
Abstract
Background:
Large animal models, such as the transgenic (tg) Huntington disease (HD) minipig, have been proposed to improve translational reliability and assessment of safety, efficacy and tolerability in preclinical studies. Minipigs are characterised by high genetic homology and comparable brain structures to humans. In addition, behavioural assessments successfully applied in humans could be explored in minipigs to establish similar endpoints in preclinical and clinical studies. Recently, analysis of voice and speech production was established to characterise HD patients.
Objective:
The aim of this study was to investigate whether vocalisation could also serve as a viable marker for phenotyping minipigs transgenic for Huntington’s disease (tgHD) and whether tgHD minipigs reveal changes in this domain compared to wildtype (wt) minipigs.
Methods:
While conducting behavioural testing, incidence of vocalisation was assessed for a cohort of 14 tgHD and 18 wt minipigs. Statistical analyses were performed using Fisher’s Exact Test for group comparisons and McNemar’s Test for intra-visit differences between tgHD and wt minipigs.
Results:
Vocalisation can easily be documented during phenotyping assessments of minipigs. Differences in vocalisation incidences across behavioural conditions were detected between tgHD and wt minipigs. Influence of the genotype on vocalisation was detectable during a period of 1.5 years.
Conclusion:
Vocalisation may be a viable marker for phenotyping minipigs transgenic for the Huntington gene. Documentation of vocalisation provides a non-invasive opportunity to capture potential disease signs and explore phenotypic development including the age of disease manifestation.
INTRODUCTION
Huntington disease (HD) is the most frequent autosomal dominant neurodegenerative disorder and caused by a CAG triplet repeat expansion in the Huntingtin gene located on chromosome 4 [1]. The misfolded mutant protein Huntingtin (mHTT) causes widespread neuropathological changes that manifest in motor, cognitive and behavioural signs and symptoms [2, 3]. Motor symptoms are mostly characterised by involuntary movements, especially chorea and impairment of voluntary movements causing incoordination, bradykinesia and rigidity [4]. Cognitive decline is another hallmark of HD [5] including problems with attention, cognitive flexibility and memory [6–8]. Cognitive signs can occur prior to the onset of overt motor dysfunction [9]. Behavioural changes include loss of energy and initiative, poor perseverance, irritability and anxiety [10]. As a result of this triad of symptoms, patients with HD develop a voice and speech disorder known as hyperkinetic dysarthria [11]. Speech pathology in HD may manifest as phonatory dysfunction, articulation deficits and abnormalities in speech timing [12], which progress with disease [13, 14]. Although speech dysfunction has significant psychological implications and effects on quality of life, hyperkinetic dysarthria is still poorly understood in the HD population. Of note, dysarthria is often accompanied by dysphagia, a symptom often less appreciated in HD clinical evaluations, but of high clinical relevance: HD patients regularly experience aspirations in advanced stages of HD [15] and aspiration induced pneumonia is the most frequent cause of death observed in HD.
While several therapeutic approaches for HD are available or in development [16, 17], to date no disease modifying treatment has been established [18]. Despite numerous drugs suggesting efficacy for disease modification in preclinical studies, usually performed in rodents [19], all attempts to translate these findings to humans were unsuccessful. This may relate to the lack of standardisation, which is increasingly appreciated [20]. The need for improvements in translational science is recognised and receives growing attention in the field [21]. Recently, transgenic and knock-in large animal models have been proposed for preclinical safety and efficacy assessments with the expectation to provide higher reliability in translating preclinical findings to humans [22, 23].
Minipigs possess a very high genetic homology with humans and they have similarities in anatomy, physiology and metabolism to humans and a similar body weight [22]. Furthermore, they exhibit similar brain structure with a gyrated brain and have a life expectancy of 15–20 years presenting advantages to study a neurodegenerative disease longitudinally [24–28]. Recently, a transgenic (tg) Libechov minipig was created using lentiviral transduction as a new large animal model for HD at the Institute of Animal Physiology and Genetics in Libechov, Czech Republic [29]. This model exhibits a stable transmission of one copy of the HD mutation across several generations. Different motor, cognitive and behavioural tests have been established for this model [30] as well as protocols for Magnetic Resonance (MR) brain imaging and MR Spectroscopy [31]. Longitudinal repetitive performance of these assessments is well tolerated suggesting that long-term studies with this model are feasible, albeit no clinical phenotype was detected so far [32].
As voice and speech disorders have been documented in a majority of HD patients [33], examining grunting patterns in the tgHD minipig may provide important information about disease stage and progression. Disruptions in grunting may be detectable in the tgHD minipig model, due to similar HD-related pathophysiologic mechanisms [34]. Grunting is the most common type of vocalisation observed in pigs and it appears to be associated with investigatory behaviour and establishing contact with other pigs [35]. Furthermore, vocalisation in pigs has been identified as a sign of stress [36–38], which could indicate behavioural abnormalities. Hence, vocalisation in tgHD minipigs may serve as a readout for translational studies assessing natural history, disease progression or therapeutic benefit of novel treatment approaches. Therefore, in this study, we assess grunting during several defined cognitive and motor tests established for phenotyping this minipig model.We hypothesised that vocalisation can (1.) reliably be detected in minipigs, (2.) differentiate tgHD from wildtype groups and (3.) be reliably assessed longitudinally and thus possibly be applied in future studies involving phenotyping tgHD minipigs.
MATERIALS AND METHODS
Subjects
In the present study 14 transgenic (tgHD) and 18 wildtype (wt) female Libechov minipigs were included. The minipigs were bred at the Institute of Animal Physiology and Applied Genetics of the Czech Academy of Science (Libechov, Czech Republic) [29] and are all offspring of the same F1 generation. All animals participated in an ongoing long-term phenotyping study [32]. Animals were housed at the central animal facility of the University Hospital of Muenster (ZTE) in six age-matched groups with mixed wt and tgHD distribution as described before [30].
Assessments in the long-term phenotyping study were carried out biannually during a period of three years. The vocalisation assessment during the tests started when pigs were 2.5 years old, hence data from three visits (V) of the test battery (V1 to V3) for a total of 32 minipigs were available.
All research and animal care procedures were approved by the Landesamt für Natur- und Umweltschutz, North Rhine-Westphalia, Germany. All assessments were performed within the ZTE.
Experimental assessments
Vocalisation was documented for every run in a subset of assessments selected from the assessment battery, namely the tongue coordination test, the hurdle test and the colour discrimination test (blue box correct) including a reversal learning paradigm (yellow box correct). The colour discrimination test was conducted for assessing cognition; the minipigs were trained to open one of three coloured boxes (blue, yellow, red) that contained treats and were placed at the end of the walkway. In the first test stage, the blue box was correct while the others were locked and could not be opened. During the reversal learning phase, only the yellow box could be opened instead. The hurdle test assessed motor coordination of gait in a more challenging environment. Therefore, two hurdles at 13 cm height were arranged in the walkway. The tongue test is a motor test targeting the deficits in tongue coordination and tongue protrusion. The minipigs were trained to pick up treats from a board installed at the end of the walkway.
All tests were performed in a standardised environment and under standardised protocols (see Schramke et al. 2016 for complete experimental procedures). Videos of conducting the tests can be seen online [32].
The custom-made experimental setup consisted of two startboxes (SB A and SB B; 1.20 m×1.20 m) with a walkway (WW; 3 m×1.20 m) in between, connected by trap-doors, as shown in Fig. 1 (modified from Schramke et al. 2016). The setup was modified as needed for each assessment and described in detail elsewhere [30]. Any vocalisation occurring in the startboxes or in the walkway were coded as separate grunts, to separate active from passive phases. In the startbox, the minipigs had to rest a predefined time while the specific experimental procedures were conducted in the walkway. The walkway housed the coloured boxes, the tongue board and the hurdles for the discrimination, tongue and hurdle tests, respectively. The two discrimination tests and the hurdle test were repeated six times per assessment resulting in a total of 192 observations in each section (SB and WW) for each test. The tongue test included only three trials per assessment, resulting in 96 observations.

Custom-made setup used for the assessment battery, containing two startboxes (SB-A and SB-B) located at both ends of the walkway (WW) and connected by trapdoors. Depending on the particular test, the setup can be modified to the individual needs.
After finishing a trial, the minipigs returned to a startbox and waited until the next trial started.
Vocalisation was documented by an individual observer for every run in each of the different sections of the experimental setup (see Fig. 1). Binary coding recorded whether the minipig showed a vocalisation (1) or stayed quiet (0).
Statistics
Statistical analysis was performed using RStudio (Version 3.4, R Core Team 2017. R Foundation for Statistical Computing, Vienna, Austria) and SAS (SAS 9.2, SAS Institute Inc., Cary, NC, USA). As the data are dichotomous and not normally distributed, non-parametric analyses with frequency tables were conducted. The Fisher’s Exact Test for a 2×2 (genotype×vocalisation) data set was conducted to analyse general differences between the genotype groups. The test was carried out separately for the setup sections (SB, WW). McNemar’s Test for paired nominal data was used to detect differences between the three visits. The accepted level of significance was set to p < 0.05 [*p≤0.05; **p≤0.01; ***p≤0.001].
RESULTS
Discrimination tests
The occurrence of vocalisation by genotype in the startbox and the walkway for the discrimination test (blue box correct) and the reversal learning (yellow box correct) is shown in Fig. 2. For both tests, the minipigs had to wait in the startbox for 30 seconds prior to each trial. During this time, tgHD and wt minipigs demonstrated more vocalisation than in the walkway. However, analysis revealed an influence of genotype on vocalisation in the two setup sections. Across sessions, the tgHD minipigs vocalised significantly more than the wt minipigs during the discrimination test in the startbox (N = 192, V1– V3: p < 0.001) and along the walkway in visit 1 and 2 (N = 192, V1: p < 0.01; V2: p < 0.05), whereas for visit 3 a trend was observable (V3: p = 0.0624) (see Fig. 2A, B). In addition, at the reversal learning phase, a significant influence of the genotype on vocalisation was found in each visit in the startbox (N = 192, V1– V2: p < 0.001, V3: p < 0.05) and in the walkway (N = 192, V1– V2: p < 0.001), albeit for visit 3 the result is marginal significant (V3: p = 0.0611) (Fig. 2D, E).
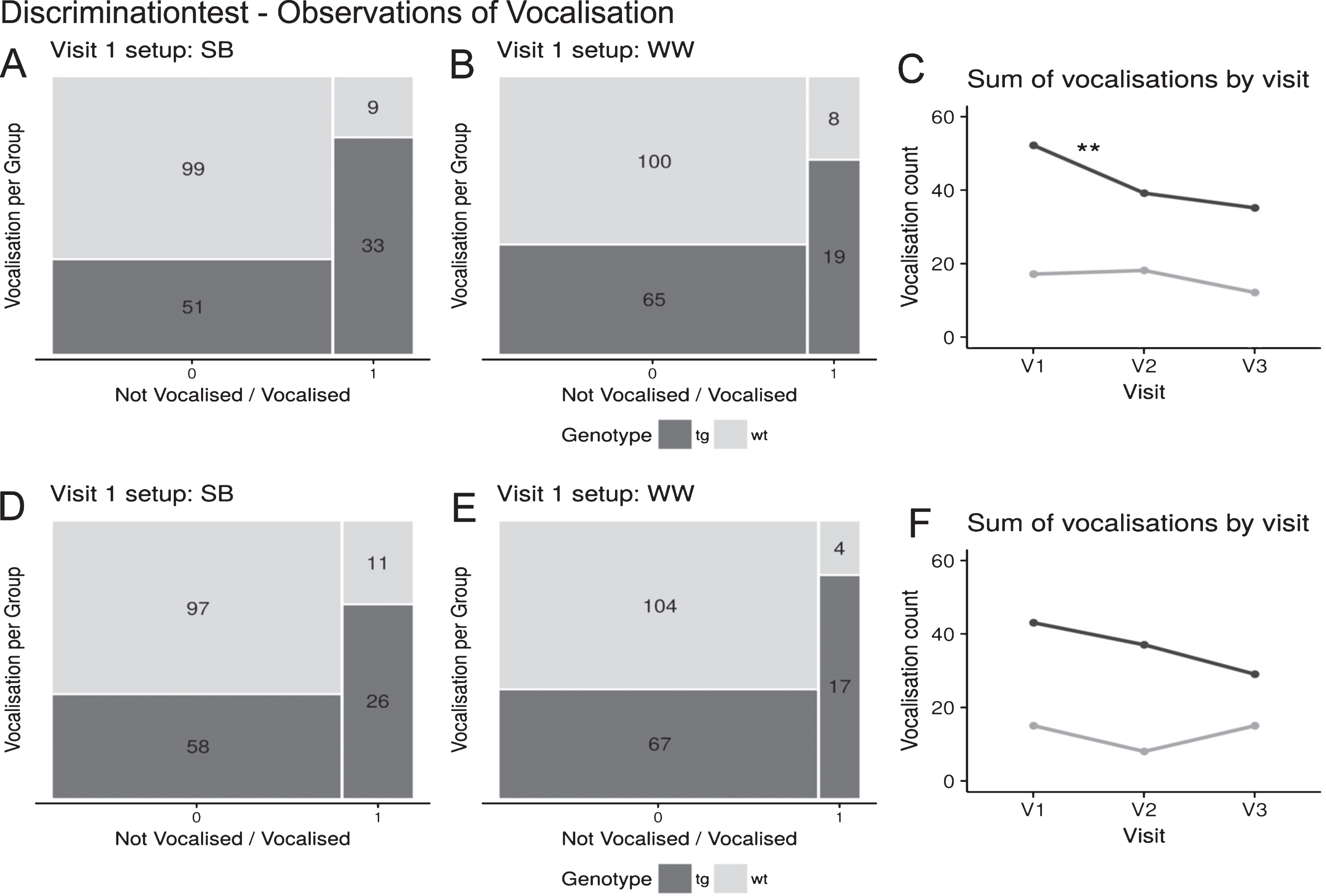
Results of the discrimination test (blue box correct) and the reversal learning (yellow box correct) for visit 1. Vocalisation (vocalised = 1, not vocalised = 0) distributed by genotype (tg, wt) for setup section SB (A, D) and WW (B, E). Results of the visit comparison (C, F) for the discrimination test and the reversal learning (vocalisation during SB and WW for each visit (V1– V3) compared between genotype groups (tg, wt)). (A-C) Discrimination test. (D-F) Reversal learning.
For the discrimination test and the other tests, respectively, the sum of vocalisations in each setup section for wt and tgHD can be seen in Table 1.
Observations of vocalisation
Total numbers of observations of vocalisation (vocalised = 1, not vocalised = 0) for all runs in each test, grouped by genotype (tg, wt) and setup type (SB, WW).
Discrimination tests: Differences between visits
Vocalisation diminished from visit 1 to visit 3 in the discrimination test for the tgHD group (see Fig. 2C). This was the result of a significant decrease in the number of vocalisations of tgHD minipigs between visit 1 (52 occurrences) and visit 2 (39 occurrences) during the discrimination test (McNemar’s Test, tg: N = 168, S = 7.3478, DF = 1; p < 0.01). No differences were found for the tgHD minipigs from visit 2 to visit 3 or for any comparison in the wt group during the discrimination test.
During the reversal learning, the wt minipigs tended to vocalise less in visit 2 than in visit 1. The trend of decreasing observations of vocalisation from visit 1 to visit 2 is shown in Fig. 2F (McNemar, wt: N = 216, S = 3.2667, DF = 1, p = 0.0707). No difference in vocalisation occurred from visit 2 to visit 3 for the wt minipigs and no differences were found for the tgHD minipigs in any reversal learning session comparisons.
Hurdle test
During the hurdle test the minipigs remained in the startbox for 10 seconds before the trap-door was opened and the pigs proceeded over the two hurdles in the walkway. A post hoc analysis was conducted to differentiate in which visit (V1 to V3) the vocalisation frequencies distinguished between wt and tgHD minipigs. The difference in the vocalisation observations between tgHD and wt subjects was present during visit 1 (SB: N = 192, p < 0.001; WW: N = 192, p < 0.01) (see Fig. 3A, B). Vocalisation occurred more often in the walkway (V1, tg and wt: 32 observations) than in the startbox (V1, tg and wt: 22 observations), mainly driven by the wt minipigs increased vocalisation during visit 1 (see Table 1). In visit 2 and 3 these results could not be confirmed.

Results of the hurdle test for visit 1 for the SB (A) and the WW (B) and for the visit comparison (C) (vocalisation (0, 1) distributed by genotype (tg,wt)).
Hurdle test: Differences between visits
Vocalisation diminished from visit 1 to visit 3 in the hurdle test for the tgHD group. During the hurdle test the wt minipigs demonstrated consistent vocalisations while the tgHD minipigs showed progressively decreased grunting. Vocalisation decreased from 42 grunts in visit 1 to 14 grunts in visit 2 (McNemar, tg: N = 168, S = 20.6316, DF = 1, p < 0.001) and just 4 vocalisations in visit 3 (McNemar, tg: N = 168, S = 6.2500, DF = 1, p < 0.05) (see Fig. 3C).
Tongue test
The distribution of vocalisation by genotype for the tongue test in visit 3 is shown in Fig. 4. In the startbox, where the minipigs had to wait for 30 seconds, the tgHD minipigs vocalised significantly more than the wt minipigs (V3; SB: N = 96, p < 0.05). In the walkway, the wt minipigs vocalised more often than they did in the startbox, hence no significant differences between tgHD and wt minipigs was detected in the walkway. For visit 2 and 3 no differences between genotypes were found for any of the setup sections.

Results of the tongue test for visit 3 and for visit comparison (C). Vocalisation (0, 1) distributed by genotype (tg, wt) for each setup section, SB (A) and WW (B).
Longitudinally, no differences in vocalisation for tgHD and wt minipigs occurred for any tongue test session comparisons (see Fig. 4C).
The distribution of vocalisation across minipigs is shown in Fig. 5. The total count of runs with vocalisation of each minipig across all tests and both setup sections (SB, WW) is plotted against the visit.
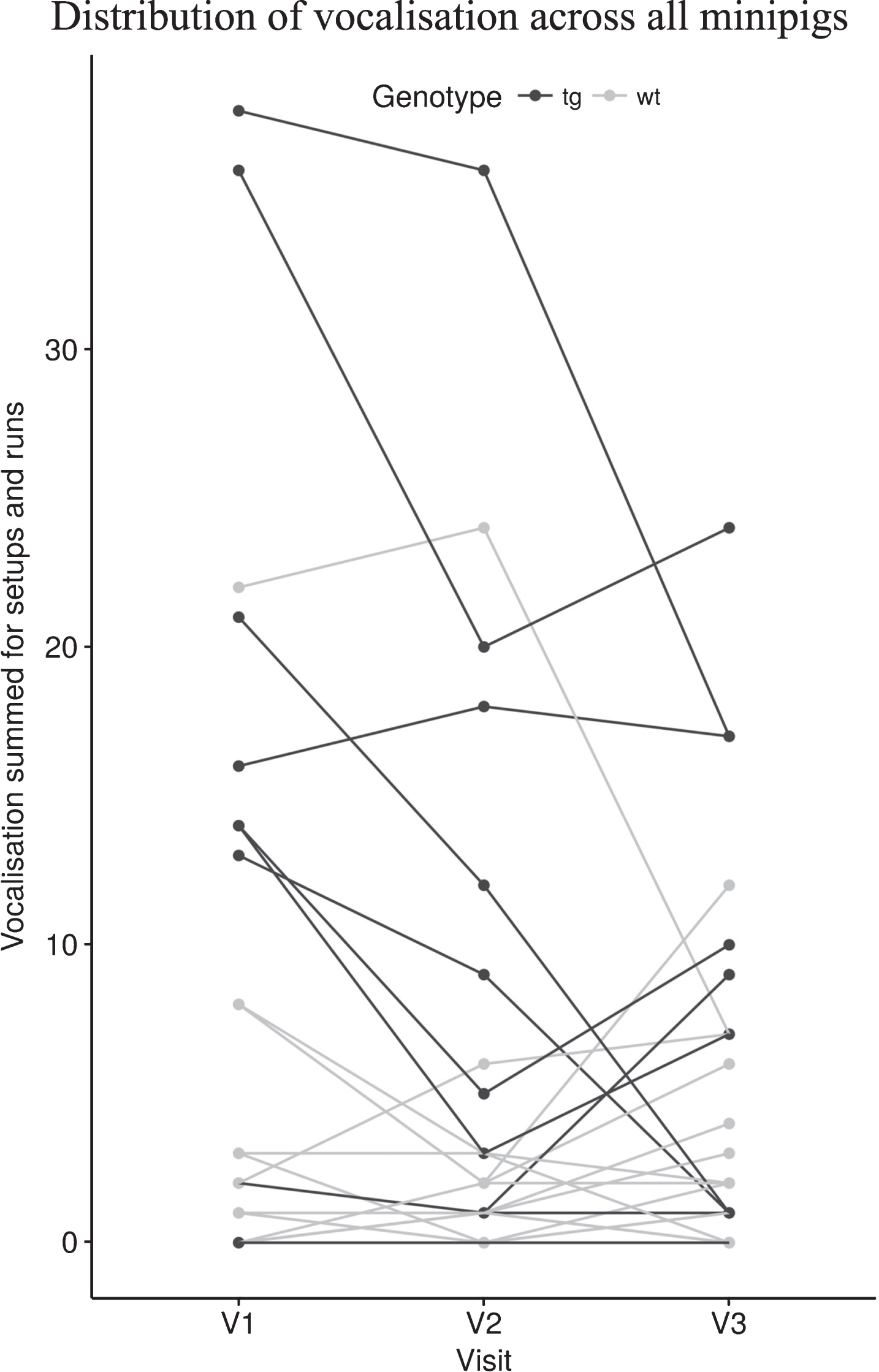
Total counts of vocalisation of each minipig summed for setup sections (SB,WW) and runs plotted against the visit (V1-V3).
DISCUSSION
In this study, significant differences in the distribution of vocalisation between tgHD and wt minipigs were demonstrated while other behavioural measures assessed previously did not detect phenotypical abnormalities [32]. Specifically, the results showed that tgHD minipigs have more frequent vocalisations than healthy wt minipigs, mainly during the discrimination test assessing cognitive function. In addition, the continued vocalising during repeated testing suggests that the tgHD minipigs did not adapt to the testing in the same manner as the wt minipigs as vocalisation may be seen as an indicator of arousal. These differences highlight the potential benefits of vocalisation as an outcome measure in future studies and suggest that vocalisation may serve as a viable behavioural marker for phenotyping tgHD minipigs.
Vocalisation was reliably expressed during the tests and can easily be documented in a non-invasive way. Vocalisation serves as an important communication tool for minipigs and is part of the natural behavioural repertoire of pigs. For example, Marchant, Whittaker, and Broom (2001) reported that social isolation of domestic pigs led to a number of sounds produced by the isolated pig [35]. In addition, pigs grunt more during stressful situations characterised by social isolation without any visual and tactile social contact [39]. The fact that the minipigs did not see other animals during the behavioural tests in this study is a form of social isolation, as the minipigs are housed in groups with eye contact to proximate groups. Such separation may have caused arousal, which was expressed by grunting, especially in the startbox, where the minipigs must wait. These findings are reasonable give the function of vocalisation or grunting associated with contact calls [35]. High-frequency vocalisations are supposed to be a sensitive indicator of pigs emotions during anticipation of negative events [40]. Anticipation of the isolation in the startbox could be a negative event as the results of this study show that the frequency of vocalisations decreases as the sessions increase.
Grunting is the most common sound produced when a pig is excited and is a sign of investigatory behaviour while foraging. As the behavioural tests were reward-based, it is possible the minipigs anticipated the reward (food) and became excited. Vocalisation shown during a rewarding event could be indicative of positive emotions [39] and anticipation of a positive event like a food reward may result in positive arousal [41].
Communication sounds are crucial for intra-species interactions [42] and thus, changes in vocalisation have a strong impact on the social group structure. It is well known that pigs produce different vocal responses to mental and physical stressors [38], so the type of vocalisation may give good and further insights to the internal state of anindividual. In several studies, pig vocalisation was measured under different kinds of stress [36, 44]. As learning and retrieval of new situations are impaired in HD patients [3], we hypothesised that tgHD minipigs would exhibit a higher level of stress during testing and not adapt to the tests in the same way. Indeed, we found increased vocalising in the tgHD minipigs across conditions. However, vocalisation occurrences decreased from visit 1 to visit 3, suggesting that while there may be an initial impairment in stress regulation, the tgHD minipigs were able to learn and adapt to the testing conditions. For the discrimination test, such a learning effect was detected longitudinally over a period of three years [32].
Recent studies suggest that speech has the potential of serving as an early and objective indicator of HD onset and disease progression [13, 14]. Likewise, we have shown vocalisation may represent an early indicator of disease in tgHD minipigs. Processing vocalisations is related to the cerebral cortex [42], which is known to be affected by HD [45] with functional impairment of cortical neurons [46]. Meaningful vocal production requires central nervous system (CNS) integrity [14] as seen when minipigs use grunts for signalling, social communication and for expressing their emotions [47].
We acknowledge some limitations of the analyses. In this study, we only focused on whether the minipigs vocalised during the test or not, we did not consider the specific kind of vocalisation. A variety of sounds can be produced by pigs, but vocalisation is mainly distinguished by shorter low frequency calls and long high frequency calls [38, 48]. For further studies, acoustic analysis by using microphones may provide objective and more quantitative information about the phonatory characteristics.
The Libechov tgHD minipig model provides high potential as a large genetic animal model for HD research. The porcine Huntington gene has high genetic homology to humans [29] and the minipigs possess a large brain and cerebral gyri [22]. The 20 year life span allows for neural degeneration to occur over an extended period of time and facilitates the execution of longitudinal studies. Using tgHD minipigs may close the translational gap between preclinical research in rodents and clinical trials in humans [49]. Minipigs acquire a high level of accuracy in different cognitive tasks [50] and tests assessing motor and cognitive abilities are well tolerated by the Libechov minipigs and can be performed in a consistent way [32]. Furthermore, conducting behavioural tests is relevant to understand the neurobiological processes responsible for behavioural changes caused by a neurodegenerative disease such as HD [25].
We conclude that vocalisation assessments may provide a functionally relevant measure in preclinical studies in HD minipigs. In future preclinical study-designs using the tgHD minipig, vocalisation should be further explored as an easy to obtain and sensitive behavioural measure.
CONFLICT OF INTEREST
The authors have no conflicts of interest to report.
Footnotes
ACKNOWLEDGMENTS
This study was funded by the CHDI foundation (![]() ) and by relevant private donations of families affected by HD. We thank the team of the Zentrale Tierexperimentelle Einrichtung (ZTE) of the University Hospital of Muenster, Muenster, Germany, for continuous support.
) and by relevant private donations of families affected by HD. We thank the team of the Zentrale Tierexperimentelle Einrichtung (ZTE) of the University Hospital of Muenster, Muenster, Germany, for continuous support.
The George-Huntington-Institute has received grant support from the CHDI Foundation to conduct the work reported in this study.