
Abstract
Background:
Huntingtin (htt) protein is an essential regulator of nervous system function through its various neuroprotective and pro-survival functions, and loss of wild-type htt function is implicated in the etiology of Huntington’s disease. While its pathological role is typically understood as a toxic gain-of-function, some neuronal phenotypes also result from htt loss. Therefore, it is important to understand possible roles for htt in other physiological circumstances.
Objective:
To elucidate the role of htt in the context of ethanol exposure, we investigated how loss of htt impacts behavioral and physiological responses to ethanol in Drosophila.
Methods:
We tested flies lacking htt for ethanol sensitivity and tolerance, preference for ethanol using capillary feeder assays, and recovery of mobility after intoxication. Levels of dopamine neurotransmitter and numbers of dopaminergic cells in brains lacking dhtt were also measured.
Results:
We found that dhtt-null flies are both less sensitive and more tolerant to ethanol exposure in adulthood. Moreover, flies lacking dhtt are more averse to alcohol than controls, and they recover mobility faster following acute ethanol intoxication. We showed that dhtt mediates these effects at least in part through the dopaminergic system, as dhtt is required to maintain normal levels of dopamine in the brain and normal numbers of dopaminergic cells in the adult protocerebrum.
Conclusions:
Our results demonstrate that htt regulates the physiological response to ethanol and indicate a novel neuroprotective role for htt in the dopaminergic system, raising the possibility that it may be involved more generally in the response to toxic stimuli.
INTRODUCTION
Huntingtin (htt) is a multifunctional protein essential for regulating embryogenesis and development of the central nervous system, as well as adult neuronal health. Htt is most commonly studied for its role in the pathogenesis of Huntington’s disease (HD), a genetic disorder characterized by the progressive degeneration of medium spiny neurons in the striatum. HD’s pathology is predominately viewed as a toxic gain-of-function due to expression of a mutant form of htt. Some HD phenotypes, however, also result from loss of normal htt function [1]. Consistent levels of htt expression are required for proper regulation of cellular processes such as axonal transport, vesicle transport, and organelle morphology and distribution [2].
Wild-type htt also has significant protective and pro-survival functions in neurons. Overexpression of wild-type htt in the rodent brain can reduce the toxicity of mutant expanded htt [3] and also protect neurons from apoptotic stimuli in vitro [4]. Htt also plays a role in enabling neurons to withstand stressful events, such as serum deprivation, mitochondrial toxins, or the transfection of death genes in vitro [4–6] and in vivo [3, 7], and can provide a dose-dependent neuroprotection from excitotoxicity in mice [8]. Moreover, htt expression enhances neurotrophic support for corticostriatal neurons by protecting against excitotoxicity and reducing neurodegenerative apoptosis [3]. These effects likely arise from its role as a transcriptional regulator and modulator of its protein binding partners. In particular, htt upregulates brain-derived neurotrophic factor (BDNF) transcription [1], while also inhibiting the activity of caspases [4]. Though htt has been shown to be an important neuroprotective molecule in multiple situations, little is known about how htt may function during or after ethanol exposure.
Htt normally has many interacting partners [9], so not surprisingly, loss of wild-type htt function has been implicated in other neurologic disease models. Wild-type htt levels are depleted in mouse models of HD, ischemic brain injury, and traumatic brain injury [7]. Evidence also suggests a key role for htt in modulating the developmental cellular response to ethanol. Acute developmental ethanol exposure results in a myriad of gene expression changes in the adult mouse, where alterations in glutamate receptors, neurotrophic molecules, and htt appear as core “hub” genes within the dysregulation, regardless of the timing of ethanol exposure [10]. Htt connects a network of genes altered by exposure to acute high developmental ethanol doses in the first (embryonic day E8/11), second (E14/16), or third trimester (postnatal day P4/7) equivalents in mice. The relationship between htt and alcohol has been previously undescribed. However, gene expression analysis in Fetal Alcohol Spectrum Disorder (FASD) models shows not only multiple adult gene alterations centered around htt, but that htt gene expression itself is also downregulated in adult mice (at postnatal day 60) after acute ethanol exposure at P4/P7 (–1.23 fold-change) [10].
A recent study using the Enroll-HD global platform pinpointed an interaction between alcohol and the clinical presentation of HD, finding that past alcohol misuse significantly increases the clinical symptoms of HD (including motor impairment and higher psychiatric symptoms) but does not appear to impact age of symptom onset [11]. In addition, some studies suggest that alcohol dependence is more common in the HD population [12–14].
Htt’s already well-established role in neuropathology, combined with clear evidence of htt involvement in persistent gene alterations after acute [10] and chronic [15] ethanol exposure, and the fact that alcohol misuse can modulate the clinical presentation of patients with HD [11] make htt’s response to ethanol a logical avenue for exploration. To date, the idea that htt may regulate of ethanol responses is based only on gene array analyses, but our study explores this role in vivo by examining behavioral and neurochemical responses to ethanol in the absence of the htt protein in Drosophila melanogaster.
Drosophila is a well-characterized model for HD pathogenesis. A homolog of mammalian htt has been characterized in Drosophila (hereafter referred to as dhtt) and shown to share conserved functional domains with the human gene [16]. Introduction of expanded polyglutamine repeats has replicated the dominant gain-of-function aspect of HD in flies by disrupting the intricate organization of the many neuronal photoreceptors the fruit fly compound eye, as well as impacting motor behavior in a degenerative manner [17–19]. The Drosophila model has also replicated loss-of function aspects of the disorder [20]. While a complete lack of htt during development is lethal in mice [21], dhtt-null flies are viable throughout development and have also been a useful model for HD [22].
In addition to HD, previous Drosophila models have also been used to investigate effects of dopamine deficiency [23], Parkinson’s disease using both gain-of-function [24] and loss-of-function [25] models, FASD [26], and alcohol reward pathways [27]. In order to clarify the role of wild-type htt in the context of ethanol exposure, we investigated how loss of htt impacts behavioral and physiological responses to ethanol in Drosophila, including sensitivity, tolerance, recovery from intoxication, and preference.
MATERIALS AND METHODS
Fly lines and husbandry
The dhtt-KO mutant fly line, which harbors a deletion of 27 exons from the dhtt gene, was previously described [22]. For all experiments, w1118 (BDSC stock number 3605) was used as a control since the dhtt-KO line was generated in this genetic background. w1118 and dhtt-KO flies were maintained at room temperature in standard vials and fed a nutritionally complete diet containing yeast, sucrose, cornmeal, and agar. PCR confirmed the deletion of the dhtt gene from dhtt-KO flies. Genomic DNA was extracted from single flies according to standard procedures, and genotyping PCR was performed using Taq 2X Master Mix (New England Biosciences) and three separate primer pairs: N forward: 5′-GCCAATGTAGCCAGAGTCTG-3′, N reverse: 5′-CGCATTCGCTGATGCTGCGTG-3′ (873 bp product); M forward: 5′-AAGCTATTCGAGCCGATGGTC-3′, M reverse: 5′-GCACCAGGAATCTCAGCATGG-3′ (721 bp product); C forward: 5′-TCGGGAATTGACTTTCGCAGC-3′, C reverse: 5′-TGCAGTTTGAGGCAGCGTTCC-3′ (405 bp product).
Ethanol sensitivity and rapid tolerance
Zhang et al. [22] reported a striking age-related decline in the mobility of dhtt-KO flies that was evident by 25 days of age. Therefore, we tested a role for huntingtin in mediating ethanol sedation sensitivity and rapid tolerance in 28-day-old flies (28 days after eclosion) [28]. Two groups of flies were assayed (each containing 45 dhtt-KO and w1118 individuals): one group previously exposed to ethanol and another with no prior exposure. For the flies exposed to ethanol before sensitivity and tolerance were measured, 10-day-old flies of both genotypes were placed on standard food supplemented with 5% ethanol for 24 hours. Following this ethanol exposure, flies were transferred to fresh vials containing standard food until the sensitivity assay was performed when the files were 28 days old.
To test ethanol sensitivity, 28-day-old flies were sedated using CO2 and separated into empty vials (10–15 individuals each). After 10 minutes of recovery, 0.5 ml of 100% ethanol was pipetted onto a fresh cotton plug and placed on each vial. The time it took for individual flies to become immobilized was measured. At one-minute intervals after the addition of ethanol, vials were tapped down three times on the tabletop and the number of stationary flies counted during a 10-second observation period. A fly was deemed stationary if it could not get off its back, remained in one location, or remained stationary and rapidly vibrated its wings throughout the entire 10-second observation period [28].
To test rapid tolerance in these flies, they were allowed to recover mobility and motor function for four hours in a fresh vial containing food before being assayed again for ethanol sensitivity as described. A two-way analysis of variance (ANOVA) with Tukey’s HSD post hoc analysis was performed to examine the effects of genotype and prior adult ethanol exposure on sensitivity and rapid tolerance.
For all box plots, the boundary of the box closest to zero indicates the 25th percentile, the bold line within the box marks the median, and the boundary of the box farthest from zero indicates the 75th percentile. Whiskers above and below the box indicate the 90th and 10th percentiles, respectively.
Alcohol preference assay
Alcohol preference was tested using a capillary feeder (CAFE) assay in which flies are presented with two food sources, one containing ethanol, the other not [29]. The CAFE apparatus consisted of a standard plastic fly vial with a cotton plug through which two calibrated 50μl glass capillaries (Drummond Scientific, cat no. 2000050) were inserted vertically. The capillaries were filled either with liquid food solution (5% sucrose/5% yeast dissolved in water) alone or food solution supplemented with 15% ethanol. Red food color was added to visualize food consumption in flies, and a small mineral oil overlay was added to capillary ends to prevent evaporation. A damp Kimwipe was placed in the bottom of the vial to maintain humidity. Prior to the start of each assay, images of the starting food levels were acquired.
Alcohol preference of 28-day-old w1118 and dhtt-KO individuals was measured. Twenty individuals of the same genotype were placed into a CAFE apparatus and allowed to feed for 16 hours at room temperature. At the conclusion of the experiment, food levels in the capillaries were again imaged. Under these experimental conditions, we never observed an empty capillary at the conclusion of the assay, indicating that the amount of and access to food solutions were not limiting.
The volumes of ethanol and non-ethanol food consumed were calculated based on pixel measurements of the images. Data is presented as the average of measurements for each genotype±S.E.M. (n = 9 vials w1118; n = 8 vials dhtt-KO). To normalize for slight differences in overall food consumption, an ethanol preference index (PI), defined as [(ethanol food consumption) –(non-ethanol food consumption) / (total food consumption)], was calculated. Positive PI values indicate a preference for ethanol and negative values indicate aversion to ethanol. Differences in the average ethanol PI of w1118 and dhtt-KO flies were compared using a Student’s t-test.
Negative geotaxis assay
A negative geotaxis assay was carried out to determine the effects of age (10-day versus 28-day-old) and prior ethanol intoxication on mobility of dhtt-KO flies [30] Geotaxis was measured in triplicate for 30 individuals for each experimental treatment (age, genotype, and with or without prior ethanol exposure). For exposure to ethanol before the geotaxis assay, flies were transferred to empty vials with cotton plugs containing 0.5 ml 100% ethanol. The flies were exposed until all flies in the vial were immobilized. They were then transferred into a fresh vial containing food and allowed to recover for four hours before being assayed for geotaxis.
To conduct the assay, a group of 10 individuals was placed in an empty standard vial. The flies were tapped down three times and given 10 seconds to demonstrate startle-induced negative geotaxis by crawling toward the top of the vial [31]. After 10 seconds, the vial was imaged and the distances of individual flies from the bottom of the vial was recorded. This procedure was repeated at one-minute intervals to yield three sets of data per experimental vial, and the average distance traveled for the three trials was calculated. Three separate vials (a total of 30 individuals) were measured for each experimental treatment. A three-way ANOVA with Tukey’s HSD post hoc analysis was used to test for the effects of age, genotype, and prior alcohol exposure on mobility.
Alcohol metabolism assay
Alcohol metabolism over time was measured using a colorimetric assay to detect absolute alcohol levels in fly homogenates [32]. Thirty flies per treatment group were placed into empty plastic vials and allowed to equilibrate. Vial plugs were then replaced with plugs to which 1 ml of 80% ethanol had been added. Flies were exposed to evaporating ethanol for 5 minutes. The flies were then transferred to fresh empty vials and the amount of alcohol in 10 individuals (five of each sex) was measured after 0, 30, and 60 minutes. Each group was homogenized in 200μl of 50 nM Tris-HCl (pH 7.5), centrifuged at 15,000 g to clarify, and 5μl of supernatant was prepared for a colorimetric assay using the Sigma-Aldrich Ethanol Assay Kit (Sigma-Aldrich; catalog no. MAK076) according to manufacturer’s instructions. Absorption was measured at 570 nm using a Synergy HT plate reader (BioTek) alongside known standards. Measurements from three homogenates were taken for each treatment group and averaged together.
Detection of neurotransmitters
Quantification of tyramine, serotonin, octopamine, and dopamine was performed via capillary electrophoresis with fast-scan cyclic voltammetry (CE-FSCV) as previously described [33]. Briefly, single brains were removed from 25-day-old flies of both genotypes, diluted with a perchloric acid-acetonitrile matrix, homogenized, and centrifuged. Electrokinetic injections were made by placing the end of the separation capillary directly into sample vials. Detection was performed with fast-scan cyclic voltammetry with a two-electrode configuration using a Dagan ChemClamp potentiostat. The electrode was scanned from –0.4 to 1.3 V and back at 400 V/s every 100 ms. Hardware and software were configured for data acquisition as described previously [34]. Estimates of neurotransmitter concentrations in fly samples were calculated from standard samples run before and after each experimental sample. Measurements were acquired from five brains each of control and dhtt-KO flies. Each analysis detected neurotransmitter in five pooled brains (divided by five to obtain neurotransmitter pg/brain; methods outlined in [35]). T-tests were used to compare the mean concentration of each neurotransmitter between genotypes.
Immunohistochemistry and imaging
Brains from 25-day-old flies were dissected in PBS and fixed in 4% paraformaldehyde/PBS for 20 minutes. After thorough washing in 0.1% Triton-x/PBS, antibody staining was performed according to standard protocols. Primary antibodies used in this study were: mouse anti-discs large (1 : 200; Developmental Studies Hybridoma Bank) and rabbit anti-tyrosine hydroxylase (1 : 100; Chemicon). Secondary antibodies were conjugated to Alexa Fluor dyes (1 : 1000; Thermo Fisher).
Z-stacks encompassing entire individual central brain lobes were collected using a Leica SP8 laser scanning confocal microscope. Central brain regions were identified based on discs large staining. The number of tyrosine hydroxylase-positive neuron cell bodies in each region were counted in ImageJ. Data were tested for significance using a Student’s t-test.
RESULTS
To characterize a possible role for huntingtin in mediating alcohol toxicity, we tested ethanol sensitivity and tolerance using a Drosophila line harboring a deletion of the huntingtin gene (hereafter referred to as dhtt-KO) [22]. Flies (n = 45 individuals for each treatment) were fed a normal diet or one supplemented with 5% ethanol for 24 hours at 10 days of age and subsequently tested for ethanol sensitivity at 28 days of age. Sensitivity was measured by the immobilization time of individual flies after acute exposure to vaporized ethanol (Fig. 1A, B). Previous ethanol exposure significantly increased sensitivity to ethanol in both dhtt-KO and w1118 flies (two-way ANOVA, p < 0.001) (Fig. 1D). Flies lacking dhtt were markedly less sensitive to ethanol compared to w1118 flies regardless of previous ethanol exposure (two-way ANOVA, p < 0.001) (Fig. 1C). To investigate whether dhtt is involved in the development of shorter term tolerance, we retested these flies after 4 hours of recovery (Fig. 1E, F). Consistent with measures of sensitivity, previous ethanol exposure at 10 days of age increased tolerance in both genotypes (two-way ANOVA, p < 0.001) (Fig. 1 H), and dhtt-KO flies showed increased tolerance compared to w1118 (two-way ANOVA, p = 0.01) (Fig. 1G).
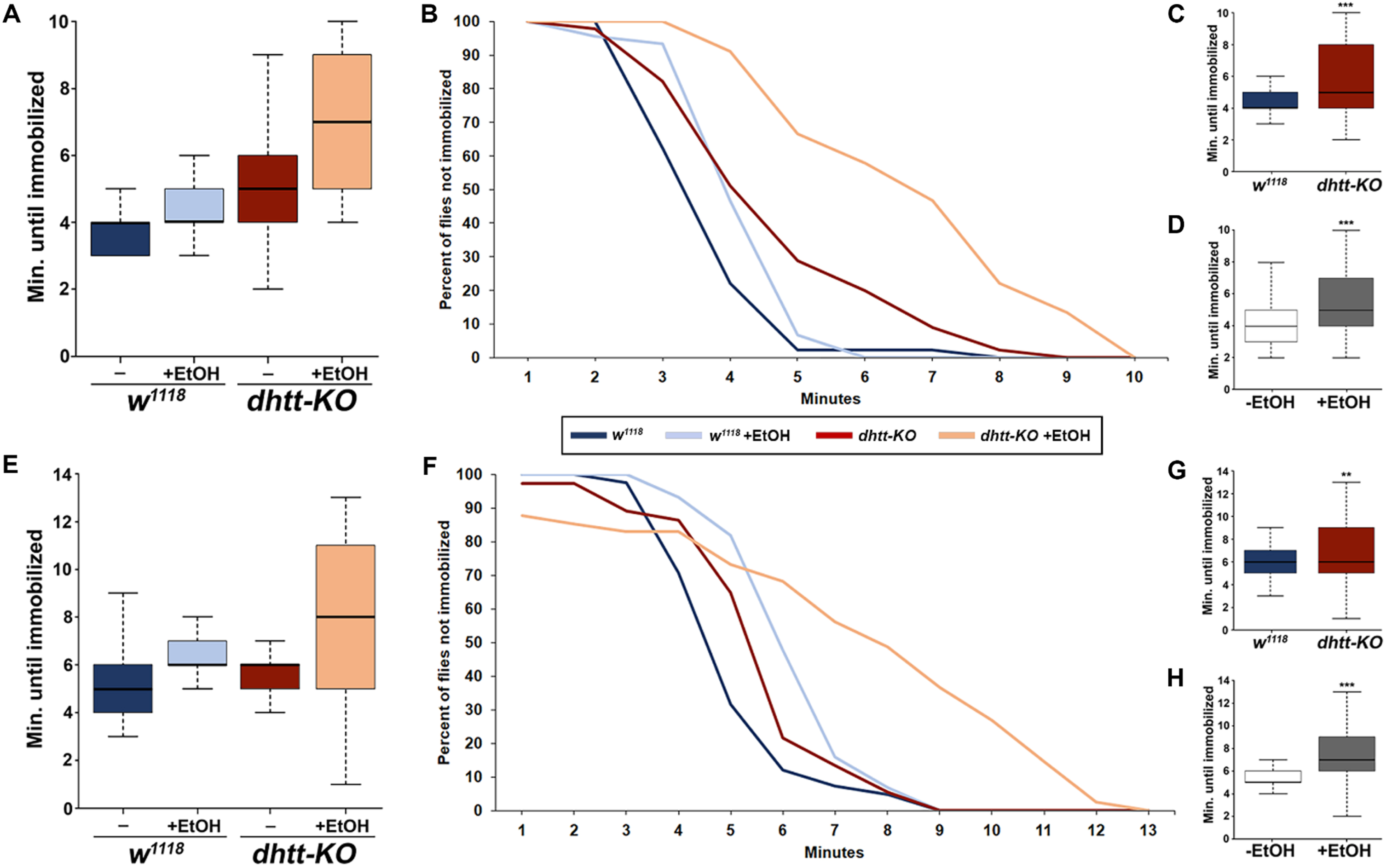
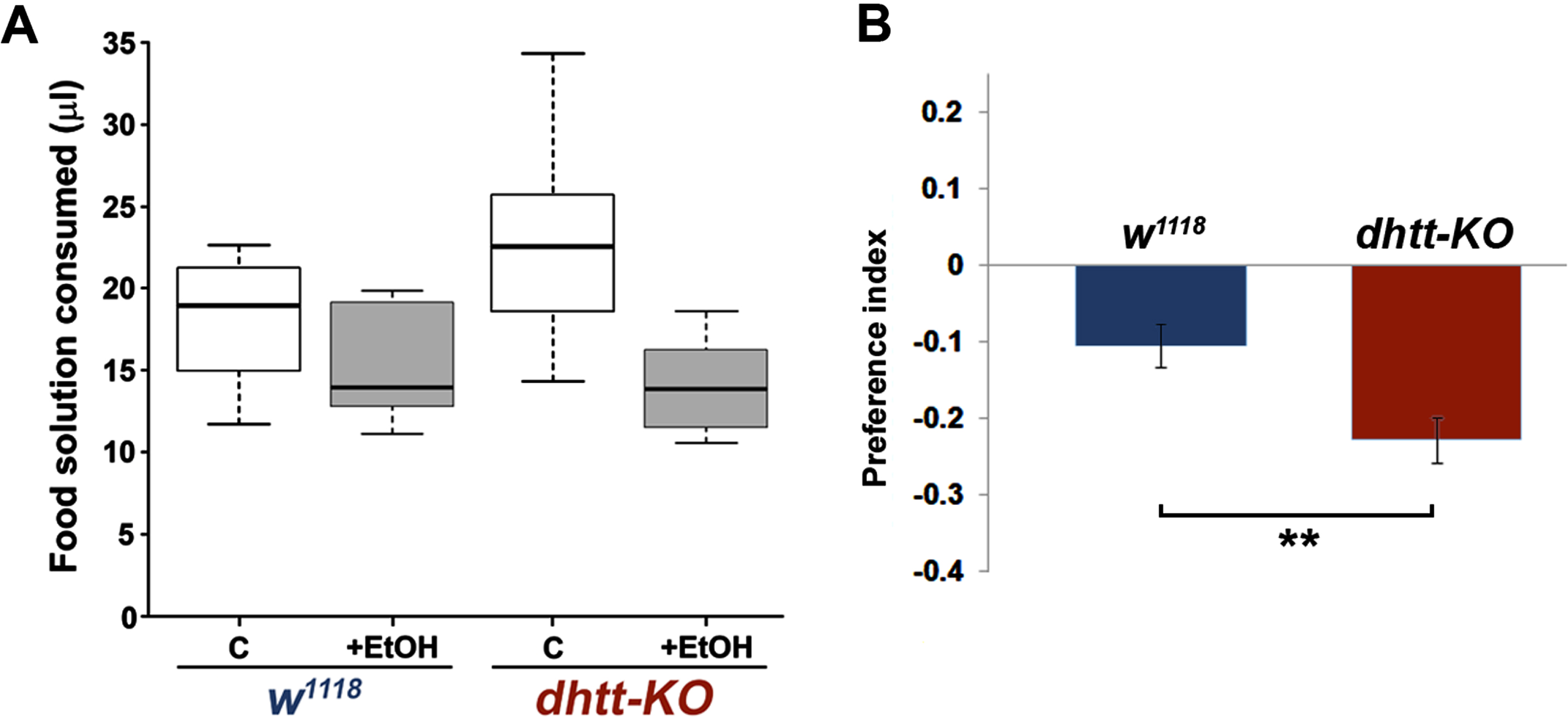
We next examined alcohol preference in 28 day-old dhtt-KO flies using a capillary feeder (CAFE) assay [29]. Twenty flies per vial were presented with two capillaries, containing a food solution (5% sucrose/5% yeast extract) or the same food solution supplemented with 15% ethanol, and allowed to feed for 16 hours (Fig. 2A). After measuring the total amount of each solution consumed by each genotype (n = 9 vials w1118; n = 8 vials dhtt-KO), ethanol preference was quantified by calculating a preference index defined as (ethanol food consumption –non-ethanol food consumption) / total food consumption. Flies lacking dhtt exhibited a decreased preference for ethanol-containing food compared to their w1118 counterparts (Student’s t-test, p = 0.007) (Fig. 2B).
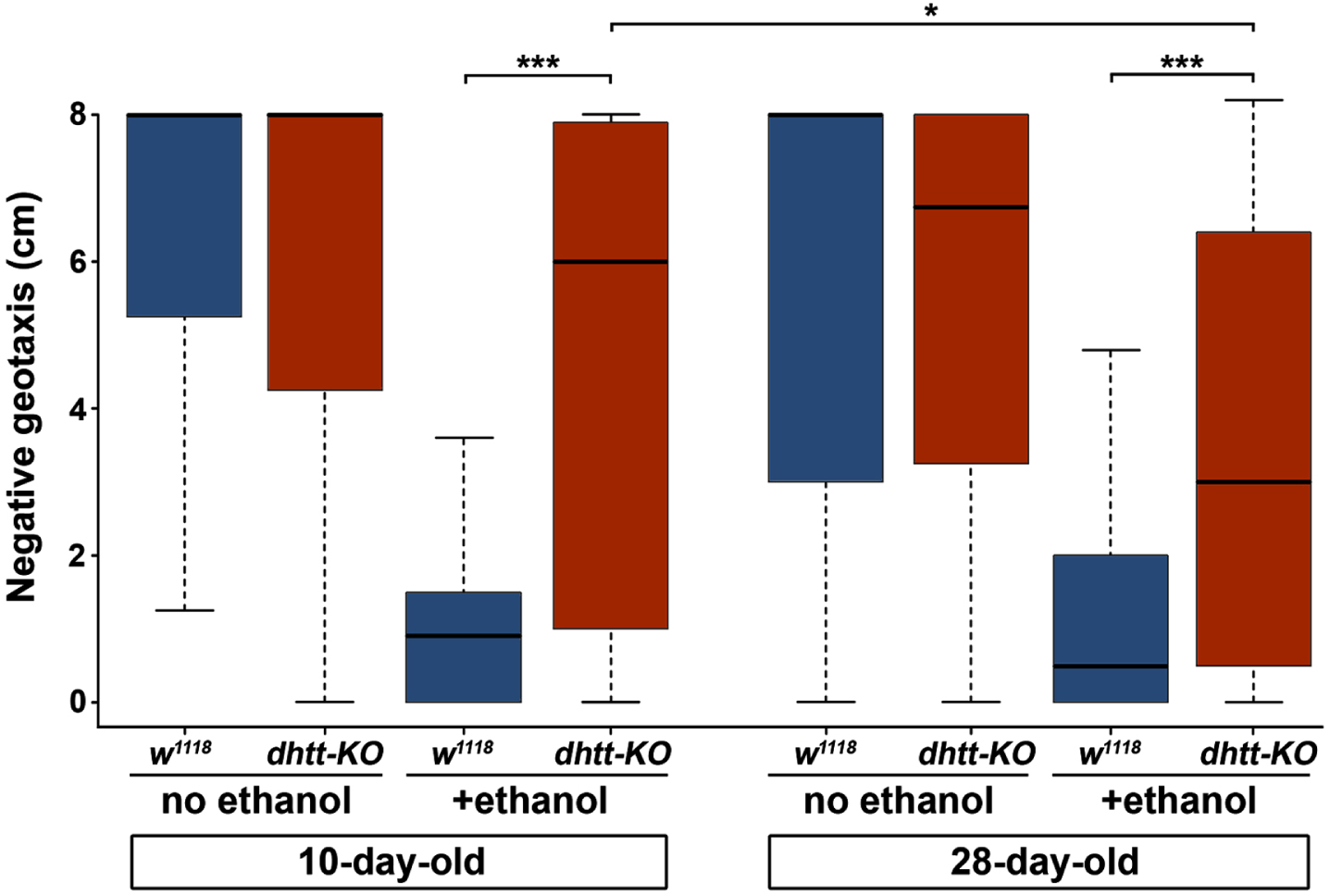
A progressive neurodegenerative phenotype that affects mobility in aged dhtt-KO flies has been described previously [22]. Therefore, we next assessed the effects of age, genotype, and prior ethanol intoxication on the mobility of flies using a negative geotaxis assay (Fig. 3). In these experiments, flies were tapped down to the bottom of empty vials and the location of individual flies along the vertical axis of the vial was measured after 10 seconds (n = 30 for each genotype and age). The assay was repeated on each vial for a total of three trials, and the data averaged. To examine the effect of prior ethanol intoxication, flies of both ages and genotypes were completely immobilized by exposure to vaporized ethanol and allowed to recover for four hours before the assay. A three-way ANOVA with Tukey’s HSD post hoc analysis was used to assess the effects of age, ethanol, and genotype on negative geotaxis scores. As expected, ethanol intoxication significantly reduced negative geotaxis scores (three-way ANOVA, p < 0.001). Of note, there was no significant interaction between age alone or age and genotype in combination (three-way ANOVA, p = 0.93 and p = 0.43, respectively). However, we did observe a highly significant interaction between genotype and the ability to recover mobility after ethanol intoxication: both 10-day-old (4.63 cm±0.24) and 28-day-old dhtt-KO (3.49 cm±0.26) flies scored higher in the negative geotaxis assay compared to 10-day-old (1.23 cm±0.11) and 28-day-old (1.05 cm±0.08) w1118 flies (three-way ANOVA, p < 0.001) (Fig. 3). Moreover, recovery of mobility decreased in 28-day-old dhtt-KO flies after intoxication, supporting an age-related decline in the ability of htt to mediate alcohol toxicity (three-way ANOVA, p = 0.029) (Fig. 3). Taken together, our results indicate that dhtt has a role in regulating physiological and behavioral responses to ethanol exposure.
The differences we observed in dhtt-KO flies could be due to an altered ability to metabolize ethanol in these individuals. To test this possibility, we exposed 10- and 28-day-old w1118 and dhtt-KO flies to ethanol vapor and measured the amount of alcohol present in pooled homogenates from 10 individuals after 0, 30, and 60 minutes of recovery. Data from three independent trials for each age and genotype were averaged together. We detected partial alcohol clearance in both 10- and 28-day-old flies after 1 hour (Fig. 4). Consistent with previous reports of age-related changes in alcohol metabolism [36, 37], older flies were less efficient at clearing alcohol after intoxication. While w1118 and dhtt-KO flies differed in the amount of ethanol metabolized after 30 minutes at both ages, after 60 minutes of recovery, the total ethanol remaining was essentially equal between genotypes in both 10-day-old (6.38 nmol±0.69 versus 6.31 nmol±0.18; p = 0.39, Student’s t-test) and 28-day-old (7.42 nmol±0.21 versus 7.08 nmol±1.16; p = 0.78, Student’s t-test) flies (Fig. 4A, C). Likewise, after 60 minutes of recovery, the percent of total ethanol metabolized was indistinguishable between genotypes at both ages (Fig. 4B, D). These data support the idea that dhtt-KO flies have normal metabolic function, and that our physiological and behavioral observations were likely due to a more direct role for dhtt.

The changes in ethanol preference and tolerance we observed in dhtt-KO flies led us to investigate how loss of dhtt affects reward pathways involved in the ethanol response [27]. We used capillary electrophoresis-fast scan cyclic voltammetry (CE-FSCV), a technique that detects molecules based on their electrochemical signature [33], to quantify neurotransmitter amounts in whole brain lysates from dhtt-KO and w1118 flies. We measured the levels of four neurotransmitters: octopamine, tyramine, dopamine (DA), and serotonin (5-HT) in 25-day-old brains (Fig. 5A–D). Of these, only DA was significantly reduced in dhtt-KO brains (10.1 pg/brain±1.7, n = 5) compared to w1118 controls (22.3 pg/brain±5.0, n = 5) brains (Student’s t-test, p = 0.049) (Fig. 5B).
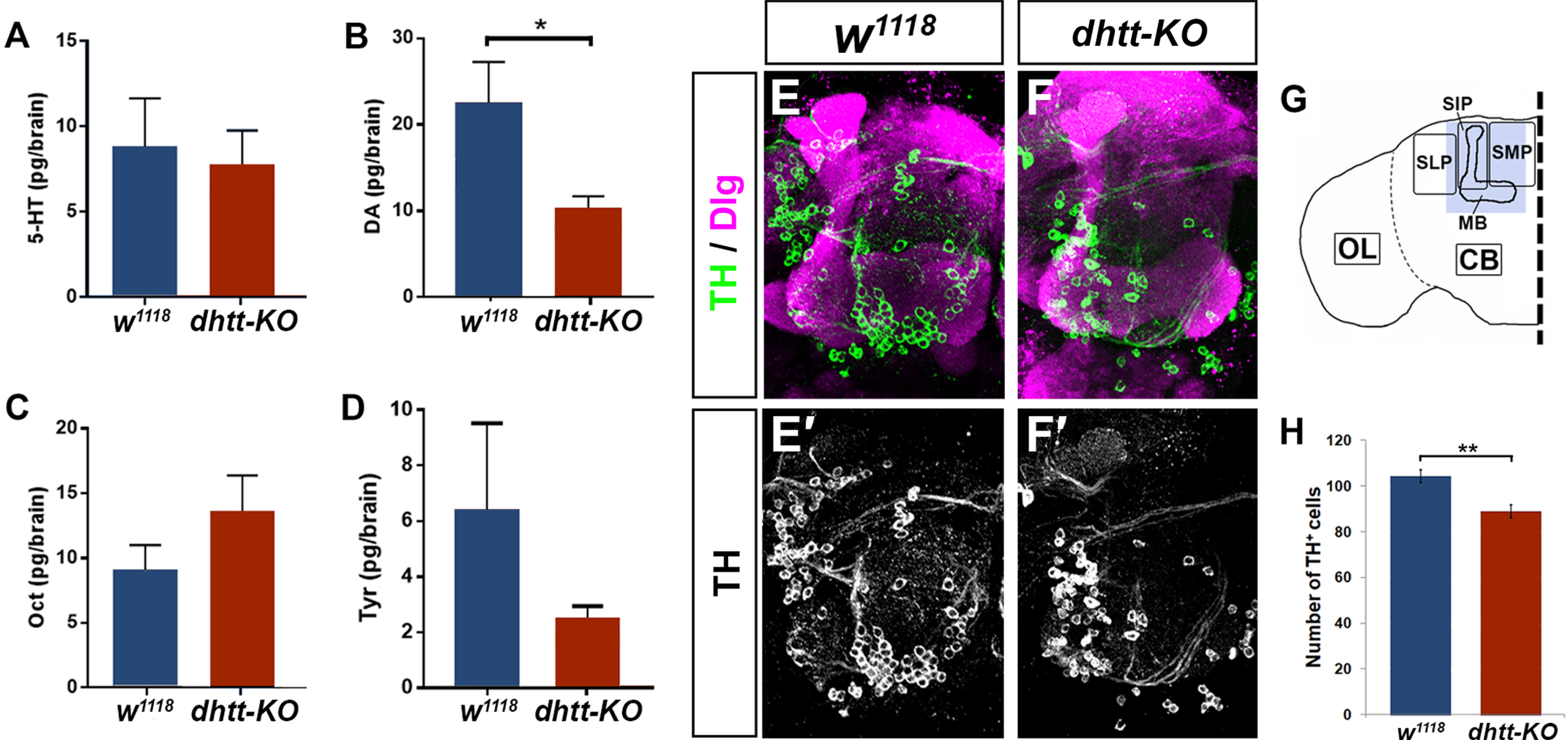
A possible explanation for the observed reduction in DA in dhtt-KO brains is a change in the population of dopaminergic cells. To test this hypothesis, we used tyrosine hydroxylase (TH) immunostaining to mark dopaminergic cells in 25-day-old brains (Fig. 5E, F). We observed reduced TH staining in the protocerebra of dhtt-KO brains compared to w1118 control brains. These changes were most pronounced in the medial, intermediate, and lateral superior protocerebrum which contains the protocerebral anterior medial (PAM) and protocerebral anterior lateral (PAL) clusters of dopaminergic neurons that project to the adjacent mushroom body. Overall, the number of TH-positive cells in this region was significantly reduced in dhtt-KO brains (88.93±2.76, n = 15 lobes) compared to w1118 controls (104.22±2.95, n = 9 lobes) (Fig. 5G), with a specific reduction in the number of TH-positive neurons in the PAM cluster. These results suggest a novel role for htt in the dopaminergic system as both DA levels and TH-positive cells in the protocerebrum are significantly reduced in dhtt-KO brains.
DISCUSSION
Prior research has positioned huntingtin as a key hub molecule, linking genes that undergo changes in expression following exposure to ethanol at all stages of prenatal development [10], and also established a link between alcohol misuse and HD symptom severity [11]. Here, we confirm this relationship by providing evidence that wild-type htt protein plays a role in regulating the physiological and behavioral responses to ethanol exposure. If wild-type htt plays a protective role during ethanol toxicity, it is possible that the presence of the expanded polyglutamine stretch found in HD could result in loss of a protective htt function following ethanol exposure during development or during adulthood in a dopamine-dependent way.
DA and ethanol are linked together by both immediate effects of ethanol on individual neurons via changes in neurotransmitter machinery and intracellular signaling cascades, as well as more long-term activation of reward pathways. In the short-term, ethanol exposure triggers inhibitory GABAergic synapses and the release of GABA [38]. This produces an overall inhibitory effect on the dopaminergic system, in part by interfering with DA release and receptor function [39]. Therefore, in the presence of already reduced DA levels brought about by a lack of htt, ethanol may further reduce DA levels, resulting in the behavioral phenotypes we observed in dhtt-KO flies.
Inhibiting TH activity in Drosophila leads to decreased ethanol-induced hyperactivity, which can be rescued by administering the DA precursor L-dopa [31]. DA receptors frequently couple to adenylyl cyclase and either increase or decrease cyclic AMP (cAMP) levels and protein kinase A (PKA) signaling, depending on the receptor subtype. Disruptions in these signaling pathways bring about changes in sensitivity to ethanol, such as is observed in Drosophila cheapdate/amnesiac mutants [40]. Future studies should explore changes in cAMP or PKA signaling in dhtt-KO brains and investigate if this could account for the observed decreased sensitivity, increased tolerance, or shortened recovery time.
The current study found that flies lacking dhtt exhibited a decreased preference for ethanol. The history of research on alcohol preference and dopamine is complicated. Prior work has shown that ethanol-preferring rodent strains have reduced dopaminergic function, which may be associated with an increased risk for alcohol abuse [41], but other studies have shown that interfering with more discrete components of the DA system can elicit decreased ethanol preference, including in D2-receptor-deficient mice [42] and female DA-transporter-deficit rats [43]. Krasavietz (or exba) mutant flies, which have defects in a gene encoding a translation initiation factor involved in learning and memory, show a strikingly similar phenotype to our dhtt results, exhibiting decreased sensitivity to ethanol sedation [44], decreased ethanol tolerance [44], and decreased motivation to consume ethanol [45]. The potential shared mechanisms between ethanol-related behaviors and learning/memory processes may also encompass normal roles of htt in flies, as the DA system is implicated in all three processes.
It is not known whether the loss of DA we observed in dhtt-KO brains is due to a progressive degeneration of dopaminergic cells in the protocerebrum or whether this population never formed properly during development. Future research should investigate if this loss is a degenerative process or if this is a developmental feature, including an assessment prior to our 25-day-timepoint to determine if normal numbers of DA neurons form initially in the absence of htt. We do observe a decrease in the protocerebral anterior medial (PAM) cluster, which is the largest of population of TH+neurons, projecting to the medial portion of the horizontal lobes of the mushroom body [46], but future experiments should examine in more detail how discrete populations of TH+neurons in the protocerebrum may be affected differently by loss of htt. For example, it is known that in Pink1 and parkin Drosophila mutant brains, PPL1 neurons are particularly vulnerable to degeneration [47].
Htt has many known interaction partners, providing ample opportunity for involvement with the DA system. For example, wild-type htt enhances calmodulin activity [48], which in turn modulates ethanol-induced behavioral and physiological responses, including locomotor activity and sensitization through calmodulin/Ca2 +-dependent PKA signaling [49]. Moreover, a Ca2 +-calmodulin-dependent system regulates DA synthesis largely via phosphorylation of TH [50]; low availability of calmodulin leads to low levels of DA. Thus, inhibition of calmodulin in the absence of dhtt could lead to the observed down-regulation of the DA response in dhtt-KO brains.
Alternatively, loss of htt could impact DA through changes in neurotrophin expression levels. In mammals, wild-type htt activates BDNF transcription and regulates its transport within the cell, which when disrupted, can decrease DA levels [1, 2]. Some aspects of the early DA dysregulation seen in HD could possibly be due to loss of normal htt function: BDNF deficits are found in both HD patients and mouse models [51]. BDNF promotes survival of DA neurons and can control dopaminergic receptor expression [52, 53]; conversely, DA itself can feedback to regulate BDNF levels [54]. Though BDNF itself is not found in Drosophila, several neurotrophin-like molecules have been identified, including neurotrophin 1 (DNT1), which has a similar structure to BDNF, and mesencephalic astrocyte-derived neurotrophic factor (MANF), which promotes dopaminergic neuronal survival [55, 56]. In an Alzheimer’s disease model, both amyloid-β (Aβ)42 or tauR406W expressing transgenic flies show significantly lower DNT1 expression and significant increases in ethanol sensitivity, indicating that pathological tau and Aβ aggregation can independently affect DNT1 expression and alter ethanol responses [57]. Whether dhtt regulates Drosophila neurotrophins is an important next step in determining the mechanism by which htt regulates the dopaminergic system.
Further experiments can solidify the connections between reduction in TH-positive neurons, dopamine brain levels, and sensitivity to alcohol. Dopamine-regulating drugs are one avenue to test the hypothesis that loss of dopamine can change ethanol tolerance. The administration of either the tyrosine hydroxylase inhibitor 3-Iodo-L-tyrosine (3IY) [23] or the dopamine precursor L-dopa through fly food could explore the impacts of exogenous dopamine modification in the dhtt mutant flies and investigate a potential phenotypic rescue with L-dopa. If loss of htt function caused by an expanded polyglutamine domain (as found in HD) is causing the observed differences in alcohol responses, inducible constructs of htt containing pathogenic and non-pathogenic repeat tracts can be designed to investigate if either would be sufficient to rescue those phenotypes.
This work provides a basis for further investigation into wild-type htt function in non-pathological contexts. In particular, the novel link we have established between htt and the DA system should be elucidated. More specifically, how development of the dopaminergic system proceeds in the absence of htt and the cellular mechanisms that underlie its involvement with the dopaminergic system remain to be characterized. It should be possible to construct a novel interactive molecular network using a genetic screen in which second-site genetic enhancers or suppressors are identified.
These results motivate future study of changes in addiction behaviors in the absence of wild-type htt or the presence of mutant forms of htt, and provide insights into impacts of alcohol administration on disease progression in HD animal models. Interdisciplinary collaboration between seemingly disparate fields has been a driving force behind major advances in the HD research community, and this may prove true again with the fields of addiction and FASD research.
Footnotes
ACKNOWLEDGMENTS
We would like to thank Sheng Zhang at the University of Texas Health Science Center at Houston for kindly providing the dhtt-KO and w1118 flies and genotyping assistance, Jay Hirsh and Sarah Siegrist at the University of Virginia for antibody reagents, Ed Lowry at Hampden-Sydney College for statistical consultations, Traci Stevens at Randolph-Macon College for the introduction to inebriometer use in flies, and Todd Shoborg at the NIH/University of Wyoming for helpful discussions. We thank Barry Condron at the University of Virginia for his support with initial experiments and review of the final manuscript. Initial experiments were performed in the context of E.C.’s undergraduate courses at Hampden-Sydney College (BIOL310 Developmental Biology) and the University of Virginia (PSYC4559: Research Methods: Scientific Writing in Behavioral Neuroscience). Parts of this work were conducted in the lab of Sarah Siegrist (which was supported by NIH R01-GM120421), where C.S. was supported by a National Research Service Award from the NIH/National Institute of Neurological Disorders and Stroke (F32-NS096919). We would also like to acknowledge the authors of previously published foundational studies not cited in this manuscript due to space constraints. Finally, we acknowledge the Monacan Nation as the traditional owners of the land upon which this research was conducted.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
DATA AVAILABILITY
The data supporting the findings of this study are available from the corresponding author on request.