific biochemical markers associated with rooting percentage in fig cuttings. In the current study, our objective is to extend our analysis to include biochemical traits not only in stem bark but also in leaves and fruit. To achieve this, we employ machine learning techniques (random forest) and multivariate statistical methods. By analyzing multiple plant tissues, our study aims to provide a more comprehensive understanding of the relationship between rooting ability and biochemical composition in different plant tissues. This approach will yield valuable insights into the factors influencing rooting success in fig cultivars, filling a significant gap in the current knowledge.
In recent years, machine learning techniques have gained popularity in agricultural research due to their ability to analyze complex data sets and identify patterns that may not be immediately apparent using traditional statistical methods [13]. The use of these techniques can provide valuable insights into the relationship between biochemical traits and the rooting ability of fig cuttings. Random forest models have emerged as valuable tools in horticultural research, as evidenced by several studies. For instance, Fukuda, Spreer [14] employed random forest models to estimate mango (Mangifera indica) fruit yields under different irrigation regimes, highlighting the potential of machine learning models to optimize irrigation practices in horticulture. Similarly, Johansen et al. (2020) [15] utilized random forest models to predict biomass and yield in a tomato (Lycopersicon esculentum) phenotyping experiment using multispectral unmanned aerial vehicle (UAV) imagery data. The study showed that the random forest approach outperformed other methods by being less sensitivite to data skewness and preventing model overfitting. Another study by Ropelewska and Noutfia [16] employed random forest models to assess grape (Vitis spp.) berries under different storage conditions. The study highlighted the exceptional 96% classification accuracy of the random forest model in differentiating grapes stored under different conditions. Furthermore, Mokhtar, El-Ssawy [17] utilized random forest models, along with other machine learning models, to predict the yield of hydroponically grown lettuce (Lactuca sativa). Their study demonstrated the potential of machine learning models in accurately forecasting crop yield in hydroponic systems. Together, these studies underscore the efficacy of random forest models in various horticultural applications, including yield estimation, disease detection, and crop behavior assessment.
This study aims to investigate the influence of various biochemical traits in fig leaves, stem bark, and fruits on the success of fig propagation by cuttings and to identify biochemical markers associated with root formation. We employed machine learning methods and principal component analysis (PCA) to identify the key biochemical factors that significantly influence the rooting potential of fig cuttings. The results of this research could have practical applications in improving easy-to-root fig cultivars, propagation techniques and ultimately contributing to more efficient and sustainable fig cultivation.
Materials and methods
In the first part, we analyzed the rooting characteristics and biochemical traits of four Iranian figs (Ficus carica cvs. ‘Siyah’, ‘Kashki’, ‘Shahanjir’ and ‘Sabz’). The evaluation included measuring the rooting percentage, root fresh and dry weights, and various biochemical markers in each fig cultivar. We thenused an advanced machine learning technique, specifically the random forest model to predict and identify the most significant traits associated with rooting.
Measurement of traits and biochemical analysis
Plant materials
In mid-August, 12 ripe fruits and 40 mature leaves were collected from the four sides of the tree, from the middle part of the growing branches of the current season. In mid-December 2022, 12 hardwood stem cuttings with a length of 22 cm were prepared from the fully mature annual shoots of each tree (48 cuttings in each cultivar) from the Estahban Fig Research Station, at the GPS coordinates latitude: 29.115078 degrees, longitude: 54.073986 degrees, at an altitude of 1736 meters above sea level. The soil texture at the site is loamy-clay with a pH value of 7.8. The average annual rainfall is 256 mm. The highest temperature in July reaches up to 45°C, while the lowest temperature in January is between –6°C and –7°C. Relative humidity averages between 11% and 68%. The region experiences 600 chill units annually. The bark samples were prepared from the proximal end of the cuttings. Each biochemical analysis was performed with four replicates and the final values presented in Table 1 are the averages of these replicates.
Analysis of rooting characteristics and biochemical traits across four fig cultivars. This table presents the mean values and standard deviations for traits such as rooting percentage, root weights, and biochemical markers in ‘Siyah’, ‘Kashki’, ‘Shahanjir’ and ‘Sabz’
Analysis of rooting characteristics and biochemical traits across four fig cultivars. This table presents the mean values and standard deviations for traits such as rooting percentage, root weights, and biochemical markers in ‘Siyah’, ‘Kashki’, ‘Shahanjir’ and ‘Sabz’
The color of the fruit peel was determined using a colorimeter (Chroma Meter CR-400, Minolta, Japan) as L*, a*, b*, C (chroma), and H° (hue angle) and the TSS (total soluble solids) (°Brix) in the fruit extract was determined using a refractometer (Pocket Mode S model, made in Japan).
Total phenolics and flavonoids
Total phenolic content was determined using the Folin-Ciocalteu’s reagent and a microplate reader (Bio Tek ELx808) at 750 nm, and the results were expressed as gallic acid equivalents. The total flavonoid content was determined using 5% sodium nitrite, 10% aluminium chloride, and 25% sodium hydroxide. Finally, the absorbance of the samples was measured at a wavelength of 510 nm using a microplate reader (Bio Tek ELx808) [18, 19].
Phenolic compounds analysis
The extraction and determination of phenolic compounds in fruits, leaves and cutting bark were carried out according to the method described by Mišan et al. (2011) [20] with some modifications using the HPLC system (Agilent 1200 series).
Antioxidant capacity
The radical scavenging activity of fruits and leaves extracts was determined using the DPPH method with some modifications. The decrease in absorbance of DPPH at 520 nm was measured with a spectrophotometer (UV-1800, Shimadzo, Japan), until the absorbance stabilized (30 min) [21, 22].
Fruit mineral nutrient analysis
Fig fruits were washed with distilled water and dried at 70°C for 48 h. After drying, they were ground into fine particles. The ground fruit samples were then ashed at 500°C, and further digested with 2N hydrochloric acid. The phosphorus (P) concentration in the extract was determined using the yellow color method [23]. Potassium (K), calcium (Ca), and sodium (Na) concentrations were measured using a flame photometer (Corning 405, ELE, UK), and iron (Fe), manganese (Mn), zinc (Zn), and copper (Cu) concentrations were measured using an atomic absorption spectrophotometer (AAS; PG 990, PG Instruments Ltd. UK).
Rooting of stem cuttings and polyphenol oxidase (PPO) activity
The cuttings were washed with water, and then their ends were placed in a solution of indole butyric acid (IBA) at a concentration of 750 mg/l for 10 seconds. The cuttings were then transferred to the culture medium containing perlite [24]. After 3 months, once the rooting process was completed, the number of rooted cuttings was recorded, and both the fresh and dry weights of the roots in each cutting were measured. The percentage of rooting was then calculated based on these measurements. To determine the PPO enzyme activity, a modified version of the method described by Fujita, Saari [25] was used.
Machine learning and multivariate statistical methods
To analyze the relationship between biochemical traits and rooting ability, we utilized machine learning and multivariate statistical methods.
Random Forest Model: This model was trained using the dataset to predict the rooting ability of fig cuttings. The performance of the model was assessed using classification accuracy, kappa statistic, mean absolute error (MAE) and mean squared error (MSE).
Feature Importance Analysis: This analysis identified the most influential biochemical traits contributing to the prediction of rooting ability. Importance scores were derived from the internal algorithm of the random forest model.
Linear regression analysis: A linear regression model was developed using the key biochemical traits (X6: a* color parameter, X26: trans-ferulic acid in fruit, and X15: leaf total flavonoids). The key traits X6, X26 and X15 were selected based on their importance scores obtained from feature importance analysis, indicating that they had the greatest influence on the predictions of the random forest model for rooting ability. The performance of the regression model was assessed using R-squared, adjusted R-squared, mean absolute error (MAE) and mean squared error (MSE).
Principal component analysis (PCA): PCA was performed to gain a better understanding of how key biochemical traits and rooting ability are related in fig cultivars.
These methods are described in detail below.
Data preprocessing
This study utilized a dataset consisting of 16 records and 41 features (traits). Initially, the data underwent standardization to convert the values into standardized scores using the scale function. Subsequently, the dataset was divided into training (70%) and testing (30 .%) sets.
Model construction
Random forest, an ensemble learning algorithm, was employed to build the prediction model. The model parameters were set as follows: the number of trees (ntree) was set to 500, the number of features considered for splitting at each node (mtry) was set to 4, and the maximum depth of the trees (max_depth) was set to 5. These parameter settings were chosen to optimize the performance of the model in predicting the desired outcomes.
Model evaluation
The constructed model was evaluated on the testing dataset. Several metrics were computed to assess the performance of the model, including accuracy, mean squared error, and the confusion matrix.
Feature importance analysis
To gain insights into the predictive power of different traits, a feature importance analysis was conducted using the random forest model.
Importance scores for each trait were extracted from the trained random forest model. These scores represent the contribution of each trait to the overall predictive accuracy of the model. The feature importance scores were visualized using a horizontal bar chart, where traits were reordered based on their importance scores. The color gradient ranging from blue (low importance) to red (high importance) was applied for clear visualization.
Linear regression model
For a more in-depth analysis and precise prediction of the target variable, we employed linear regression. In this step, the regression model was applied to the chosen traits, which were identified as influential through the random forest algorithm. The model’s predictions were then assessed and validated against the test dataset, providing valuable insights into the relationships between the selected traits and the target variable. The four statistical evaluation criteria were used to assess the performance of the model, including the coefficient of determination (R2), adjusted R-squared (Adj R2), mean absolute error (MAE) and mean squared error (MSE) for the test dataset. These statistical indices were calculated as follows:
Coefficient of determination (R2):
Where y
i
are the actual values, ŷ
i
are the values predicted by the model, and
Adjusted R-squared (Adj R2):
Mean squared error (MSE):
Mean absolute error (MAE):
Heatmap visualization
A heatmap was generated using the ClusVis tool to visualize four traits (a*, trans-ferulic acid in fruit, leaf total flavonoids, and rooting percentage) across four different fig cultivars, each with four replicates. The web-based open tool Clustvis (https://biit.cs.ut.ee/clustvis/) utilizes several R packages internally, including pheatmap, ggplot2, brilliant, gridSVG, gProfileR, FactoMineR, RColorBrewer, pcaMethods, shinyjs, and shinyBS [26].
Principal component analysis (PCA)
The PCA was conducted using base R functions to investigate the relationship between key traits associated with rooting ability in fig cutting and the rooting percentage in fig samples.
The schematic diagram of the integrative strategy is illustrated in Fig. 1.

Schematic overview of the integrative strategy for revealing key traits associated with rooting ability in fig cutting.
The results of the analysis of rooting and biochemical characteristics of the four fig cultivars are presented in Table 1. The table contains the means and standard deviations of traits such as rooting percentage, root fresh and dry weight, and biochemical markers in the evaluated cultivars. The analysis highlights the diversity and unique profiles of each cultivar.
The random forest model was trained on the provided dataset, and according to the results of the prediction, all 4 instances in the test dataset were correctly classified, resulting in a classification accuracy of 100%. The Kappa statistic, which measures the agreement between the model predictions and the actual values, was calculated as 1. This high value indicates excellent model performance. The MAE for the predictions was found to be 0.3654, reflecting the average absolute difference between predicted and actual values. The MSE, a measure of the average squared differences between the predicted and actual values, was calculated as 0.1916. The model achieved a 100% accuracy rate, correctly classifying all instances in the test dataset. The error rate, representing the proportion of incorrectly classified instances, was found to be 0%.
Feature importance analysis
Trait importance analysis provides insights into which traits (biochemical traits) contributed the most to the random forest model’s predictions for the rooting ability of fig cuttings. The importance values, calculated based on the model’s internal algorithm, are shown in Fig. 2. These importance scores indicate the relative contribution of each trait in the model’s decision-making process. Traits with higher importance values have a greater impact on the model’s predictions. In this study, a*, trans-ferulic acid (mg/100 g dw) in fruit, and leaf total flavonoids were among the most influential traits in determining the rooting ability of fig cuttings.
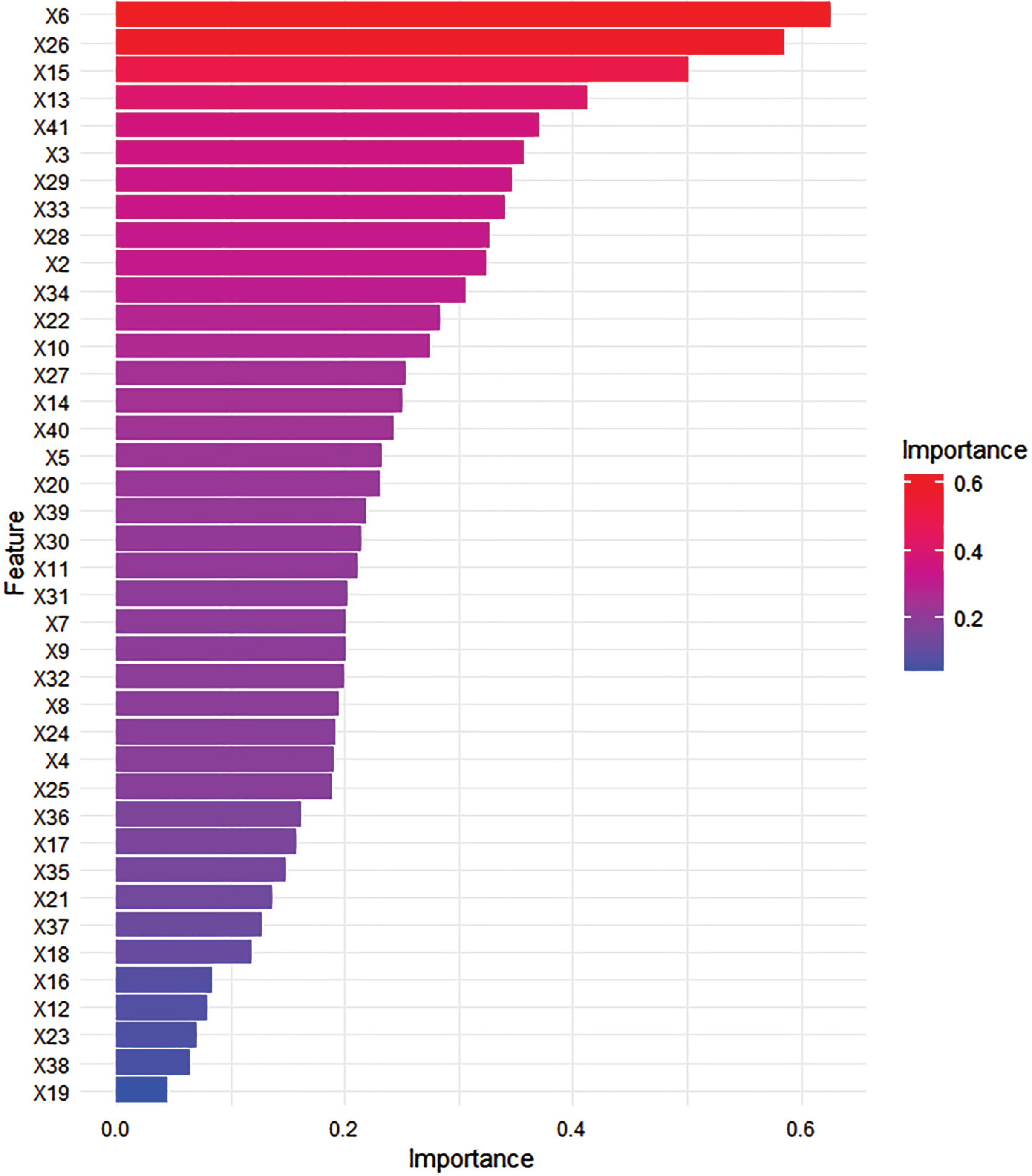
Relative importance of biochemical traits in predicting rooting ability of fig cuttings. This figure illustrates the calculated importance scores for each trait, highlighting a*, trans-ferulic acid content in fruit, and leaf total flavonoid levels as key determinants in the model’s ability to predict rooting success in fig cuttings. For a comprehensive understanding of the variables x1 to x41 and their respective values, Table 1 is to be referred to.
The linear regression model was developed to predict the rooting ability of fig cuttings based on a subset of key biochemical traits, including a*, trans-ferulic acid (mg/100 g dw) in fruit, and total flavonoids in leaves. The model’s performance was evaluated using various metrics. Table 2 presents the MAE, MSE, R2, and adj R2 values for the linear regression model using the key traits X6, X26, and X15. The MAE for the linear regression model was calculated as 0.7554. This metric represents the average absolute difference between the actual values and predictions made by the model, providing a measure of the model’s accuracy. Lower MAE values indicate better performance. The MSE was 0.6980. The MSE measures the average squared differences between actual and predicted values. A lower MSE value indicates that the model predictions are closer to actual values. The R-squared (R2) value, a measure of the goodness of fit, was found to be 0.9002. R-squared quantifies the proportion of variance in the dependent variable that is explained by the independent variables. A higher R-squared value signifies a better fit of the model to the data. The adjusted R-squared (Adj R2) value, a modification of the R-squared value, was calculated to be 0.8004. The Adj R2 accounts for the number of independent variables in the model, providing a more reliable measure of goodness of fit, especially when there are multiple predictors.
Linear regression model performance metrics
Linear regression model performance metrics
The results demonstrate that the linear regression model, utilizing the selected important traits, performs well in predicting the rooting ability of fig cuttings. The linear regression model is represented by the following equation:
The heatmap generated using the ClusVise tool provides a detailed visualization of the biochemical traits across different fig cultivars. These differences are represented using a color scale, where higher values are indicated in red and lower values in blue (Fig. 3).
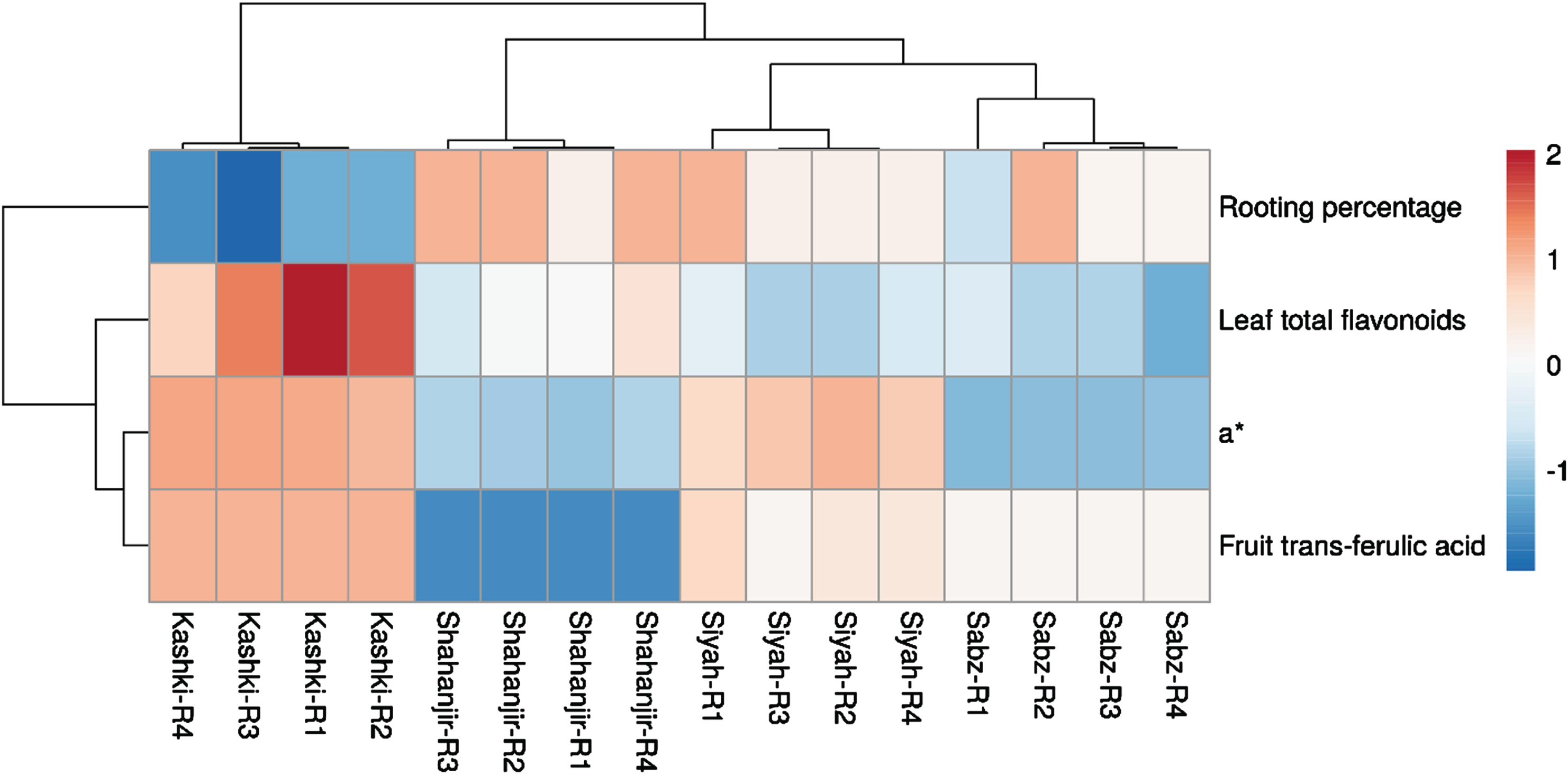
Heatmap of the four traits in four cultivars (Ficus carica cvs. ‘Siyah’, ‘Kashki’, ‘Shahanjir’, and ‘Sabz’) samples. The traits, which were hierarchically clustered, are shown as rows, while the samples are shown as columns. The data of the traits have been normalized and are presented using a colour scale. Higher values are shown in red, while lower values are shown in blue. The codes R1, R2, R3, and R4 represent the four replicates of each fig cultivar.
Furthermore, according to PCA analysis, PC1 and PC2 together accounted for 86.25% of the variance in the dataset, a finding that is particularly noteworthy (Fig. 4). The PCA Biplot illustrates the relationships between the samples as well as the measured traits in figs. The cosine of the angles between vectors indicates the degree of correlation between traits. The angles between rooting percentage vector with a*, trans-ferulic acid content in fruit, and leaf total flavonoid levels were obtuse, indicating a negative correlation (Fig. 5).

The bars show the percentage of variance of the dataset explained by each principal component.

PCA Biplot of four traits including a*, leaf total flavonoids, fruit trans-ferulic acid, rooting percentage in 16 samples. The codes R1, R2, R3, and R4 represent the four replicates of each fig cultivar.
A dataset with frothy-one traits was used in this study. These data included a dependent variable (rooting percentage) named “Y”. The dataset was preprocessed and standardized prior to analysis to ensure consistency and accuracy. The results demonstrate the effectiveness of the random forest model in predicting the rooting ability of fig cuttings based on the biochemical traits of leaves, stem bark, and fruits. The model achieved a perfect classification accuracy of 100%, indicating its ability to accurately distinguish between successful and unsuccessful rooting outcomes. Additionally, the high Kappa statistic of 1 suggests strong agreement between the model’s predictions and actual values.
The model’s performance metrics, including MAE and MSE, indicate that the predicted values are close to the actual values. This suggests that the model’s predictions are not only accurate but also precise. This precision can be particularly valuable for fine-tuning propagation strategies.
The absence of misclassified instances (error rate of 0%) is noteworthy, as it highlights the robustness of the random forest model in fig cutting rooting prediction.
These findings have several implications for fig propagation and orchard management. The ability to predict fig cutting rooting potential with high accuracy can significantly benefit agricultural practices. By leveraging machine learning techniques, such as random forest, growers, and horticulturists can optimize their fig propagation processes, leading to higher success rates in expanding fig orchards.
In summary, our study demonstrates that random forest, with optimized hyperparameters, serves as a powerful tool for predicting fig cutting rooting ability based on biochemical traits. The proposed approach has practical applications in the fields of agriculture and horticulture, offering an efficient and data-driven means of improving fig propagation processes. Given the crucial role of fig cultivation in agriculture and ecosystems, advances in propagation techniques can yield broad economic and ecological implications.
Feature importance analysis
The feature importance analysis provides valuable insights into the biochemical traits that play a pivotal role in determining rooting ability of fig cutting. By identifying the most influential traits, we can gain a deeper understanding of the factors that significantly impact the success of fig propagation. In our random forest model, each trait was assigned an importance score, indicating its contribution to the model’s predictions. Figure 2 clearly shows the relative importance of each trait, aiding in the interpretation of the model’s behavior and contributing to the understanding of the underlying data patterns.
Among the traits examined, a*, trans-ferulic acid (mg/100 g dw) in fruit, and leaf total flavonoids emerged as the most influential factors in predicting rooting ability of fig cutting. The a* exhibited the highest importance score, emphasizing its critical role in root formation. In colorimetry, the a* value is a key indicator of redness in objects. A higher a* value suggests a red color, while a lower or negative value indicates greenness. In the context of fig fruits, this parameter reflects the degree of red pigmentation in the skin. Our study revealed an intriguing negative correlation between the a* value in fig fruit skin and the rooting success rate of cuttings. Essentially, this implies that figs with redder skins tend to have cuttings that are less successful at root formation. Furthermore, higher levels of certain hormones that promote red pigmentation might inversely affect the plant’s capacity to allocate resources for root development in its cuttings. In other words, the resources that a plant normally uses for root development might be redirected toward the production of red pigments. Significantly, the process of root formation in cuttings, also known as adventitious rooting, is a complex process influenced by various factors, such as phytohormones and resource allocation, as highlighted by Druege et al. (2019) [27]. Our research offers a unique perspective on the relationship between a visible characteristic of the fruit, color, and a critical aspect of plant propagation, root formation. These findings underscore the complexity of plant physiology and the interconnectedness of various plant traits. Further investigation into the biochemical properties and physiological mechanisms associated with a* could provide valuable insights for fig growers seeking to optimize their propagation techniques.
The second most important trait, trans-ferulic acid (mg/100 g dw) in fruit, is another key determinant of fig cutting rooting success. We observed an inverse correlation between the levels of trans-ferulic acid in fruits and the ability of plants to develop roots. We hypothesize that the increase in trans-ferulic acid levels in fruit during the fruiting phase in summer may lead to a lower concentration of this compound within the plant, potentially influencing root development during winter. Our hypothesis underscores the intricate relationship between seasonal variations in a plant’s biochemical profile and its growth, indicating that changes in metabolite concentrations over time could greatly affect plant developmental processes. Trans-ferulic acid, a hydroxycinnamic acid commonly found in plant cell walls, is primarily esterified to polysaccharides [28]. It plays a crucial role in providing structural support to the cell wall and participating in the synthesis of important compounds. One notable compound derived from trans-ferulic acid is vanillin [29], which has been implicated in root development. Research indicates that vanillin content is higher in roots, particularly in the root cortex, compared to the above-ground parts of plants [30]. The effects of trans-ferulic acid on root formation vary depending on the concentration and the specific plant species. For example, a study on Malus “Jork 9” demonstrated that trans-ferulic acid increased the number of adventitious roots from 0.9 to 5.8, indicating a positive influence on root formation [31]. However, the same study also revealed that the impact of phenolics, including trans-ferulic acid, is influenced by the timing of application, suggesting that other factors can modulate the effect of trans-ferulic acid on root formation [31]. In a study on mung bean (Vigna radiata), it was observed that ferulic acid (FA) can interfere with vital biochemical processes essential for root growth and development, suggesting a potential inhibitory effect of trans-ferulic acid on root formation. The study concluded that trans-ferulic acid inhibits root growth, development, and in vitro rooting processes in mung bean by disrupting these biochemical processes. Furthermore, the study highlighted the concentration-dependent nature of trans-ferulic acid’s effects, with different concentrations yielding different impacts on root formation [32]. This suggests that trans-ferulic acid’s influence on root development is not solely determined by its presence but also by the specific concentration at which it is applied. Overall, trans-ferulic acid, as a key biochemical attribute, plays a multifaceted role in root development, with the potential to affect root formation positively or negatively depending on various factors such as concentration, timing, and plant species.
However, Table 1 shows that trans-ferulic acid in fruit was detected at very low levels or not at all in some samples. Despite these low levels, our statistical model, particularly the random forest, highlights its importance in predicting rooting ability. This can be explained by the fact that machine learning models such as the random forest are able to detect subtle patterns and relationships in the data that may not be immediately apparent through traditional analysis [33]. The model showed that the presence or absence of trans-ferulic acid, even at trace levels, is strongly correlated with rooting percentages.
Moreover, the literature shows that certain compounds, even at low levels, can significantly affect physiological processes [34, 35]. Thus, although trans-ferulic acid is not abundant, it could still play a crucial role in the biochemical pathways related to root development. Additionally, the interaction between trans-ferulic acid and other biochemical traits could enhance its effect on rooting [36, 37]. Thus, the results of the model suggest that these small amounts of trans-ferulic acid are indeed biologically important for root development.
Leaf total flavonoids play a substantial role in the model’s predictions. As previously mentioned, there is a negative correlation between flavonoid content in leaves and root formation. High flavonoid levels might indicate a significant investment in leaf development or protective mechanisms against summer stress. This allocation could influence the availability of resources necessary for root development from cuttings in winter. Flavonoids interact with several plant hormones, including those involved in root development. The altered hormonal environment resulting from high summer flavonoid levels could have lasting effects, potentially impacting the plant’s capacity to form roots from cuttings in winter. The involvement of different flavonoids on the rooting process of cuttings has been well revealed. For example, the concentration of compounds such as rutin and quercetin in olive cuttings of Arbequina cultivar (easy-to-root) is higher during both the induction and root initiation phases compared to Kalamata (difficult-to-root) cultivar. These compounds play a role in protecting Indole-3-acetic acid (IAA) against IAA oxidase, which ultimately contributes to increased rooting [38].
The formation of adventitious roots includes many physiological and molecular changes that are affected by many internal and external factors, including plant growth regulators, hormones and secondary metabolites such as phenolic compounds [38-41]. While previous research has shown that higher phenolic content (total phenol) of the cuttings accelerates rooting [42, 43], it has been found that specific phenolic compounds have different effects on the rooting process of cuttings, which may also depend on the cultivar. Some phenolic compounds have no effect on this process [44], while others, such as sinapic acid and vanillic acid in sweet cherry cuttings [44], tannic acid and gallic acid in chestnut cuttings [45] and cinnamic acid in chamaelaucium hardwood stem cuttings [46], have a negative effect on rooting due to the stimulation of IAA oxidation. Phenolic compounds can act as antioxidants and protect IAA from decarboxylation and tissue from oxidative stress [31, 42]. Additionally, certain compounds such as chlorogenic acid in olive cuttings stimulate the translocation and accumulation of starch and sugars at the base of the cuttings, altering sink strength and thereby accelerating rooting, particularly in the initial stages of the rooting process [47]. Indeed, some phenolic compounds serve as precursors for lignin synthesis, thereby influencing stem anatomy and the formation of adventitious roots [48]. It has been shown that the addition of various phenolic compounds to the Jork9 stem on the MS medium accelerates rooting. Among these compounds, the effects of ferulic acid and phloroglucinol were the highest [31].
It is worth noting that while these traits emerged as the most important traits in our model, the interplay of multiple biochemical traits should not be overlooked. A holistic approach to fig propagation, considering the complex interactions between various traits, is essential for comprehensive orchard management.
Based on the advanced machine learning employed in our study, the traits of a* color parameter, trans-ferulic acid content in fruit, and leaf total flavonoid content have emerged as potential biomarkers for predicting and enhancing the rooting ability of fig cuttings in horticultural practices. These findings provide valuable insights for horticulturists to focus on these specific traits when selecting and preparing fig cuttings for propagation. By considering these biomarkers, they can optimize their practices and improve the success rate of fig cutting rooting, leading to more efficient and effective horticultural outcomes.
Linear regression model
Our results indicate that the linear regression model, built on a subset of critical traits, offers a promising framework for predicting the success of fig propagation. We selected a subset of traits, namely a*, trans-ferulic acid (mg/100 g dw) in fruit, and leaf total flavonoids, based on their importance scores. These traits were selected due to their significant influence on the model’s predictions. The linear regression model exhibited a strong performance, as evidenced by its high R2 of 0.9002. This indicates that a substantial proportion of the variance in rooting ability of fig cutting can be explained by the selected traits. The accuracy of the model, reflected in the low MAE of 0.7554 and MSE of 0.6980, suggests that the model’s predictions closely align with the actual values. These metrics underscore the model’s precision in estimating rooting success. The coefficients of the linear regression model provide insights into the relationships between the selected traits and the rooting ability of fig cutting. For instance, negative coefficients, such as those of a*, trans-ferulic acid in fruit, and leaf total flavonoids, suggest that higher values of these traits are associated with reduced rooting ability. These interpretations can guide growers and researchers in understanding the biochemical factors affecting fig propagation. By focusing on the critical biochemical traits identified in this analysis, growers can make informed decisions about propagation techniques, and orchard management. While the linear regression model shows promise, continued research is necessary to validate its performance in real-world fig orchards and under diverse environmental conditions. Additionally, exploring the interactions between multiple traits and considering additional factors can enhance the accuracy and applicability of the model.
In conclusion, linear regression analysis of important biochemical traits provides a reliable framework for predicting the rooting ability of fig cutting.
The heatmap showes detailed differences in biochemical traits among the different fig cultivars, highlighting significant variations in rooting percentage, leaf total flavonoids, and fruit trans-ferulic acid levels. The heatmap’s hierarchical clustering of traits and cultivar samples offers insights into the relationships between these biochemical traits and root outcomes (Fig. 3), supporting the findings regarding the potential of data-driven approaches in optimizing fig propagation. The heatmap also shows a negative correlation between the biochemical traits a*, trans-ferulic acid in fruit, and leaf total flavonoids with rooting percentage, suggesting that higher values of these traits may inhibit root formation in fig cuttings.
Principal component analysis (PCA)
PCA of the biochemical traits of a*, leaf total flavonoids, fruit trans-ferulic acid, and their relationship with rooting percentage in fig samples yielded insightful results. The analysis revealed that the first two principal components PC1, and PC2 cumulatively accounted for 86.25% of the variance in the dataset, underscoring the strong interconnection between these biochemical traits and rooting efficiency. PC1 explains 68.19% of the variance, indicating that a significant portion of the total variability in the dataset is captured by this component alone. The rooting percentage had a high positive loading (0.53) on PC1, suggesting that it was the most influential variable in this component in the same direction compared with other variables (a*, leaf total flavonoids, fruit trans-ferulic acid) (Table 3). This indicates that a substantial portion of the variability in rooting percentage can be attributed to variations in leaf total flavonoids, fruit trans-ferulic acid, and a*, as well as their individual and combined effects on rooting percentage. Furthermore, leaf total flavonoids have a very high negative loading (–0.71) in PC2, making it the most dominant variable in this component (Table 3). As revealed in the PCA Biplot, a*, leaf total flavonoids, and fruit trans-ferulic acid were dominant traits of the samples (‘Kashki’) with lower rooting percentages. These findings support the hypothesis and suggest further investigation into how these specific biochemical traits influence root development in figs.
PC1, PC2, PC3, and PC4 loadings from the PCA of four traits including a*, leaf total flavonoids, fruit trans-ferulic acid, rooting percentage
PC1, PC2, PC3, and PC4 loadings from the PCA of four traits including a*, leaf total flavonoids, fruit trans-ferulic acid, rooting percentage
In this study, we conducted an extensive analysis of fig cutting rooting ability, focusing on the influence of biochemical traits on the prediction of successful root formation. We used a random forest model, a feature importance analysis and linear regression to gain insights into the factors that contribute to successful fig propagation. The random forest model demonstrated excellent predictive power, achieving 100% classification accuracy. This signifies that the model can accurately distinguish between successful and unsuccessful fig cutting rooting based on biochemical traits. The high Kappa statistic further reinforces the model’s effectiveness. As a result, this model has great potential for aiding agricultural and horticultural practices by enhancing fig propagation processes and orchard management. Feature importance analysis unveiled key biochemical traits that strongly influence fig cutting rooting ability. The a*, trans-ferulic acid (mg/g) in fruit, and total leaf flavonoid emerged as the most influential traits. The analysis forms the basis for targeted interventions in fig cultivation. Linear regression analysis, focusing on important traits, provided a robust framework for predicting fig cutting rooting success. The model’s high R-squared value (0.9002) and low error metrics (MAE of 0.7554 and MSE of 0.6980) signify its accuracy and precision in estimating rooting ability. The coefficients of the model offer interpretability, allowing growers to understand the biochemical factors impacting fig propagation. In addition, the results of PCA confirm the results expressing that the increase in a*, leaf total flavonoids, and fruit trans-ferulic acid results in a lower rooting percentage. However, it is crucial to further research and validate these models in real-world fig orchards to ensure their practical applicability, especially in diverse environmental conditions. Our study identified important biochemical traits such as a* (a colorimetric parameter in the fruit), fruit trans-ferulic acid, and leaf total flavonoids, that significantly affect the rooting ability of fig cuttings. These findings have important implications for fig breeders and physiologists seeking to improve fig cultivation.
First, identifying traits related to rooting, particularly appearance traits such as fruit color indicators, will allow breeders to select fig cultivars with better rooting ability through common selection methods. This can lead to higher success rates in establishing new plants, supporting sustainable production practices. Second, understanding the role of these biochemical traits can provide insights into the biochemical pathways that are crucial for root development. This knowledge can be used to optimize these pathways either through genetic engineering or by altering growth conditions.
In summary, the newly identified biochemical traits not only aid in improving the propagation efficiency of fig plants but also contribute to sustainable agricultural practices and improved nutritional quality. Future research should further explore these traits and their interactions to maximize their potential benefits.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Availability of data and materials
All data has been provided with the manuscript. If any additional information is required, then the corresponding author can be contacted.
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval and consent to participate
This work does not involve living animals and no consent is needed.
Author contributions
Conceptualization: Zahra Zinati, Abbas Mirsoleimani
Formal Analysis: Abbas Mirsoleimani, Zahra Zinati
Investigation: Abbas Mirsoleimani, Zahra Zinati, Shima Abbasi
Methodology: Abbas Mirsoleimani, Zahra Zinati, Shima Abbasi
Project administration: Abbas Mirsoleimani, Zahra Zinati
Supervision: Abbas Mirsoleimani
Visualization: Zahra Zinati
Writing – original draft: Zahra Zinati, Abbas Mirsoleimani
Writing – review & editing: Zahra Zinati, Abbas Mirsoleimani
All authors declare that they had access to the data used in this study.