
Abstract
BACKGROUND:
In addition to calcium and phosphate homeostasis in peripheral tissues; vitamin D performs a neuroprotection role in the nervous system. The neuroprotective actions of vitamin D include: increasing vitamin D receptor (VDR) expression, control glutathione synthesis and nitric oxide synthase activity and induce neurotrophins such as nerve growth factor (NGF). VDR mediates cellular actions, and biological responses of the vitamin D.
OBJECTIVE:
To study the effect of VDR and NGF expression levels by vitamin D3 treatment in induced oxidative stress in primary cortical neuronal cultures.
METHOD:
Primary neuronal cultures were set up from the cortex region of neonatal rat’s brain. They were cultured for up to 72 h in the presence of 0.25μg/ml vitamin D3. These cells were exposed to 0.5 mM H2O2 for two hours before collecting cell pellet and medium for biochemical assays. Control and H2O2 treated cells were cultured in the absence of vitamin D3 treatment. Sandwich ELISA was used to study NGF expression. Western blotting and immunofluorescence of cultured cells were used to estimate the expression of VDR.
RESULTS:
Vitamin D3 treatment increased more significantly (P < 0.001) NGF levels with and without induced oxidative stress. Protein expression studies confirmed the positive correlation between VDR expression and vitamin D3 treatment after 72 h in culture. Moreover, pre-treating the cells with vitamin D3 before H2O2 exposure significantly increase (P < 0.05) VDR expression in comparison with the cells exposed to H2O2 alone.
CONCLUSION:
The neuroprotective effect of vitamin D3 against oxidative stress could be through up-regulating VDR and NGF levels.
Keywords
Introduction
Due to the high energy requirement and producing high amounts of ROS as a result, the central nervous system is more susceptible to oxidative stress [1]. The brain has an inadequate ability to detoxify ROS due to the lack of glutathione producing capacity of neurons. Therefore, the first cells to be affected by ROS are the neurons; as they are highly susceptible to oxidative stress [2]. Oxidative stress is considered to be one of the major causative agent for neuronal cells dysfunction and apoptosis, contributing to the pathogenesis of neurodegenerative diseases including: Alzheimer’s disease (AD), Parkinson disease (PD) and Amyotrophic lateral sclerosis (ALS) [3–6].
The active form of vitamin D has shown to be protective against oxidative stress [7, 8]. A study on rat cultured mesencephalic neurons showed that glutathione concentration increased significantly upon treatment with 1,25-(OH)2D3 [9]. Also, lower doses (1–500 nM) of 1,25-(OH)2D3 suppressed the ROS generation resulting from 1-methyl-4-phenylpyridium and L-buthionine-sulfoximine (BSO/MPP1) exposure [8].
Vitamin D receptor (VDR) mediates the cellular actions and biological responses of the 1,25(OH)2D3 hormone. Environment, genetics, and epigenetics play a crucial role in VDR regulation [10]. The selective presence of VDR in specific cell types supports the idea that gene expression of VDR is uniquely regulated. Studies have indicated that VDR gene expression can be up-regulated by the presence of l,25(OH)2D3 and direct stimulation by parathyroid hormone in bone and kidney cells mediated by both transcriptional and posttranscriptional mechanisms [11].
Studies in our lab indicated that treatments of cortical neurons with Vitamin D3 resulted in the protection of these neurons against induced oxidative stress by reducing lipid peroxidation and increasing glutathione levels [7]. The present study was designed to further look at the effect of vitamin D3 treatment in induced oxidative stress on the VDR protein expression as well as the effect on NGF levels.
Method
Setting up the primary cortical neuronal cell cultures
One week old Wister Albino rats (both genders) were used in this study. The rats were kept at a facility of King Saud university research Centre under the strict guidelines provided by the Experimental Animal Laboratory and approved by the animal care and use committee at the College of Applied Medical at King Saud University. All procedures dealing with animals were followed according to an ethically approved protocol (approval number KSU-SE-18-01). Primary cortical neuronal cells were set up using the same protocol in our lab as described by Al Johri et al. 2019 [7]. The cells were plated onto 12-well poly-lysine coated cell culture dishes (Millipore company) with a plating density of 5×106 cells/ml.
Treatment of cultured cerebral cortical neuronal cells
The cerebral cortex cells were divided into following Four groups in triplicate. Optimum dosage of Vitamin D3 and hydrogen peroxide dosage were used as described by AlJohri et al., 2019 [7].
Group 1: control cultures were grown for up to 72 h in the absence of any treatment.
Group 2: cells were grown for up to 72 h in the presence of 0.25μg/ml vitamin D3.
Group 3: cells were grown for up to 72 h in the presence of 0.25μg/ml vitamin D3, followed by treatment with 0.5 mM hydrogen peroxide for 2 h to induce oxidative stress.
Group 4: Cells were grown for up to 72 h without any treatment and then subjected to 0.5 mM hydrogen peroxide treatment for 2 h.
All Cultures were maintained in a humidified incubator with 5% carbon dioxide atmosphere at 37°C.
Measurement of Nerve Growth factor (NGF assay)
Rat β-NGF ELISA kit Cat no (RAB0381) was purchased from Sigma Aldrich U.S.A. 100μl of conditioned medium was used for each assay. The amount of NGF released into the culture medium (conditioned medium) was measured by the above chemokine sandwich ELISA kit according to the protocol provided by the company.
Western blotting
Total proteins from the cell extracts of cultures were extracted using reagent containing radioimmune precipitation assay (RIPA) buffer (Sigma Aldrich, USA) and 0.1 mM phenylmethanesulfonyl fluoride (PMSF). The proteins were precipitated by adding four times the sample’s volume of cold acetone and incubated at –20°C overnight.
25μg of reduced protein samples were separated in 12% SDS-PAGE at 100 volts for two hours. Proteins were transferred to nitrocellulose membrane and blocked 5% non-fat milk. Membranes were incubated in rabbit anti-phospho-VDR primary antibodies (pSer208 from Sigma Aldrich, USA) (LI-COR mouse a-Tubulin antibodies used as standard) and secondary antibodies (IR dye 800 anti-mouse from LI-COR Biosciences, USA). Bands were detected in the near-infrared region (800 nm) using Odyssey Imaging System (LI-COR Biosciences, USA). The western blots were quantified using image J software and expressed as relative absorbance of VDR expression with respect to alpha tubulin as VDR expression was normalized to α-tubulin expression in cells by calculating the normalization factor of each blot.
Immunofluorescence (IF) of VDR
IF was used to determine the cellular localization of the VDR on the primary cortical neuronal cells. Cells were fixed with 3.7% formaldehyde for 1 h, permeabilized by Triton X-100 followed by washing with deionized water and blocking with 2.5% bovine serum albumin (BSA). After that, they were incubated with the primary antibody for VDR (pSer208 from Sigma Aldrich) overnight at 4°C. After washing with PBS tween the cells were further processed with secondary antibody (IR dye 800 anti-rabbit from Li-COR bioscience) for 1 h. The cells were visualized by confocal microscopy (Nikon A1R).
Statistical analysis
The data were analyzed using statistical applications of Prism (version 7.0a). The data were statistically expressed in (mean±standard deviation). An independent sample t-test was performed to assess the difference between control and treated groups.
Comparison between control and treated groups were made using one-way analysis of variance (ANOVA). A probability value (p-value) less than 0.05 was considered to be significant.
Results
Effect of vitamin D treatment on Nerve growth factor (NGF) levels
NGF is a Neurotrophin essential for growth, proliferation, and survival of the nerve fibers in target neurons. In this study, we looked at the effect of vitamin D3 treatments with and without induced oxidative stress on the NGF levels. We found a significant reduction of (p < 0.001) of NGF by the cortical neuronal cultures with the treatment of H2O2 alone after 24 h, 48 h and 72 h (Fig. 1). The levels of NGF increased with vitamin D3 treatment alone as well as in induced oxidative stress by treatments with H2O2 in the presence of vitamin D3 significantly (p < 0.001) after 48 h and 72 h in culture as shown in Fig. 1.
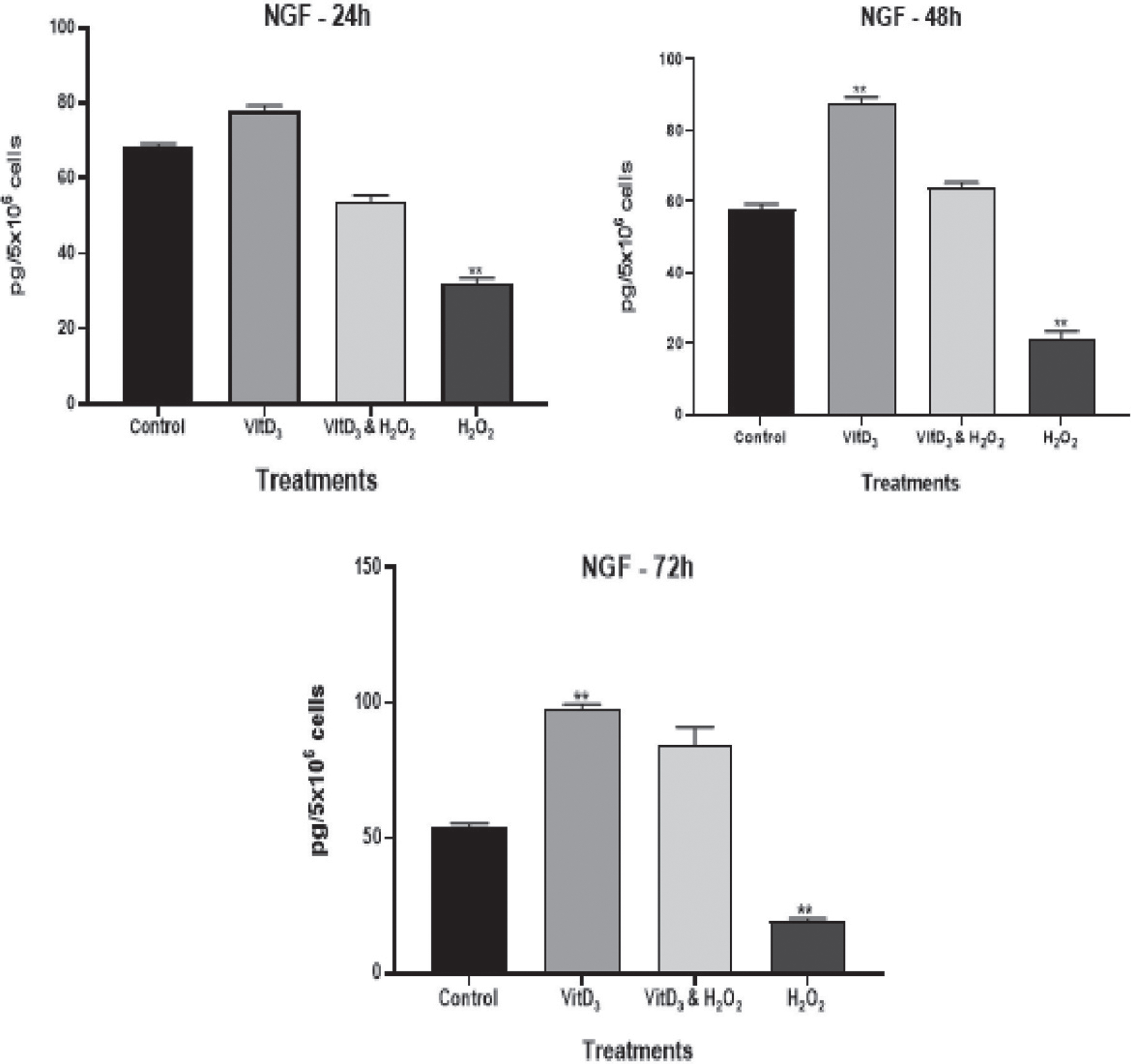
Effect of vitamin D treatment on nerve growth factor (NGF). Estimation of the effect of vitamin D3 treatment on nerve growth factor (NGF) at different time periods; (**) p-value < 0.001.
A 48 KDa VDR band was seen in all samples in western blot. Figure 2 showed immunoblots of VDR protein expression in cell lysate of cultured neuronal cells grown for 24, 48, and 72 hours respectively. These bands were further quantified using ImageJ (1.52a) program. The results, as shown in Fig. 2, showed no significant differences between different treatment groups at 24 h. However, when the neuronal cells were grown for 48 h, VDR protein expression was significantly (p≤0.05) more in the presence of vitamin D3 pretreated cells as compared to the cells exposed to H2O2 only (Fig. 2).
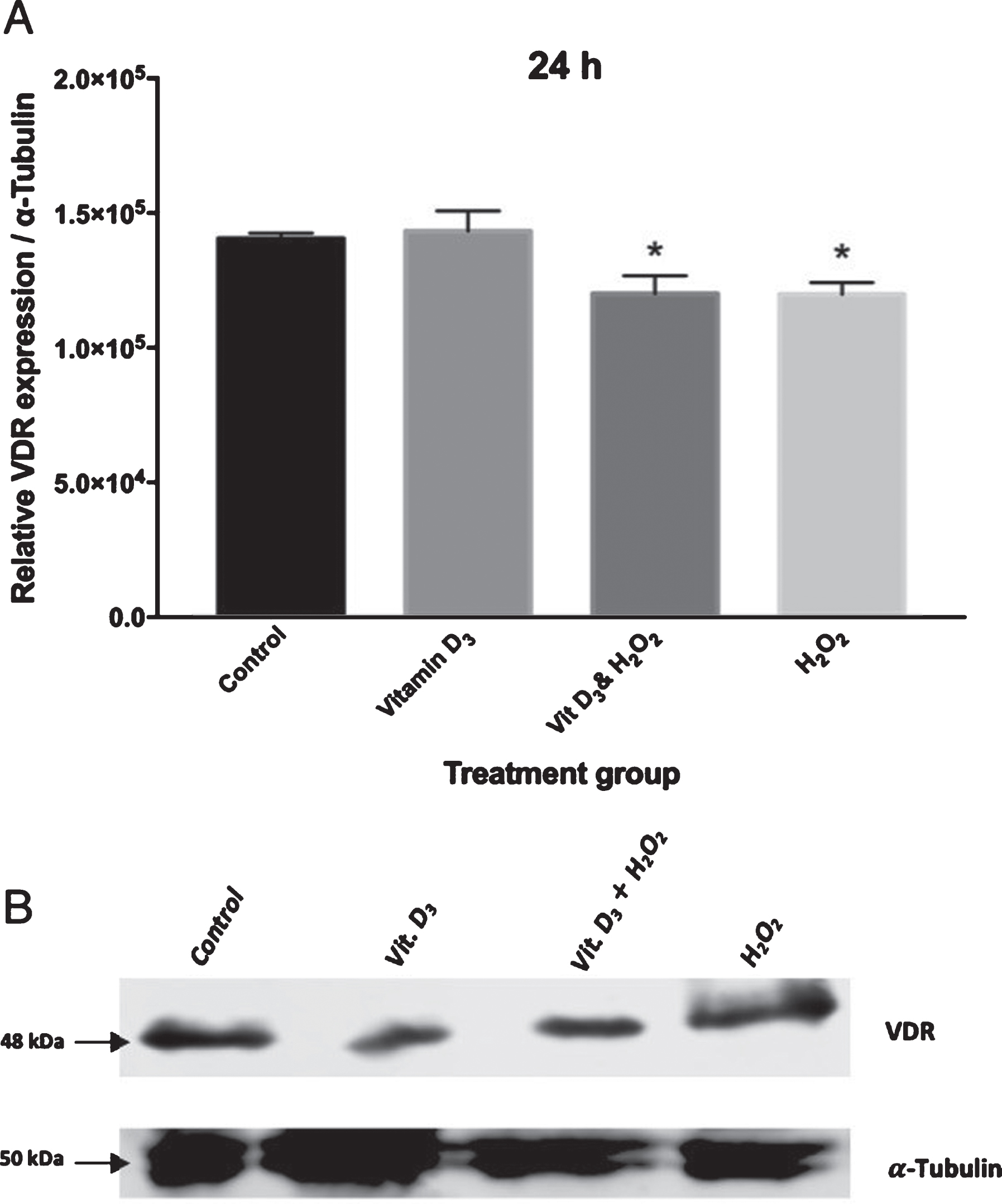
(A) Effect of vitamin D treatment on VDR by Western Blotting; in cells cultured and treated for up to 24 hours. (*) p-value <0.05 (B) Relative band intensity of VDR expression with respect to α-tubulin were quantified using ImageJ software. VDR expression was normalized by α-tubulin expression.
In 72 h cultures, H2O2 exposure significantly decreased VDR protein expression in comparison with the control and vitamin D3 pre-treated neuronal cells (p < 0.001) as well as the cells pre-treated with vitamin D3 before H2O2 exposure (p < 0.05).
Cell surface staining with the VDR antibody on the primary cortical neuronal membrane was observed. These primary cortical neuronal were mixed cells consisting of glial, astrocytes as well as neurons as observed in our earlier studies [12]. The presence of VDR on the neuronal membrane was observed before in other studies [13]. The control cells showed a lot of VDR localization on the cell surface (Fig. 3). The cells pretreated with Vitamin D3 in the presence and absence of induced oxidative stress by H2O2, showed positive IF staining in the Nucleus, cytoplasm and cell membrane (Fig. 3).

(A) Effect of vitamin D treatment on VDR by Western Blotting; in cells cultured and treated for up to 48 h hours. (*) p-value <0.05. (B) Relative band intensity of VDR expression with respect to α-tubulin were quantified using ImageJ software. VDR expression was normalized by α-tubulin expression.

(A) Effect of vitamin D treatment on VDR by Western Blotting; in cells cultured and treated for up to 72 hours. (**) p-value <0.001; (*) p-value <0.05. (B) Relative band intensity of VDR expression with respect to α-tubulin were quantified using ImageJ software. VDR expression was normalized by α-tubulin expression.

Cell surface immunofluorescence staining of primary cortical neural cells cultured and treated for 48 hours; stained with VDR antibodies and counterstained with Hoechst. The images were taken using confocal microscope at 63x magnification. The arrow indicates the nucleus of neuronal cell.
There is an active role of vitamin D in proliferation, differentiation, and immunomodulation. Studies reported the role of vitamin D in brain development and function in health and disease. The key aspects of vitamin D role in the nervous system include neuroprotection, neurotrophic, neurotransmission, and neuroplasticity [14, 15]. Correspondingly, vitamin D can affect neurotrophins expression, brain cells differentiation, antioxidant activity and the expression of genes and proteins involved in neuronal structure and metabolism [16].
The main mechanisms through which vitamin D confers neuroprotection is by up regulating glutathione levels as observed in other studies [7]. Other studies indicated that neuroprotection could be due to the effect of vitamin D in directly promoting the synthesis of nerve growth factor. It has been observed that vitamin D deficiency in neonatal rats leads to reduced NGF protein content and other genes involved in neuronal structure [17]. In this study, we looked at the effect of vitamin D3 treatment on the protein levels of NGF by Sandwich ELISA technique, and it indicated that vitamin D3 treatment could up-regulate the NGF. The treatment with H2O2 resulted in decreased production of NGF, but in the presence of vitamin D3, the toxic effect of H2O2 was reversed.
Recent research has established that the treatment of cultured primary cortical neurons with 0.25μg/ml of 1,25(OH)2D3 resulted in the greater protection of the neurons against the induced oxidative stress. H2O2 exposure resulted in lost 40% of cell viability, pre-treating the cells with 1,25(OH)2D3 resulted in improving cell viability [7, 18].
Western blot results showed that VDR expression was not significantly affected in control and treated samples after 24 h in culture. However, in cell lysates from 48 h, there was a significant positive correlation of vitamin D3 treatment and up-regulation of VDR expression in comparison to the cell lysate from the cells exposed to H2O2 alone.
Moreover, exposure of 72 h cultured cells to H2O2 alone significantly decreased VDR expression when compared to control and vitamin D3 treated cells. These results were indicative of the neuroprotective role of vitamin D against oxidative stress. Up-regulation of free radical scavenging and oxidative stress down-regulation was shown to be a result of VDR activation [19].
In general, therefore, it seems that the protective effects of vitamin D3 are positively correlated with more extended periods of vitamin D3 pre-treatment. The reason for this could be that during the initial 24 h culture periods, the cells were settling down and started to differentiate, proliferate and developing their neuronal connections. Once they were settled and established their dendritic and axonal connections, they responded better to the treatment of vitamin D3 by up-regulating VDR. These results support earlier studies, where the effect of glutamate toxicity on VDR expression in rat’s primary cortical neurons culture was investigated. This study revealed that VDR expression significantly increased after a brief exposure to glutamate in the presence of vitamin D3 [20].
The IF studies indicated the presence of VDR in the cytoplasm and cell membrane, which suggested its non-genomic role. It was reported that vitamin D performs its nongenomic role in brain by initiating intracellular and synaptic processes, which was further supported by in-vitro experiments on cerebral cortex of young rats, showing the involvement of vitamin D3 in both calcium and kinase dependent signaling pathways [21]. Since the presence of VDR in mitochondria was reported [22], the non-genomic role of vitamin D3 specially with respect to oxidative phosphorylation and mitochondrial biology is gaining a huge interest as most of the neurodegeneration is due to the mitochondrial dysfunction. Although the localization of VDR in the mitochondria has very limited evidence, recently the mitochondrial effect of vitamin D has been characterized in many cell types [22, 23]. The molecular role of VDR in mitochondria remained very ambiguous but various data revealed that VDR plays a central role in protecting from excessive oxidative phosphorylation and production of reactive oxygen species that leads to apoptosis. Future studies on the role of vitamin D3 in mitochondrial oxidative phosphorylation and mitochondrial membrane potential is warranted in order to understand the exact molecular role of vitamin D3.
Conclusion
In addition to calcium and phosphate homeostasis in peripheral tissues; vitamin D performs a neuroprotection role in the nervous system. The neuroprotective actions of vitamin D include: increasing vitamin D receptor (VDR) expression, control glutathione synthesis and nitric oxide synthase activity and induce the neurotrophins such as nerve growth factor (NGF). The present study reiterated the neuroprotective role of vitamin D3 against oxidative stress. The results of this study identify the up-regulation of VDR and NGF as a way of vitamin D protection of neurons against oxidative stress. These findings will be of interest in considering vitamin D as a potential therapeutic agent against neurodegenerative diseases.
Footnotes
Acknowledgments
Postgraduate Ebtesam Alsulami did this study as part of her Master’s thesis. The study was supported by a scientific and medical studies research center at King Saud University and a research grant for postgraduate studies King Abdul Aziz Science and Technology.