
Abstract
Background:
Transgenic sheep are currently the only large animal model of Huntington’s disease expressing full-length mutant human huntingtin. These transgenic sheep provide an opportunity to test adeno associated virus (AAV) therapies directly targeting the huntingtin gene. A recent study demonstrated that self-complementary (sc) AAV with artificial miRNA against human huntingtin reduced mutant human huntingtin in caudate and putamen after a single injection near the internal capsule.
Objective:
To identify an AAV serotype among AAVrh8, AAV9 and AAVrh10 with the highest neuronal uptake and distribution, with no obvious cell loss in the neostriatum of the sheep.
Methods:
We tested AAVrh8, AAV9 and AAVrh10 by stereotactic direct unilateral injection into the neostriatum of sheep, near the internal capsule. Four weeks after administration, we examined the viral spread and neuronal uptake of each serotype of AAV containing GFP. We compared single stranded (ss) and scAAVs. Further, we measured the distribution of AAVrh8 and AAV9 to a variety of tissues outside the brain.
Results:
Sc AAV9 had the best combination of neuronal uptake and distribution throughout the neostriatum. scAAVrh10 demonstrated good spread, but was not taken up by neurons. scAAVrh8 demonstrated good spread, but had less neuronal uptake than AAV9. Six hours after convection-enhanced administration to the neostriatum, both AAVrh8 and AAV9 viral genomes were detected in blood, saliva, urine, feces and wool. By four weeks, viral genomes were detected in wool only. Administration of AAVrh8, AAV9 and AAVrh10 was not associated with loss of neostriatal, medium spiny neuron number as measured by DARPP32 immunohistochemistry.
Conclusions:
Altogether, we found scAAV9 had the best neuronal uptake and spread, showed no loss of neurons at one-month post-injection, and was not measurable in body fluids one month after injection. This information will guide future clinical experiments requiring brain injection of AAV for therapeutics for gene or miRNA deliveries in sheep transgenic for the human huntingtin gene.
INTRODUCTION
The therapeutic potential of adeno-associated virus (AAV) in Huntington’s disease, and other genetic causes of neurodegeneration, is its ability to deliver RNA interference (RNAi) constructs to silence mutant genes in neurons for many years [1, 2]. AAV primarily resides in the nucleus and integrates sparingly into the genome [3]. There are many different AAV capsid serotypes [3], each with particular cellular tropisms, distributions, and transduction efficiencies [4]. Several AAV serotypes, most notably AAV2, have been used in clinical trials for gene therapy of inherited diseases [5]. The AAV serotypes appear to be safe and exhibit neuronal tropism after administration into mouse and non-human primate brains [6, 7], but species selective neuronal uptake in sheep has not been reported. The rationale to study sheep is based on the availability of a transgenic HD sheep model with a full-length human mutant huntingtin with 73 CAG repeats [8]. RNAi with artificial miRNA against mutant human huntingtin in the transgenic sheep model highlights the usefulness of AAV-miRNA based lowering of mutant huntingtin in a large animal [9]. In this study, we tested three AAV serotypes, AAVrh8, AAV9 and AAVrh10, for neuronal uptake, distribution and safety in the brains of sheep, which, like the human neostriatum, separates into the caudate and putamen. We had three goals in this study: we compared the extent of neuronal uptake and distribution of each AAV serotype delivered by a convection-enhanced method into the sheep neostriatum; we examined each AAV for evidence of neuronal loss; and we studied distribution of AAV administered into the striatum to compartments outside the brain.
METHODS
Experimental animals
Male Dorset sheep were obtained from Archer Farms, Inc. (Darlington, MD) and Earle M. Parson & Sons, Inc. (Hadley, MA) and housed under standard conditions with access to food and water ad libitum in the Department of Animal Medicine in the University of Massachusetts Medical School (UMMS). The sheep were maintained and used as approved by the UMMS Institutional Animal Care (docket#2288-11 to N.A.), in accordance with the institution’s program of animal care (accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International). All animals were weight-matched at time of surgery (approximately 40lbs) and were housed alone post-surgery.
AAV preparation
rAAV vector plasmid carrying a vector genome with the expression cassette for reporter gene enhanced green fluorescent protein (GFP) and flanked by AAV2 ITRs is co-transfected into HEK 293 cells with a packaging plasmid and adenovirus helper plasmid. The packaging plasmid will express regulatory proteins (rep2) of AAV2 and AAV9, AAVrh.8 and AAVrh.10 capsid proteins (cap9, cap rh.8 and cap rh.10), which will excise the recombinant genome from the rAAV vector plasmid, replicate the genome, and package the genome into AAV virions. Adenovirus serotype 5 E1, E2a and E4 proteins, and VA I and II RNAs expressed from the adenovirus helper plasmid will provide helper functions essential for rAAV rescue, replication, and packaging. The packaged rAAV is actually a chimeric virus in which the rAAV genome with the transgene cassette flanked by AAV2 ITRs is packaged with the capsids of AAV9, AAVrh.8 and AAVrh.10 respectively. The recombinant viruses are purified by standard CsCl gradient sedimentation method and desalted by dialysis [10]. The vectors are quality control tested by qPCR titration for DNase resistant vector genome concentration, by silver-stained SDS-polyacrylamide gel analysis to establish the purity of each lot, and by electron microscopic analysis to independently test full/empty virus ratios [10].
Anesthesia
Anesthesia was performed and continuously monitored by qualified members of the Animal Medicine Department in accordance to all applicable rules and standards. Animals were pre-medicated with a combination of buprenorphine (0.01 mg/kg), acepromazine (0.05 mg/kg), and glycopyrrolate (0.01 mg/kg) intramuscularly, and then induced with ketamine (6 mg/kg) and diazepam (0.3 mg/kg) intravenously. A dose of ketoprofen (2.2 mg/kg) was administered subcutaneously for additional anesthesia. Anesthesia was maintained during the entire procedure with 2% isoflurane in oxygen and the animal was ventilation mechanically. Physiological monitoring included heart rate, arterial oxygen saturation, body temperature, and end tidal CO2 during the procedure. Veterinary staff of the Department of Animal Medicine provided all post-operative care.
Surgical procedure
The animals were transferred to the operating suite and prepared for sterile surgery. They rested in the sphinx position (ventral recumbency) on foam cushion using a standard large animal operating table. The Kopf MR compatible large animal stereotactic frame (1430) was used to secure the head using standard tapered ear bars and non-invasive superior orbital rim fixation with a snout bar to stabilize.
The dorsum of the skull was shaved, prepped and draped and a slightly curvilinear transverse skin incision was made, exposing both coronal sutures. A burr hole (<0.5 cm diameter) was created approximately 1 cm rostral and lateral of the bregma. The MRI-Intervention infusion cannula was positioned in the micro-manipulator and primed with the infusate to insure no air was injected into the brain. The depth was measured from the inner table of skull. The dura was opened as a small slit through which the cannula passed and fully occluded the dural opening to prevent cerebrospinal fluid (CSF) leakage during the infusion. The cannula was slowly advanced to the desired depth and the infusion started at a rate of 3μl/min for a total of 300μl using a syringe pump. Ten min after completion of the convection enhanced delivery (CED, about 20 min) the cannula was withdrawn and the dural defect covered with a small patch of peri-cranial tissue. The skull defect was closed with a bone wax plug to prevent sub-galeal CSF leakage. The scalp was closed using an absorbable suture in the galea and glue to seal the skin margins.
Post-surgical imaging and recovery
Following closure of the scalp the ear bars were removed and the orbital clamps released. While under the general anesthesia the animals were transported to the adjacent MRI room where the immediate post-op study was performed. To visualize the tissue distribution of the infusate using MR imaging, infusates were mixed with 2.0 mmol/l gadolinium (Gd) (Supplementary Figure 1).
The MRI-Intervention cannulas, specifically designed for convection-enhanced-delivery (CED) first described by Oldfield et al., allow for the direct administration of a desired substance under a pressure gradient into the brain parenchyma independent of molecular size or blood-brain-barrier permeability. The stepwise design of the cannula tip resist backflow along the cannula exterior and thus enhances the widespread radial distribution of the infusate dictated predominantly by the rate and volume infused. The CED pressure gradient allows for the distribution of large agents, like AAV, to distribute in high concentrations over relatively large tissue volumes.
The Department of Animal Medicine supervised the post-operative care: alertness and general behavior, blood pressure, heart rate, weight, feeding frequency, defecation and urination frequency.
Necropsy for histology and bio-distribution
After day twenty-eight post-injection, sheep were administered 200–500 U Heparin/kg IV (jugular), then euthanized with 150 mg/kg sodium pentobarbital intravenously. For histology, sheep were perfused intracardially with 8 L of PBS followed by 8 L of 4% paraformaldehyde (PFA) at a pressure of 400–500 psi. For bio-distribution, sheep were perfused intracardially with 8 L of PBS. Then, tissues were harvested in a manner to avoid cross-contamination, snap frozen in liquid nitrogen and stored at –80°C.
Tissue collection for histology
After necropsy, brains were post-fixed in 4% PFA for 24 h. The left and right hemispheres of the brain were separated and transferred to 30% sucrose for a minimum of 14 days. 24 h prior to slicing the brain, the striatum was isolated from the brain, covered in Optimal Tissue Cutting (OCT) Compound (Tissue –Tek) and frozen at –20°C. Coronal 40μm sections of brain were made with a sliding microtome (Reichert-Jung Tetrander sliding microtome) through the striatum. Each section was floated in 0.01% Sodium Azide in 1X PBS and stored at 4°C.
Vector bio-distribution
Blood, saliva swabs, urine, wool and stool samples were collected at baseline, 6-h post-injection, day one, day seven, and day twenty-eight time points. At each time point, sheep were moved to clean stalls for sampling to obtain fresh, uncontaminated samples. Samples were stored at –80°C until genomic DNA was extracted. Genomic DNA (gDNA) was isolated from saliva swabs, urine and wool using a DNeasy blood and tissue kit (Qiagen, Inc., Valencia, CA, USA) with modifications made to the manufacturer’s instructions. Saliva swabs were soaked in 1x phosphate buffered saline for 10 min at room temperature before proceeding with the extraction process. Urine samples were processed according to the manufacturer’s protocol using double the amount of sample suggested. Wool was cut into 1 cm square sections that were then used for gDNA extraction by lysing the sample overnight at 56°C in 300μL Buffer ATL, 20μL proteinase K and 20μL 1M DTT prior to proceeding with the extraction process. gDNA was extracted from feces using the QIAamp DNA stool mini kit (Qiagen, Inc., Valencia, CA, USA) according to manufacturer’s instructions.
At necropsy, blood and tissues were harvested in a manner to avoid cross-contamination, snap frozen in liquid nitrogen and stored at –80°C until genomic DNA was extracted per a previously described protocol [11]. Genomic DNA was isolated from blood, gonad, muscle, adrenal gland, heart, liver, cerebellum, cortex and striatum using a DNeasy blood and tissue kit (Qiagen, Inc., Valencia, CA, USA) according to the manufacturer’s instructions.
Genomic DNA concentrations were determined using a Nanodrop 1000 Spectrophotometer (Thermo Fisher Scientific) according to manufacturer’s instructions. rAAV genome copies in the gDNA were quantified by real time PCR using an ABI 7900 HT sequence detection system (Applied Biosystems, Carlsbad, CA, USA) and results were analyzed using the SDS 2.3 software. Briefly, primers and probe were designed to the CB promoter of the vector cassette. A standard curve was performed using plasmid DNA containing the same CB promoter target sequence. PCR reactions contained a total volume of 100μL and were run at the following conditions: 50°C for 2 min, 95°C for 10 min, and 45 cycles of 95°C for 15 s and 60°C for 1 min.
DNA samples were assayed in triplicate. To assess PCR inhibition, the third replicate was spiked with target plasmid DNA at a ratio of 100 copies/μg gDNA. A sample that contained greater than or equal to 100 copies/μg gDNA was considered positive for vector genomes. A sample that contained fewer than 100 copies/μg gDNA was considered negative for vector genomes. We were not confident with the reproducibility of values less than 100 copies/μg gDNA and therefore made the arbitrary cut-off at that level. If less than 1μg of gDNA was analyzed to avoid PCR inhibition as determined for each tissue, the vector copy number reported was normalized per μg gDNA and the plasmid spike in was reduced to maintain the ratio of 100 copies/μg gDNA.
Immunohistochemistry for GFP, DARPP32, and NeuN+GFP
A sheep atlas (Welker Wisconsin Collection) was used as a guide to brain sectioning. To visualize DARPP32 or GFP positive cells, every twentieth 40μm brain section was incubated for 3 min in 3% hydrogen peroxide in 1X PBS, 20 min in 0.5% Triton-X-100 in 1X PBS (DARPP32 only), and 4 h in 1.5% normal goat serum (Vector Laboratories S-1000) in 1X PBS. Sections were incubated in anti-DARPP32 (Abcam ab40801; 1:10000 dilution) or anti-GFP (Life Technologies G10362; 1:5000) in 1.5% normal goat serum at 4°C overnight. Sections were then incubated in biotinylated anti-rabbit secondary antibody (Vector Laboratories BA-1000; 1:200 dilution) in 1.5% normal goat serum diluted in 1X PBS for 10 min. The sections were incubated with 2% A and 2% B reagents from the Vectastain ABC Kit (Vector PK6100). The reaction was visualized by 3,3’-diamainobenzadine (DAB) with Metal Enhanced DAB Substrate Kit (Pierce).
To visualize NeuN+GFP cells, every twentieth 40μm brain section was incubated for 3 min in 3% hydrogen peroxide in 1X PBS, 20 min in 0.5% Triton-X-100 in 1X PBS, and overnight in 1.5% normal goat serum (Vector Laboratories S-1000) in 1X PBS at 4°C. Sections were incubated in anti-GFP (Life Technologies G10362; 1:2000 dilution) and 1.5% normal goat serum at 4°C for 1 h, followed by incubation in anti-NeuN (Millipore MAB377; 1:1000 dilution) in 1.5% normal goat serum 4°C for 1 h. Staining was visualized using a goat anti-mouse IgG secondary antibody, Alexa Fluor 594 (Life Technologies A11005:1:1000 dilution), and a goat anti-rabbit IgG secondary antibody, Alexa Fluor 488 (Life Technologies A11034; 1:1000 dilution) fluorescent secondary antibody for 40 min.
Quantification of histology
To measure safety of AAV delivery to the striatum of a large animal, the number of DARPP32 positive cells was counted on both the ipsilateral and contralateral side of the brain. 20X images of the striatum were captured with a Nikon Eclipse E600. To be consistent between brain sections, images were taken by starting in the medial, dorsal edge of the striatum and the microscope stage was moved 0.5 cm towards the ventral end of the striatum. Once the ventral end was reached, the microscope stage was moved 0.5 cm toward the ventral side of the striatum, then 0.5 cm towards the dorsal end of the striatum until a maximum of 10 images were collected. Each image was assigned a random number to eliminate bias. ImageJ (NIH) was used to facilitate manual counting of the cells. The total number of DARPP32 counted cells in the striatum of one animal was normalized to striatal volume to represent the total number of DARPP32 cells/mm3. The contralateral hemispheres were treated as a control.
Another method to evaluate safety of AAV is to measure the cross sectional area of DARPP32 positive cells. 40X images of the striatum were captured with the same method described above. NIS-Elements BR 3.10 was used to measure the area of individual DARPP32 cells. To ensure cells in the same plane were measured, the unstained nucleolus needed to be visible for the cell to be measured. The cross sectional area of each measured cell in the striatum of one animal was averaged together to represent the mean area of DARPP32 cells. The contralateral hemispheres were treated as a control.
To assess the neuronal uptake of AAV, fluorescent NeuN+GFP staining was quantified on a Nikon Eclipse E600 with a Chiu Technical Corporation Mercury 100-W lamp. Under 60X, we counted the number of NeuN cells and the number of GFP cells that co-localized with those NeuN cells. The data were represented as the percent of NeuN cells that are GFP positive.
To measure the spread of AAV, GFP (DAB) stained sections were scanned (Epson perfection V750 pro, 1200dpi) on to the computer, along with DARPP32 (DAB) sections. ImageJ (NIH) was used to measure the area of DARPP32 (the striatal volume) and the area GFP as described above. The volume of DARPP32 and GFP was calculated by determining the distance between each section and the thickness of each section, and then multiplying by the measured area. The data were represented as the percent of striatum that is GFP positive.
Statistics
One-way analysis of variance (ANOVA) and unpaired student’s T-test was performed using GraphPad Prism6 for percent co-localization and GFP spread analysis. Post hoc was performed using the Tukey’s method to control for multiple comparisons. Unpaired student’s T-Test was performed using GraphPad Prism 6 for stereological counting of DARPP32 labeled cells, cross-sectional area of DARPP32 labeled cells, and striatal volume.
RESULTS
scAAV9 transduced the highest percentage of striatal neurons, compared to AAVrh8 and AAVrh10
To determine the best neuronal uptake between AAV9 and AAVrh10, each serotype was studied as single stranded (ss) AAV or self-complementary (sc) AAV. The AAV serotypes shared the same CβA promoter and expressed GFP. In sheep brain, the neostriatum has three structures: anterior striatum, caudate and putamen. 100μl of each AAV serotype was injected unilaterally into a space between the right caudate and putamen, at the internal capsule (a single injection). After four weeks, brains were harvested and immunohistochemistry or immunofluorescence was performed. To determine co-localization of a particular AAV serotype in neurons, we performed immunofluorescence for NeuN and GFP. We used histological and stereological evaluations to measure extent of co-localization of NeuN and GFP through the striatum. Neuronal uptake was low with all of the 100μl volumes of AAV highest serotypes (under 20%; Fig. 1A). Since injection volume could be a determinant of neuronal transduction, we tested an injection volume of 300μl of scAAV9. Neuronal uptake was significantly high in the neostriatum with 300μl of scAAV9 (40% of striatal neurons; Fig. 1B, Supplementary Figure 2). Finally, to confirm that 300μl scAAV9 achieved the highest transduction of striatal neurons possible, we injected a third AAV, AAVrh8, at a volume of 300μl. 300μl injection of AAVrh8 transduced 23% of striatal neurons, a significant reduction compared to sc AAV9 300μl (Fig. 1B). Considering that both scAAVrh10 100μl and ssAAVrh10 100μl had poor neuronal transduction compared to scAAV9 100μl and ssAAV9 100μl, we chose not to pursue testing AAVrh10 at a higher volume. Together, these data suggest that scAAV9 300μl had the highest neuronal transduction.
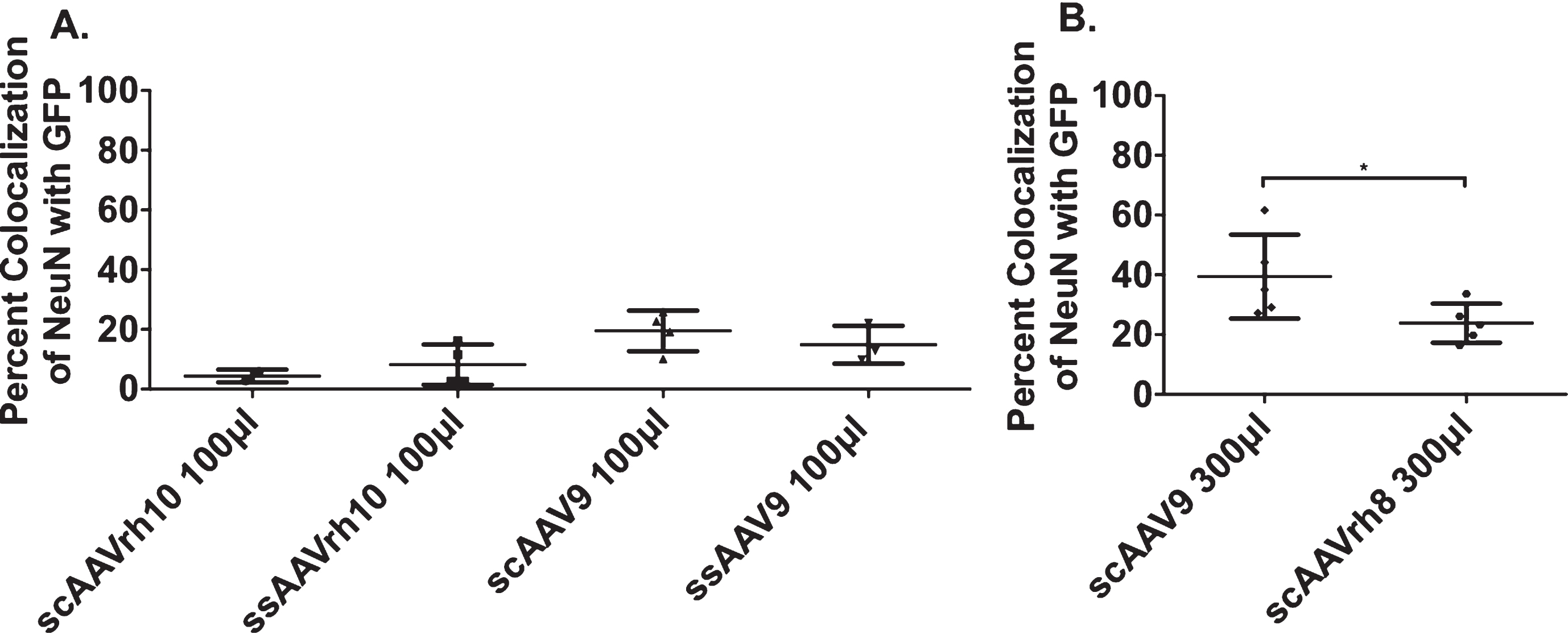
scAAV9 300μl transduces the highest percent of neurons. Sheep were injected with six different variants of AAV and were sacrificed 28 days post-injection. After brain slicing, serial sections were co-labeled with NeuN and GFP to measure the transduction efficiency of each AAV variant. Data are represented as percent co-localization (±SD) A) 100μl Injections. scAAVrh10 (n = 2) had 4.4% (±2.1) co-localization, ssAAVrh10 (n = 4) had 8.2% (±6.8), scAAV9 (n = 4) had 19.5% (±6.8), and ssAAV9 (n = 3) had 14.9% (±6.4) co-localization. B) 300μl Injections scAAV9 (n = 5) had 39.4% (±14.1) and scAAVrh8 (n = 5) had 23.8% (±6.6) co-localization. All error bars represent standard deviation. *p < 0.05; Mann-Whitney T-Test.
scAAV9 300μl spreads throughout the neostriatum
The distribution of AAV through the neostriatum, or how far the virus reaches from the site of injection, was quantified using stereology (described in Methods). Spread of a viral vector can be a separate property from neuronal uptake. We performed immunohistochemistry for GFP on every twentieth section, measured the distribution of GFP positive cells in the sections and compared GFP spread to that of DARPP32, a marker for striatal neurons. First, we compared ssAAVs and scAAVs of the same serotype. Interestingly, while we found ss and sc had no effect on neuronal transduction, we found that scAAVs improved AAV distribution in the neostriatum (Fig. 2A). scAAVrh10 offered the best spread in the striatum (100μl, 72%; Fig. 2A), although neuronal uptake was sparse. Given that scAAV9 300μl had the best neuronal uptake (Fig. 1), we assessed the distribution of scAAV9 300μl and scAAVrh8 300μl. We found that scAAV9 had the best combination of spread through the striatum and neuronal uptake: scAAV9 100μl (56%; Fig. 2A) and scAAV9 300μl (57%; Fig. 2B, C) (Table 1), although greater neuronal uptake was evident with the larger volume. scAAV9 300μl was detected in the ipsilateral thalamus and substantia nigra, areas that project to the striatum (Fig. 2D, E).
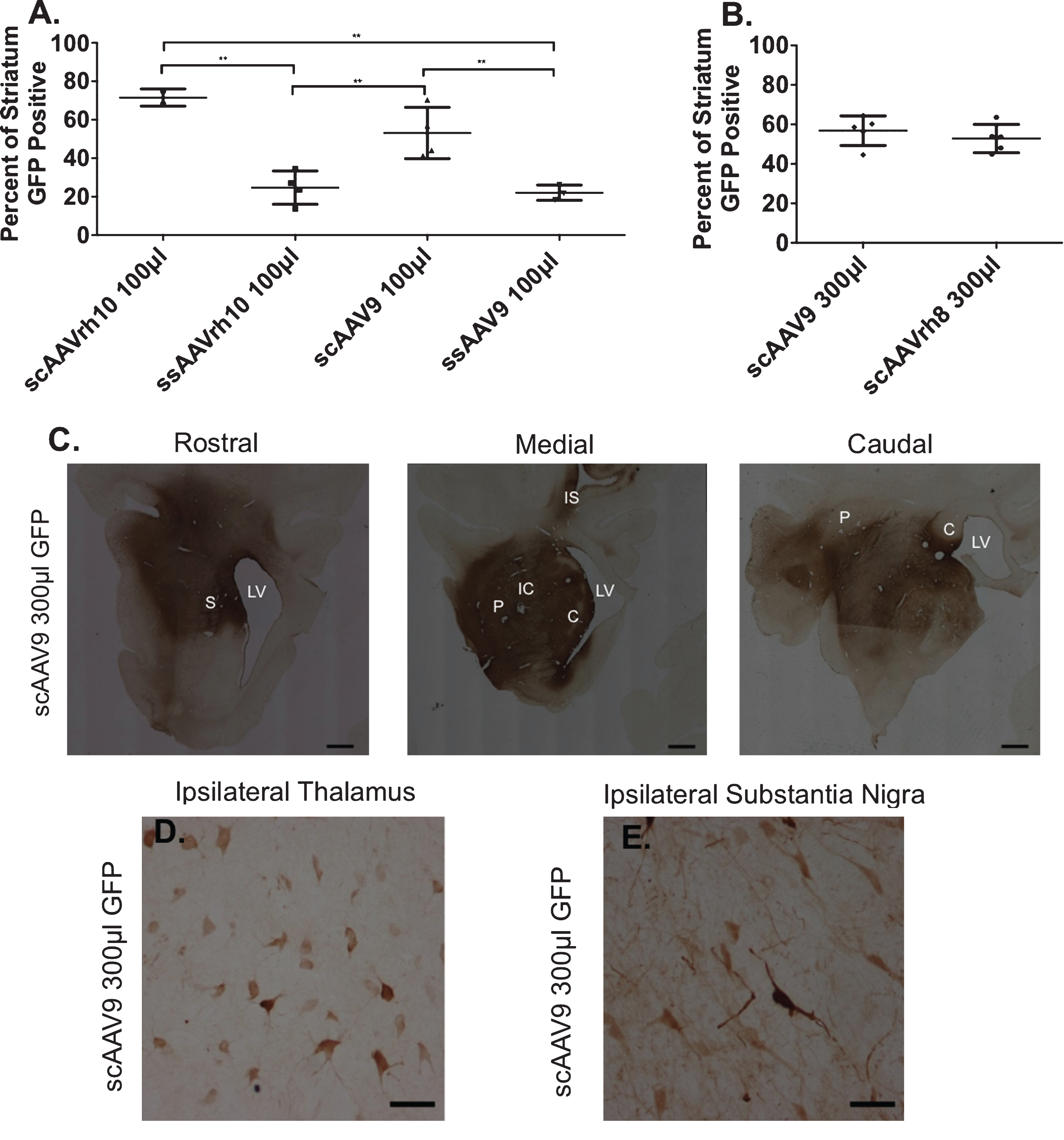
scAAV9 300μl spreads through the striatum. Serial sections were immunolabeled with anti-GFP antibodies to assess the distribution of AAV in the brain. Data are represented as percent of striatum that is GFP positive (±SD). A) 100μl Injections. scAAVrh10 spread 71.5% (±4.5), ssAAVrh10 spread 24.7% (±8.7), scAAV9 spread 53.1% (±13.3), and ssAAV9 spread 22.1% (±4.0). **p < 0.01; Ordinary one-way ANOVA Tukey’s multiple comparisons test. All error bars represent standard deviation B) 300μl Injections. scAAV9 spread 56.8% (±7.5) and scAAVrh8 spread 52.8% (±7.1). All error bars represent standard deviation. B-E.) Representative image of scAAV9 300μl anti-GFP immunohistochemistry shows transduced cells in the rostral, medial, and caudal striatum (scale bar = 30 cm) (C), ipsilateral thalamus (D), and ipsilateral substantia nigra (E) after intrastriatal injection. Scale bar = 50μm. S = Striatum, LV = Lateral Ventricle, P = Putamen, C = Caudate, IC = Internal Capsule, and IS = Injection Site.
Summary of Results
Brain administration of AAVrh8 and AAV9 leads to their widespread detection outside the brain
To find if AAVrh8 or AAV9 neostriatal injection entered non-brain structures after 300μl injection, we measured the number of scAAVrh8 and scAAV9 vector genome copies in blood, wool, feces, urine and saliva at baseline, 6 h, day 1, day 7, day 14, day 21 and day 28 post-injection (Fig. 3A-E). Except for wool, all samples had basal levels of AAV vector genome copies by two-week post-injection (measurement below 102 copies are below confidence limits). Secretions into the wool contained AAV copies at day 28. We also measured vector genome copies in the gonads, muscle, adrenal gland, heart, liver, cerebellum, cortex, and striatum of animals injected with either scAAV9 or scAAVrh8 at necropsy. The vector genome copies in all organs, other than the brain, fell below 102 28 days after injection (Fig. 3F, G).

Vector Genomes copies fell below positive cut off line for all samples tested except wool. To assess the distribution of AAV to different parts of the body after direct injection into the left striatum, we measured the number of AAV vector genome copies in blood, wool, feces, urine, and saliva at seven different time points for AAV9 300μl (n = 5) and AAVrh8μl (n = 8). We also measured the number of AAV vector genome copies in the gonad, muscle, adrenal gland, heart, liver, cerebellum, cortex, and striatum 28 days after injection, for AAV9 300μl (n = 3) and AAVrh8μl (n = 3). R = Contralateral side, L = Ipsilateral side. We selected a line of detection to be 102 AAV vector genome copies/μg gDNA. A) Vector genomes in the blood fell below 102 by day 7 for AAVrh8 and day 28 for AAV9. B) Vector genomes in the feces fell below 102 by day 14 for AAVrh8 and day 7 for AAV9. C) Vector genomes in the urine fell below 102 by day 7 for AAVrh8 and day 14 for AAV9. D) Vector genomes in the saliva fell below 102 by day 21 for AAVrh8 and day 14 for AAV9. E) Vector genomes in the wool did not fall below 102 for both AAVrh8 and AAV9. F-G) AAV9 (F) and AAVrh8 (G) vector genome copies in the gonad, muscle, adrenal gland, heart, and liver fell below 102. As expected, AAV9 and AAVrh8 genome copies were above the positive cut off line in the cerebellum, cortex, and striatum on both the ipsilateral and contralateral sides.
No significant loss of DARPP32 containing neurons after scAAV9 administration to the neostriatum
To evaluate safety, we measured the total number of DARPP32 labeled cells per mm3 of tissue, the mean area of DARPP32 labeled cells, and the volume of the striatum. The contralateral (non-injected) brain served as a control. No differences in the total number of DARPP32/mm3 cells in the ipsilateral striatum, mean area of DARPP32 labeled cells, or in the overall volume of the striatum were detected when animals were injected with 300μl of scAAV9 (Fig. 4A-C), or any of the other AAV variants tested (Supplementary Figure 3). Furthermore, alertness, blood pressure, respiratory rate, heart rate, weight change, and eating frequency were in the normal range. Post-operative recovery was uneventful.
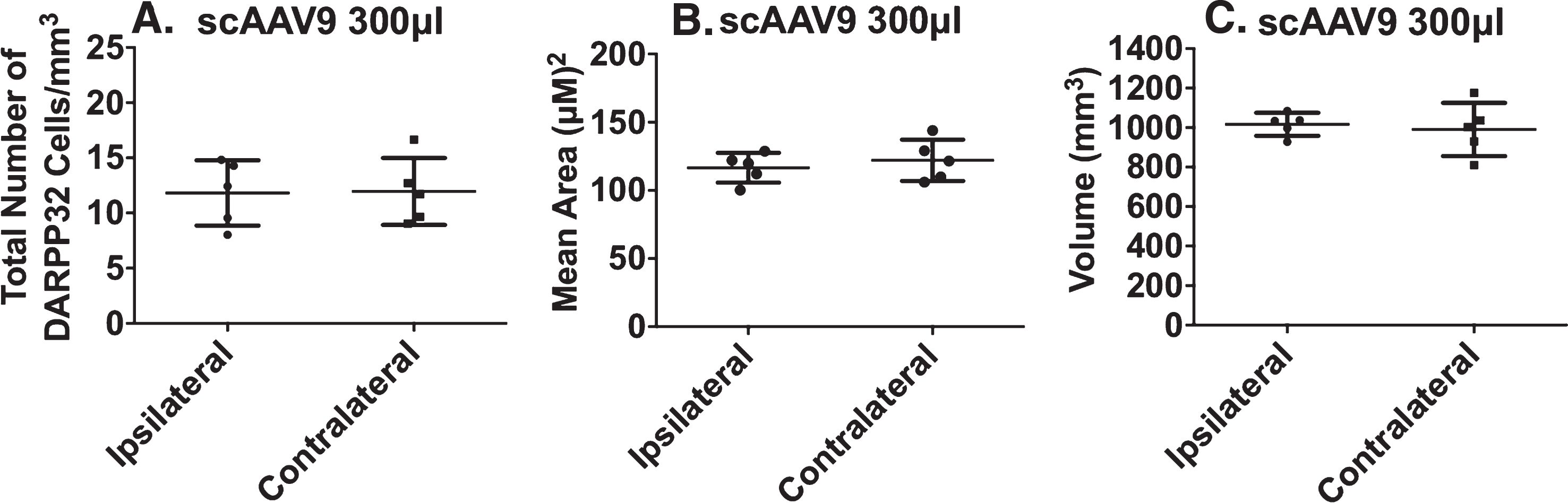
No DARPP32 cell toxicity after injection with scAAV9 300μl. Serial sections were immunolabeled with DARPP32 to assess the safety of scAAV9 300μl in the brain (n = 5). A) Stereological methods were used to count the total number of DARPP32 cells, in a maximum of ten different locations per tissue section, throughout the striatum. The total number was then normalized to the volume of the striatum to determine the total number of DARPR32 cells/mm3. There is no significant difference in the number of DARPP32 cells/mm3 on the ipsilateral compared to the contralateral side of the brain (unpaired student’s t-test; p = 0.9451). B) The cross sectional area of DARPP32 cells were measured, one cell per location in a maximum of ten different locations per tissue section, throughout the striatum. The areas were then averaged together to determine the mean area (μM)2. There is no significant difference in the mean area of cells on the ipsilateral compared to the contralateral side (unpaired student’s t-test; p = 0.5306). C) The volume of the striatum was calculated by measuring the total area of DARPP32 per section and multiplying by the distance between each section and the thickness of each section. There is no significant difference in the striatal volume on the ipsilateral compared to the contralateral side (unpaired student’s t-test; p = 0.7096).
DISCUSSION
Our findings demonstrated that AAV neuronal uptake can be species specific. We considered three AAV serotypes: AAVrh8, AAV9 and AAVrh10. Our initial selection was AAVrh10, because AAVrh10 infects mouse neurons [6] and has been directly administered to patients [12, 13]. To our surprise, in the sheep striatum, AAVrh10 did not transduce neurons. Compared to AAVrh10 and AAVrh8, AAV9 had the best combination of uptake and spread at one month post-administration. In a prior study in HD transgenic sheep, a long-lasting microglia response was absent [9]. These findings contrasted with the report that an AAV9 and a capsid variant, PHPB, manifested toxicity in non-human primates and pigs [14, 15]. Dose differences might explain the divergent results. The titer in our study was 1×10 E13, whereas the dose in the NHP study was 1×10 E14. Unaltered, AAV9 can crosses the blood-brain barrier although to a limited extent [16]. Modification by alanine on its capsid [17], or other capsid modifications such as AAV-PHP.B [18], could facilitate AAV9 crossing the blood-brain barrier. This property in AAV-PHP.B might be species specific [14]. Under certain conditions, administration to the cerebral spinal fluid, especially through the cisterna magnum, leads to widespread neuronal and glial uptake of the virus in the brain and spinal cord [19]. Properties of AAV serotypes might vary among species, an important implication for therapeutic applications.
scAAV is thought to have rapid production and higher gene delivery efficiency because of the immediate availability of double stranded RNA [20]. The space available in scAAV is smaller than ssAAV, but can accommodate artificial microRNAs. Use of scAAV-miRNA against human mutant huntingtin successfully reduced expression of human mutant huntingtin in transgenic Huntington disease sheep, without recruiting an innate immune response [9]. For the purpose of this study, rapid expression of the AAV was advantageous for demonstration of neuronal uptake and spread in the brain. But, rapid expression might not be needed for a treatment that should last in HD patients for many years. ssAAV is easier to produce than scAAV and therefore is more accessible. Ultimately, both types of AAV would serve to deliver miRNA to neurons, to lower huntingtin.
AAV9 showed good safety criteria. AAV9 did not overtly perturb morphology of medium spiny neurons in the striatum, as evaluated by counts and morphology of DARPP32 neurons. At one month, AAV9 injected sheep did not manifest obvious behavioral changes. The dimensions of the striatum are unchanged by injection of AAV9, indicating that various cell types are largely unchanged. Vital signs of the sheep remained normal through the duration of the study.
Convection enhanced, intra-parenchymal brain administration of AAV in sheep did not limit spread of the virus to the brain. By 6 h after delivery, AAVrh8 and AAV9 entered the circulation and soon thereafter, was measurable in feces and filtered or secreted by the kidney to enter urine. However, by two weeks, AAVrh8 and AAV9 was no longer detectable in blood, feces, urine or saliva. The exception was secretion into wool, in which a low but steady state of virus was present at four weeks. Sheep might lick other sheep; in theory, the wool could be a portal for viral sharing. Nonetheless, the implication was that AAV shedding of this non-replicative virus was temporary. This finding was not new for administration of AAV to brain. AAV2 injected into brain was detected in spleen six months after injection and was cleared by liver, heart and kidney [21]. By virtue of its detection in blood, AAV9 had access to the body whole. Thus, brain administration of AAV9 can reach peripheral organs that expressed mutant huntingtin, thereby providing access to other sites of potential disease.
CONFLICT OF INTEREST
Guangping Gao is a co-founder of Voyager Therapeutics and holds equity in the company. Guangping Gao is inventor on patents with potential royalties licensed to Voyager Therapeutics and other biopharmaceutical companies.