
Abstract
In this study, the objective is to model a neuronal population using cellular automata (CA) rules that are both simple and realistic. Unlike in previous research, the primary aim is to capture a range of neuronal behaviours with minimal complexity and computational overhead. To achieve this, a two-dimensional lattice of cellular automata was implemented in the C
Introduction
Modelling and simulating complex systems have been the subject of research for decades [1–3]. Differential and state equations are usually employed by researchers to quantify and model dynamic systems [4]. Linear differential equations are suitable for linear and deterministic systems, allowing the prediction of system behaviour based solely on the initial conditions. However, in nonlinear systems, mutual effects and nonlinear behaviour should be considered [1]. One of these complex nonlinear systems is the human brain, comprising various mutual spatial networks [5]. These networks consist of numerous nonlinear units known as neurons, which, at any given moment, fire or cease firing following specific patterns, thus giving rise to spatiotemporal dynamics within the entire system. Nevertheless, the collective behaviour of these neurons gives rise to spatial and temporal brain activity patterns in various dynamic states [1, 6–8]. Contemporary functional neuroimaging modalities such as quantitative electroencephalogram (QEEG), functional magnetic resonance imaging (fMRI), and positron emission tomography (PET) reflect the activity of thousands of neurons [9]. However, these modalities do not offer a mathematical or physical theory to explain the underlying mechanisms governing the collective activity of neuronal populations [4, 10, 11]. Computational bio-models of the collective activities of neuron populations are imperative for comprehending macroscopic brain data and gaining detailed insights into the mechanisms at play in applied neuroscience. Thus, computational modelling has emerged as a compelling field within computational neuroscience, providing neuroscientists with a tool to explore the functioning of neuron populations.
An attractive mathematical algorithm for the biological modelling of neuron activity is offered by Cellular Automata (CA) [12]. The modelling of complex systems was made simpler with the introduction and development of CA, resulting in the incorporation of a spatial dimension into these models. Unlike differential equations, CA-based models do not involve numerous parameters and exhibit significant computational efficiency [13, 14]. In contrast to neural networks, which operate as black boxes and come with high computational costs, the events and processes of CA-based models can be monitored by defining CA rules, allowing for the observation of the effects of even the slightest variations in parameters [12, 14]. Consequently, the proposal of a simple one-dimensional CA model to replicate patterns of brain waves in EEG signals was put forth [15]. Similarly, Gevertz and Torquato demonstrated the modelling of vasculature evolution within brain tumour growth by employing a combination of CA and reaction-diffusion equations [16]. Shrestha et al. simulated the large-scale growth of tumours by utilizing a combination of CA and steady-state equations [17]. More recently, abnormal and normal EEG signals were simulated by Acedo and his colleagues through the use of a large number of automata and dense connections between them [18]. However, their CA model is afflicted by a substantial computational cost, necessitating distributed computation. Tsoutsouras et al. introduced a two-layer CA model (50
This article represents the initial phase of a broader brain modelling project. Our objective at this stage is to model a population of neurons using rules that are as realistic as possible within the framework of CA, allowing for the modelling of various behaviours of a neuron population with minimal complexity and computational demands, a departure from previous studies. In essence, a straightforward economic model with the capabilities of prior models is intended to be developed. Furthermore, in contrast to previous studies, a greater emphasis will be placed on exploring the various behaviours of the model and its associated parameters.
Methods
Cellular Automata (CA)
CA is a discrete and nonlinear algorithm comprising several components with the capacity to manifest complex behaviour when interacting [12]. In essence, CA is composed of a network of interacting neurons governed by specific rules. Each neuron possesses various states and is situated within a defined neighbourhood. CA is characterized by four fundamental concepts, namely states, rules, a lattice of sites, and neighbourhoods. As a result, CA can be described by [21].

Where
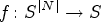

At times, a function cannot be determined for a specific application, and a rule, which is more general than a function, is defined to derive the next state of each cell. Consequently, the general formula for updating a cell’s state in CA is provided as follows:
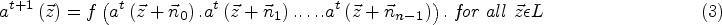
The model was implemented with the C
Lattice and cell
In the current research, the aim was to simulate the activity of a population of neurons and their interactions. To achieve this, a two-dimensional network was considered, where each site is represented by a neuron. For the sake of simplicity and cost-effectiveness in calculations, the size of this network was set to 20
States
Three major types of behaviour are exhibited by a neuron: rest, action potential (AP), and spike [22]. To maintain simplicity, the focus of this research is solely on modelling the rest and AP behaviours, with spike falling outside the scope of this study. Consequently, each cell is characterized as a neuron with two distinct types of behaviour.
At rest, a neuron’s potential remains constant at
The simplified CA model of neuron behaviour, encompassing rest and AP states, is depicted in Figure 1. Different states of the neuron are defined by a set of
Expression of the action potential as a discrete pulse divided into 11 states for determining the conditions of neurons within the cellular automata lattice.
The human cerebral cortex contains more than 1010 neurons [22]. These neurons continuously interact and communicate with each other to perform their respective functions. Research has demonstrated that each neuron receives between 1,000 and 100,000 synapses and transmits signals to a significant number of neighbouring neurons [22, 23]. To account for this, two parameters,
Rules
A set of rules is required to model the interaction among neurons. Within this subsection, three fundamental rules governing neuron interactions are introduced:
Rule 1. Neuron activation, transitioning from its resting state, occurs when the number of active neighbours equals or exceeds
Rule 2. Following activation, neurons must progress through the absolute refractory period and the relative refractory period before returning to the resting state.
Rule 3. During the absolute refractory period, neurons are not susceptible to reactivation through neighbouring stimuli. However, during the relative refractory period, neurons may be activated if stimuli from their surroundings reach
Results
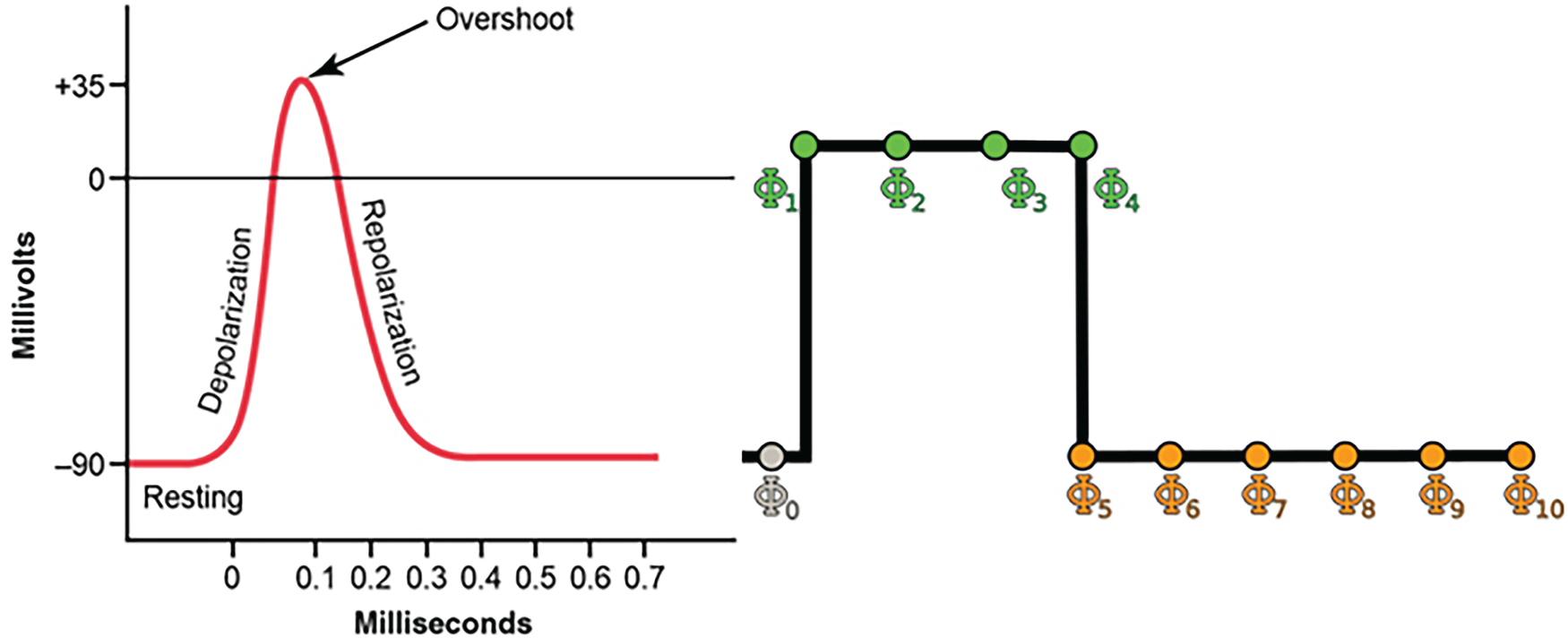
To implement the model, the initial step involves determining the distribution of neurons across resting, absolute refractory, and relative refractory states. Following a process of trial and error, the following distribution was selected for neurons in each state: 30% of neurons in the resting state, 45% of neurons in the absolute refractory state, and 25% of neurons in the relative refractory state. Upon implementation, four distinct model behaviours were observed: fixed point, rhythmic, chaotic, and dual behaviour. Figure 2 illustrates an example of a fixed-point output from the model. In this scenario, the output of the neuronal CA converges to zero for various lattice parameter values, indicating that all neurons return to the resting state and the lattice dynamics reach a stable fixed point.
Fixed point behaviour of the model; loss of lattice dynamics for the parameters shown above the graph. The vertical axis shows the amplitude in volts.

Rhythmic behaviour of the model; an example of a rhythmic time series output of the model. The vertical axis shows the amplitude in volts.
A rhythmic behaviour of the model was observed by altering the count of neighbours of the neurons and adjusting the thresholds of neuronal stimulation. In Figure 3, an example of the model’s rhythmic signal outputs is depicted. To gain a better understanding of these behaviours, Figure 4 illustrates the phase-space reconstruction [24, 25], power spectrum [26], and recurrence plot [27, 28] of such outputs.
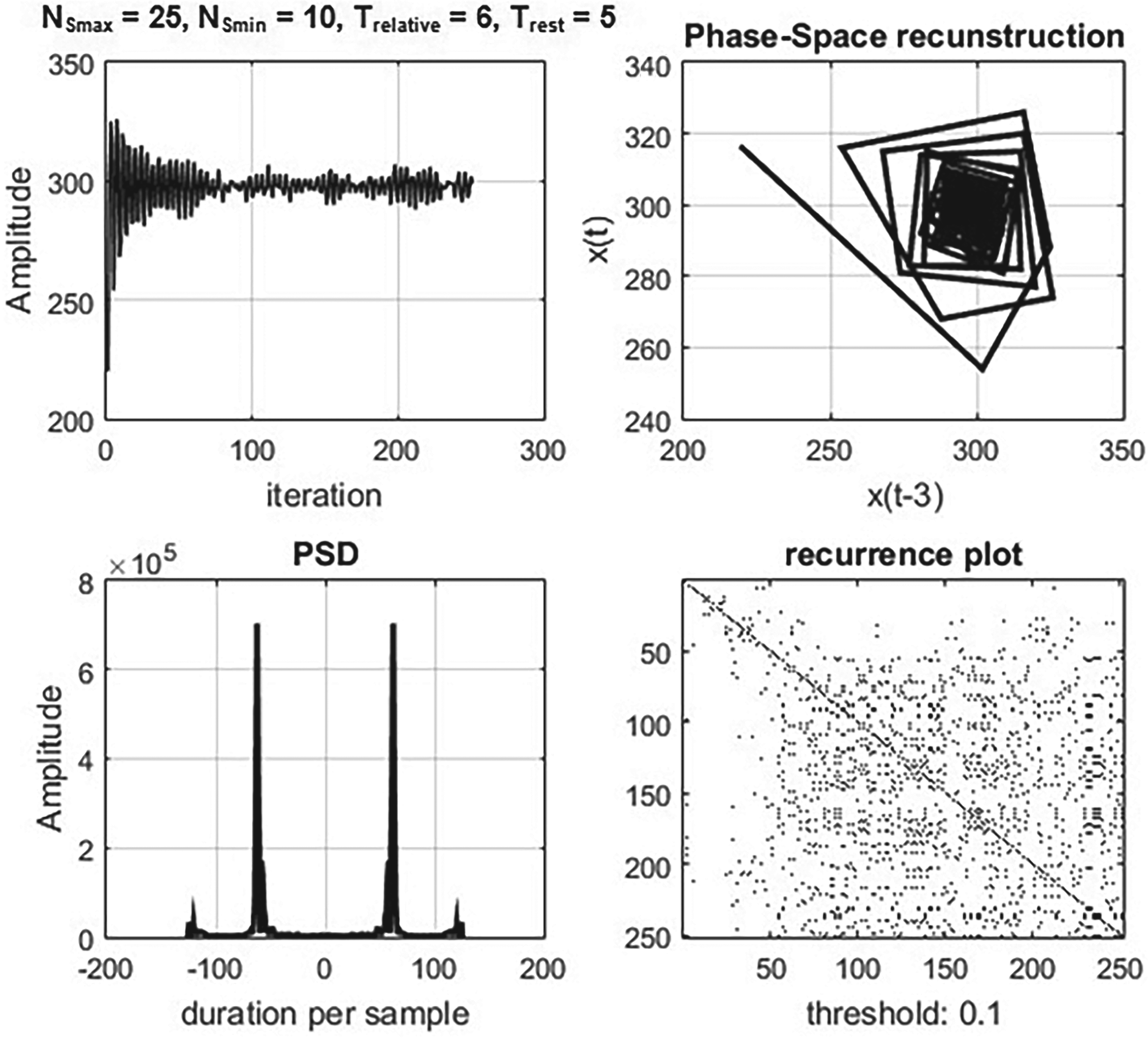
Phase-space reconstruction, power spectrum and recurrence plots of a rhythmic signal output of the model.
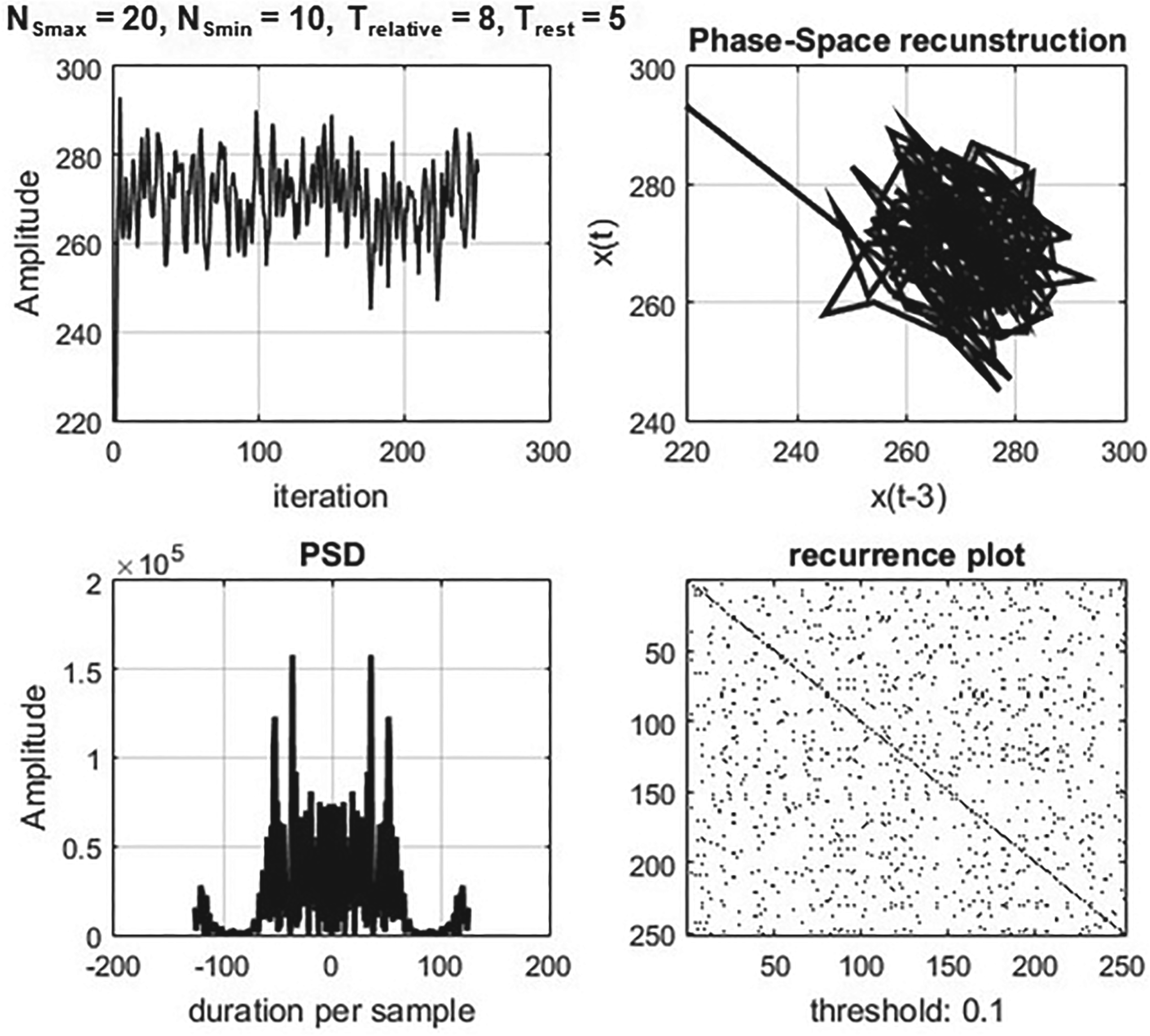
Phase-space reconstruction, power spectrum and recurrence plots of a chaotic time series output of the model.
A sample of a chaotic output of the model is depicted in Figure 5. This figure displays the phase-space reconstruction, power spectrum, and recurrence plot of the chaotic outputs.
The results revealed that the maximum and minimum number of neighbours of the neuron had significant effects on the neuronal CA outputs. Therefore, an examination of these parameters was conducted by varying their values. In Figure 6, the output time series of the lattice is presented while keeping the parameters

The effect of increasing
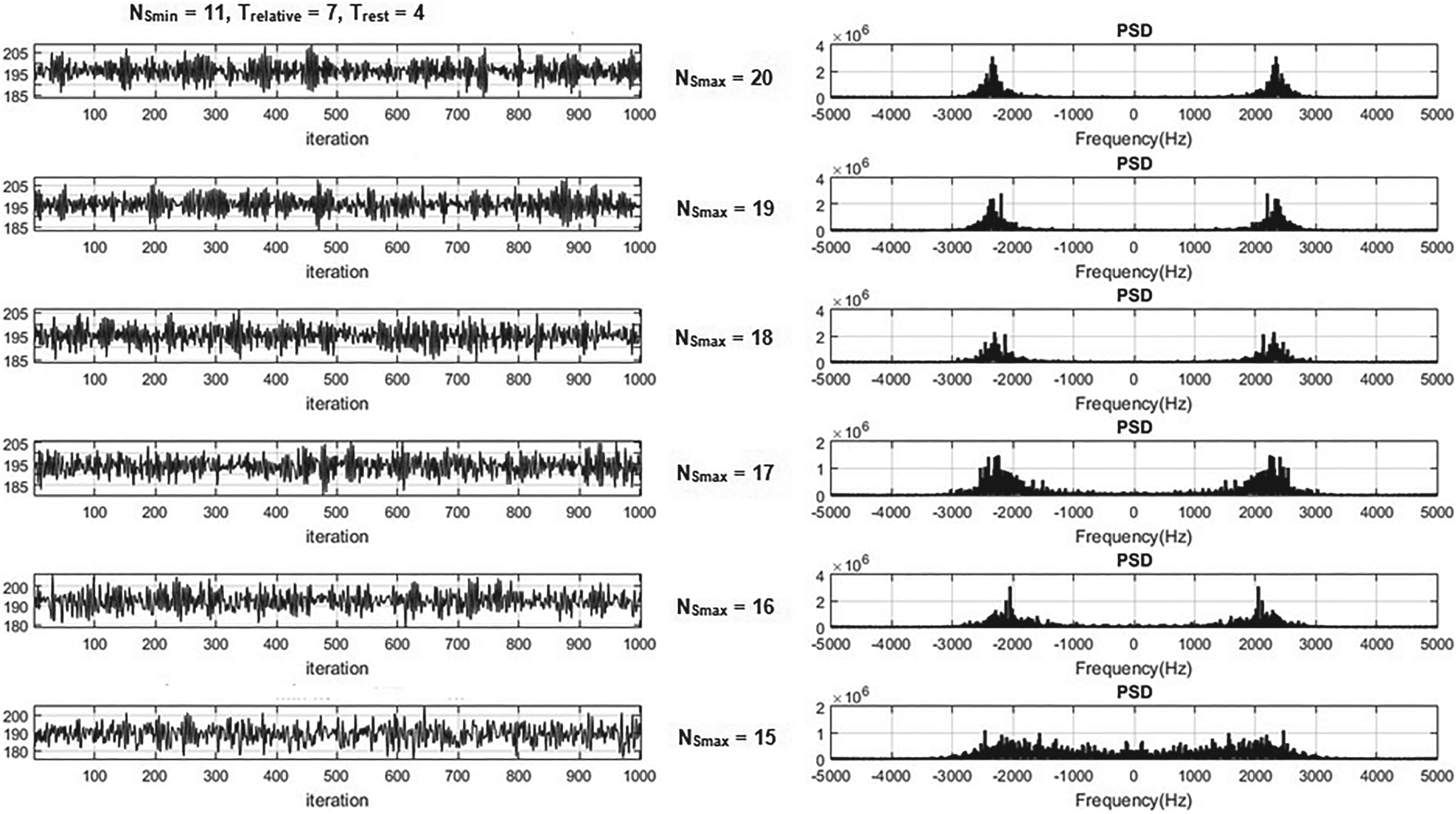
The effect of decreasing
Surprisingly, an interesting phenomenon was observed with a further reduction of
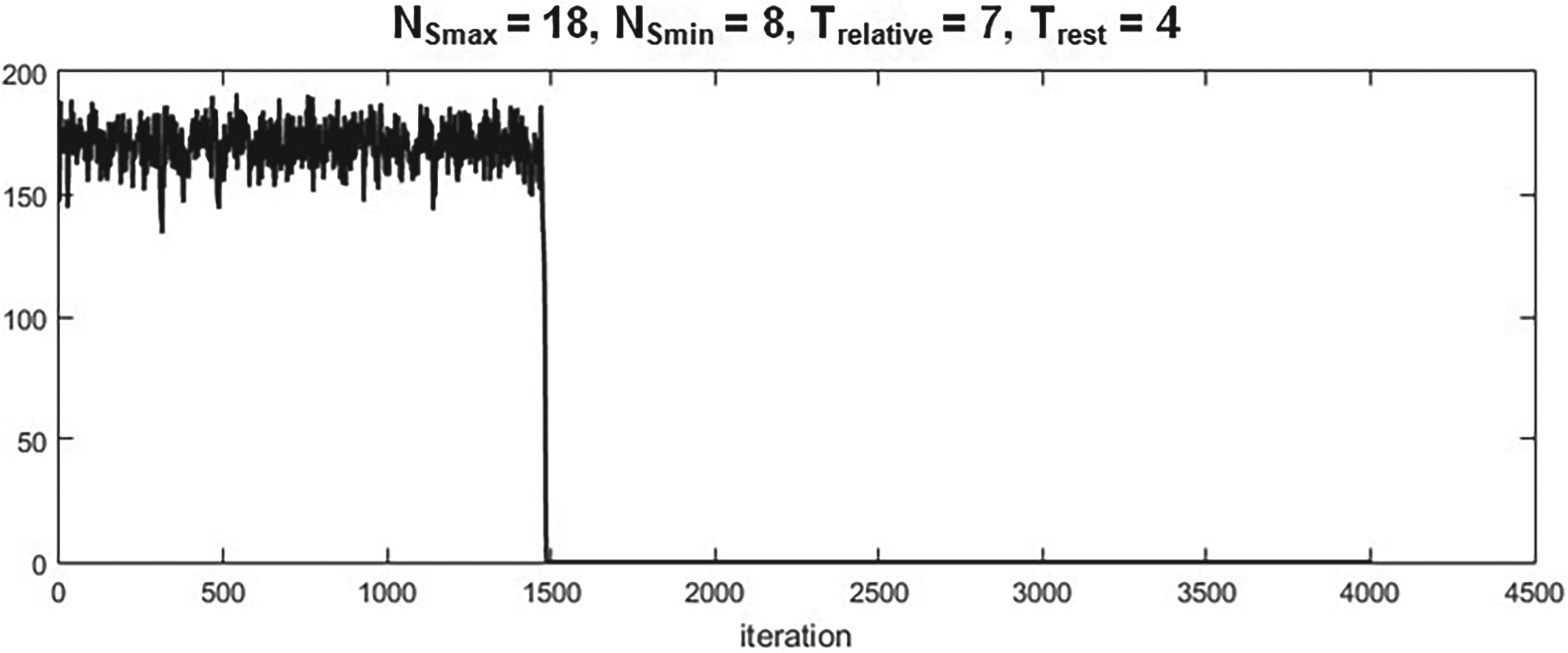
Dual behaviour phenomenon. As the maximum number of neighbours of a neuron decreases, a dual behaviour occurs before the loss of lattice dynamics.
Cellular Automata (CA) play a crucial role in statistical mechanics and is applicable in various complex dynamical systems. However, limited research has been undertaken on the utilization of this concept for modelling and simulating brain activity, which stands as an exemplary instance of a highly intricate system. Additionally, only a few studies have attempted to simulate electrical waves in the cerebral cortex using the CA approach, and these studies have been carried out with notable constraints. For example, a model was proposed by Beigzadeh and Hashemi [29] that incorporated significant details of a neuronal population in the visual cortex, successfully generating various dynamics with a frequency spectrum akin to a chaotic time series. Nevertheless, their model was plagued by excessive complexity and numerous parameters that required meticulous tuning and extensive efforts. Furthermore, an accurate nonlinear analysis of the model outputs or a direct comparison with real data was not provided by the authors. In a probabilistic CA model constructed by Acedo et al. [18], a substantial network of neurons (106) was created based on specific aspects of an action potential and a neuronal population in the brain. Despite the limited parameterization of the model, the large number of neurons resulted in a computationally intensive process, necessitating the use of distributed computing resources. In contrast, in their study, Goltsev et al. [30] demonstrated that small networks comprising 50–1000 neurons within CA networks could exhibit oscillations reminiscent of those observed in larger networks. In our study, a progressive approach was taken by incorporating four distinct states to simulate the functionality of neurons accurately. In this study, the effectiveness of cellular automata was investigated in replicating the behaviour of a population of neurons and simulating electrical waves within the cerebral cortex. To achieve this, novel and straightforward rules for the model were developed, which minimized not only computational burdens but also reflected realistic behaviours of the neuronal population. Taking inspiration from the action potential phenomenon, three distinctive states for each neuron were successfully modelled: resting, firing, and refractory states. Our simulations demonstrated that the proposed model could generate various behaviours and dynamics, ranging from fixed points to chaotic behaviours observed in a real cortex under different circumstances.
The output of the model was found to be highly dependent on the number of neighbours for each neuron. As observed in the results, as
A distinct spectral pattern resembling 1/f, indicative of self-organized critical dynamical systems akin to the brain [7], was exhibited by the simulation data. Furthermore, a transition in the system’s dynamic behaviour was observed by modifying the minimum and maximum number of neighbourhood connections or cell synapses. This transitional state, known as the critical state or the edge of chaos, exists between two distinct behaviours [31]. In this critical state, the system operates at a state of criticality [31] and is recognized for its optimal memory and information processing capabilities [32]. Several theoretical neuroscience studies propose that phase transitions in the brain, occurring within large finite systems, manifest not as discrete points but rather as narrow regions encompassing many criticality characteristics [33]. Similarly, in our CA network, this phase transition was not limited to a specific point but instead took place within a small range of the aforementioned parameters. Therefore, the potential for our model to serve as a versatile instrument for a multitude of clinical purposes is evident. With its foundation rooted in established physiological knowledge and mathematical principles, valuable assistance is offered by our model in simulating and enhancing comprehension of the brain’s functionality across diverse scenarios. Moreover, it holds significant promise for medical research and education, as it facilitates the exploration of various hypotheses pertaining to the human brain.
One of the limitations of this study is that the current model only simulates a small neuronal population. Additionally, the model lacks the inclusion of important parameters, such as time delays in neuronal responses. Furthermore, the absence of real brain data in the model represents another limitation in the present study. Another critical limitation is the model’s failure to consider different types of brain synapses. The model is inherently constrained by its primary applicability to short-term processes related to brain electrical activity, making it unsuitable for accurately representing longer-term dynamics. To address this limitation and enhance its overall adaptability, one potential approach involves dynamic adjustments of model parameters within the framework of a probabilistic cellular automaton. Moreover, it is essential to acknowledge that the validation process for such models often presents significant difficulties and complexities.
Conclusion
In the current research, the ability of CA to model the behaviours of a neuron population was explored. In this study, new, novel, yet straightforward rules for the CA network were employed to keep computational loads low and exhibit sensible behaviours of the neuron populations. By considering the electrical activity of a real neuron, three statuses for every cell (i.e., resting, absolute refractory, and relative refractory statuses) could be modelled. Our model features four controllable variables (the minimum number of neighbours of the neuron, the threshold for stimulating the neuron in rest, the maximum number of neighbours of the neuron, and the threshold for stimulating the neuron in the relative refractory period), which could demonstrate fixed point, rhythmic, aperiodic, and chaotic behaviours. As a result, our model can be utilized to investigate various neuronal populations and provide more insight into their different mechanisms. For instance, based on our results, particularly the observation of dual behaviour, the mechanisms involved in the demise of a biological neuronal population, such as brain death, can be described using the proposed model. Our research group is actively working on improving, optimizing, and developing this model. In the future, different sizes of the CA lattice will be examined, and efforts will be made to determine an optimal range for each parameter of the model in line with development goals. Furthermore, model rules will be developed, and other components of the neuron population, such as inhibitory synapses and time delays in neuronal responses, will be introduced into the model. The ultimate goal is to model various components of the human brain. Additionally, it’s worth noting that the proposed model currently examines the activity of neurons only in the resting state. Therefore, future studies should consider different inputs, equivalent to various types of sensory and motor brain inputs, for the proposed model and assess its performance in comparison with real data.
Footnotes
Funding
National Key Project of the 14th Five Year Plan for Educational Research by the Ministry of Education, this work was supported by the National Teacher Research Special Fund Management Office of the Ministry of Education.