
Abstract
Alginate is a polysaccharide derived from Sargassum sp. and is a potent immunostimulant with antibacterial activity, including against Vibrio spp. This genus of bacteria is found in freshwater and marine environments and is a common infectious, pathogenic bacteria both for aquatic cultivans and humans. Here, we determined the ability of sodium alginate polysaccharides and oligosaccharides (AOS) to act as immunostimulants in Artemia challenge tests and antibacterial diffusion disc assays against Vibrio parahaemolyticus, V. vulnificus, and V. harveyi. The AOS was produced by thermal heating. Dry sodium alginates were weighed out from 4.21 to 6.47 grams with a yield varying from 21.05 to 32.35%. Alginate polysaccharides were challenged against V harveyi and showed 8 positive results. The highest inhibitor zone was 12.962±3.623 mm. Based on 18 tests, AOS showed 12 positive results, with the highest inhibitor zone being 10.250±0.09 mm. The encapsulated alginate against Vibrio parahaemolyticus, Vibrio harveyi, Vibrio vulnificus, and the non-challenged tests without any Vibrio spp. addition resulted in the best concentrations of 800 ppm (polysaccharide) and 600 ppm (oligosaccharide), respectively. The lower concentration of oligosaccharides alginate were more effective and has the potential to be superior as an antibacterial agent and immunestimulant, as opposed to alginate polysaccharide.
Keywords
Introduction
Alginate mostly found in the cell wall of brown seaweed includes Sargassum sp [1]. The structure is characterized by its G/M ratio, the arrangement of uronate residues, the degree of acetylation, and the molecular weight (MW) [1]. Alginate oligosaccharides (AOS) is the degradation product of alginates and has a lower MW [2]. In plants, AOS can act as a growth promoter and improves resistance to pathogens and stressors such as heavy metals by triggering plant immunity through a pathogen-associated molecular pattern [3]. AOSs have also served as prebiotics for lactic acid bacteria for humans and animals [1, 4]. Finally, alginate has been shown to be a potent immunostimulant [5] and antibacterial agent [6].
Vibrio spp. bacteria live in freshwater and marine environments [7]. The bacteria are commonly found in freshwater commercial hatcheries [8], water and sediment freshwater fish farms [9], waters and sediment estuaries [10], on the sea-surface (pelagic) in the Mid-Atlantic [11], in 1200 m deep-sea mud [12], and in deep-sea seamount [13], in addition to being in seafood [14]. Most Vibrio spp. are pathogenic bacteria [15], namely Vibrio vulnificus [16], Vibrio parahaemolyticus [17], and Vibrio harveyi [18]. Indeed, natural products play a crucial role in treating the biological systems, in which considerable attempts have been always dedicated to provide insightful information about their various sides concepts [19–21]. Accordingly, several methods including experimental and bio informatics based methods have been developed to approach the goal [22–24]. It is important to mention that investigating interactions between potent compounds and biological related systems could help to make progress on the drug design aspects [25–27]. Progress of serious diseases in recent years pushed forward such investigations on developing therapeutic agents [28–30]. Therefore, we did this work to show advantages of employing AOS compound for working as antibacterial agent V. vulnificus causes the diseases like septicemia [31] and necrotizing wound infections [32] in fish and humans. The infection of V. vulnificus occurs through contact with seawater or raw seafood [33]. In 1988–1997, 345 sporadic V. parahaemolyticus infections were reported as gastroenteritis (59%), wound infections (34%), septicemia (5%), and other exposures (2%). V. parahaemolyticus infections have occurred due to the consumption of raw or undercooked shellfish and exposure of wounds to seawater [34]. Consuming raw or undercooked seafood contaminated with V. parahaemolyticus can lead to acute gastroenteritis characterized by diarrhea, headache, vomiting, nausea, and abdominal cramps [35].
V. harveyi has infected vertebrates and invertebrates such as shrimp and fish [36]. V. harveyi isolates have been obtained from a diversity of hosts and geographic locations. Fourteen pathogenic isolates have been extracted from rainbow trout, Oncorhynchus mykiss (Walbaum), and Atlantic salmon (Salmo salar L.). This Vibrio sp. resulted in 100% mortality after intraperitoneal injection of 106 fish cells. Previous reports have shown that extracellular products from five pathogenic bacteria were harmful to fish [37]. Moreover, two researchers investigated five isolates of V. harveyi from 256 suspended sediments collected from the sea coast and exalted shrimp ponds. Work from our lab and others have shown that these bacteria were resistant to ciprofloxacin, penicillin, rifampicin, and vancomycin [38]. Vibrio spp. were also resistant to beta lactam-type of antibiotics, specifically Ampicilin, Amoxicilin, Co-Amoxiclav, Ciprofloxacin HCL, and Azithromycin.
Previous research by Castillo et al. [39] constructed a database of 5,674 prophage-like elements covering 1,874 Vibrio genome sequences. Sixty-four species showed properties such as virulence and antibiotic resistance that were widely distributed among environmental vibrios, whereas other strains were classified as non-pathogenic.
Here, we determine the ability of sodium alginate polysaccharides and oligosaccharides (AOS) to act as an immunostimulant in Artemia challenge tests and antibacterial diffusion disc assays against Vibrio spp., namely V. parahaemolyticus, V. vulnificus, and V. harveyi.
Experimental methods
Sample collection, preparation and alginate extraction
The fresh Sargassum sample was cleansed from debris, dirt and rinsed from salts under tap water. The cleansed macroalgae was the dried up in normal temperature and protected under direct sun exposure. Alginate preparation was carried out using the sodium alginate pathway as described by Yudiati and Isnansetyo [40]. Firstly, air-dried brown algae (Sargassum sp.) was crushed using a commercial blender. Then, 20 g of Sargassum sp. powder was weighed and mixed with 500 mL aquadest, 5% sodium carbonate (Na²CO3), and stirred up for 1.5 hr. pH was checked and adjusted to 8.5 with hydrochloric acid (HCl). Alginate solution was stirred for up to 24 hours. The extract was filtered, the supernatant was added with 0.13 M potassium chloride (KCl) and 96% alcohol at a ratio of 1:1 (v/v) was added [29]. The mixed solution was centrifuged to separate the precipitate from the liquid, and the pellet was dried in a 55°C drying cabinet for 24 hours. This procedure resulted in alginate polysaccharides. Oligosaccharides (AOS) were obtained by heating in an oven laboratory at 145°C for 4.5 hours [2]. In our study, the sodium alginate was fit to the standard alginate (Sigma®, USA) [41] and was 217.5 KDa in MW with an 89.95% degree of acetylation [2].
Strains of bacteria
In this study, Vibrio spp. were collected from the Laboratory of Biology, Faculty of Fisheries and Marine Sciences, Diponegoro University. These bacteria were isolated directly from shrimp ponds. There were three types of Vibrio spp.: Vibrio parahaemolyticus, Vibrio vulnificus, and Vibrio harveyi.
Preparation of artemia and bacteria culture
For preparation of culture, we used commercial Artemia salina cyst Supreme Plus® (Golden West Artemia). Prior to use, all glassware materials and medium were sterilized in an autoclave. All plastic materials used for experiments were sterilized with 70% antiseptic alcohol and UV light disinfectant. All trials were performed inside the controlled Laminar Air Flow. Artemia was hatched in a 1000 mL black plastic bottle equipped with continous aeration for 24 hrs at 25 ppt. [42].
Vibrio spp. was cultured in nutrient broth for 24 hrs at 28 °C in 25 ppt salinity. The nutrient broth consisted of 5.0 g peptone, 5.0 g NaCl, 2.0 g yeast extract, 1.0 g beef extract, and 15.0 g agar (for solid media) [43].
Antibacterial test
Alginate polysaccharides and oligosaccharides were prepared at a concentration of 1000 ppm. We used the paper disc (Whatman no 1, diameter 5 mm) diffusion described previously by Bayer et al. [45]. Nutrient agar consisted of 5.0 g peptone, 5.0 g NaCl, 2.0 g yeast extract, 1.0 g beef extract, and 15.0 g agar [43]. These compounds were diluted in one liter saline water for medium culture and tests. The antibacterial test used 70% alcohol as a positive control. Bacterial density was adjusted to the standard 0.5 Mc Farland (1-2×108 CFU).
Bacteria grown in liquid media were swabbed using sterile cotton on NA solid medium. The paper disc was inserted into 1000 ppm alginate and dried at room temperature in a petri dish. Afterwards, we put the paper disc into a petri dish filled with bacteria and incubated dishes for 24 hours at 37 °C. For our results, we measured the inhibition zone surrounding the paper disc area using a digital caliper. All experiments were performed in triplicate.
Artemia gnotobiotic challenge test
Alginate polysaccharides and oligosaccharides were prepared at a concentration of 400, 600, and 800 ppm. Similar to methods described by Allender et al. [45], nauplii Artemia were immersed in alginate solution for 1 hr. Addition of Vibrio spp. was performed by placing the bacteria into vial bottles at the standard McFarland concentration (1-2×108 CFU/mL). Ten Artemia nauplii were placed into the vial containing Vibrio spp., and dead Artemia were counted every six hours [46]. The cumulative mortality index (CMI) of Artemia was calculated by summing the mortality counts noted at each time interval [46, 47].
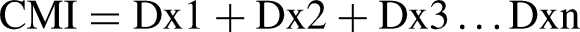
Where D is the number of individual mortalities at the time x1, x2, x3...xn. The mortality reduction was determined based on controls which were Artemia non-Vibrio for non-Vibrio treatment and Artemia + Vibrio for treatment + Vibrio. We modified these controls from Immanuel et al. [46]. Experiments were performed in triplicate.
Results and discussion
Alginate is composed of mannuronic acid (M-block) and guluronic acid (G-block) [40]. The number of block types varies between the alginate and the irregular or random arrangement of the two monomers [48]. Sodium alginate from Sargassum latifolium collected during late spring from the intertidal zone of Hurghada, Egypt had a M/G ratio ranging from 0.94–2.7 [49]. Another report of alginate polysaccharides from brown algae Stypocaulon scoparium collected from the Mediterranean coast of Algeria had a D-mannuronate to L-guluronate (M/G) ratio of 0.92. Acid hydrolysis of alginate using the HPAEC technique, on the other hand, showed an M/G ratio of 0.6 and 50% yield [50]. In this study, results from six extractions are show in Table 1.
Yield of sodium alginate extracted from 20 g dry Sargassum sp. powder.

Yield of sodium alginate extracted from 20 g dry Sargassum sp. powder.
Based on the data presented in Table 1, sodium alginate’s wet weight was 136.56–161.56 grams. Sodium alginate’s dry weight was 4.21–6.47 grams, and the obtained yield was 21.05%–32.35%. Previous research on similar alginate from Sargassum latifolium by Fawzy et al. [49] showed that sodium alginate yield ranged from 18.9% to 40.43%. Another previous study on three different types of alginate of S. siliqosum (acid, sodium and neutral alginate) found a yield range of 4.8%–40.34% [40]. Finally, work by Rhein–Knudsen et al. [51] with six types of seaweed, showed alginate yields of 16±0.6% for Padina gymnospora, 21±0.5% for Padina antillarum, 29±1.1% for Laminaria digitate, 22±1.7% for Sargassum vulgare, 27±1.0% for Macrocystis pyrifera, and 30±1.4% for Sargassum natans. Results from antibacterial test varied, and the results are presented in Table 2.
Performance of antibacterial tests measuring zone clearance inhibition on different types of alginate and different strains of Vibrio spp. with alcohol antiseptic as a positive control.
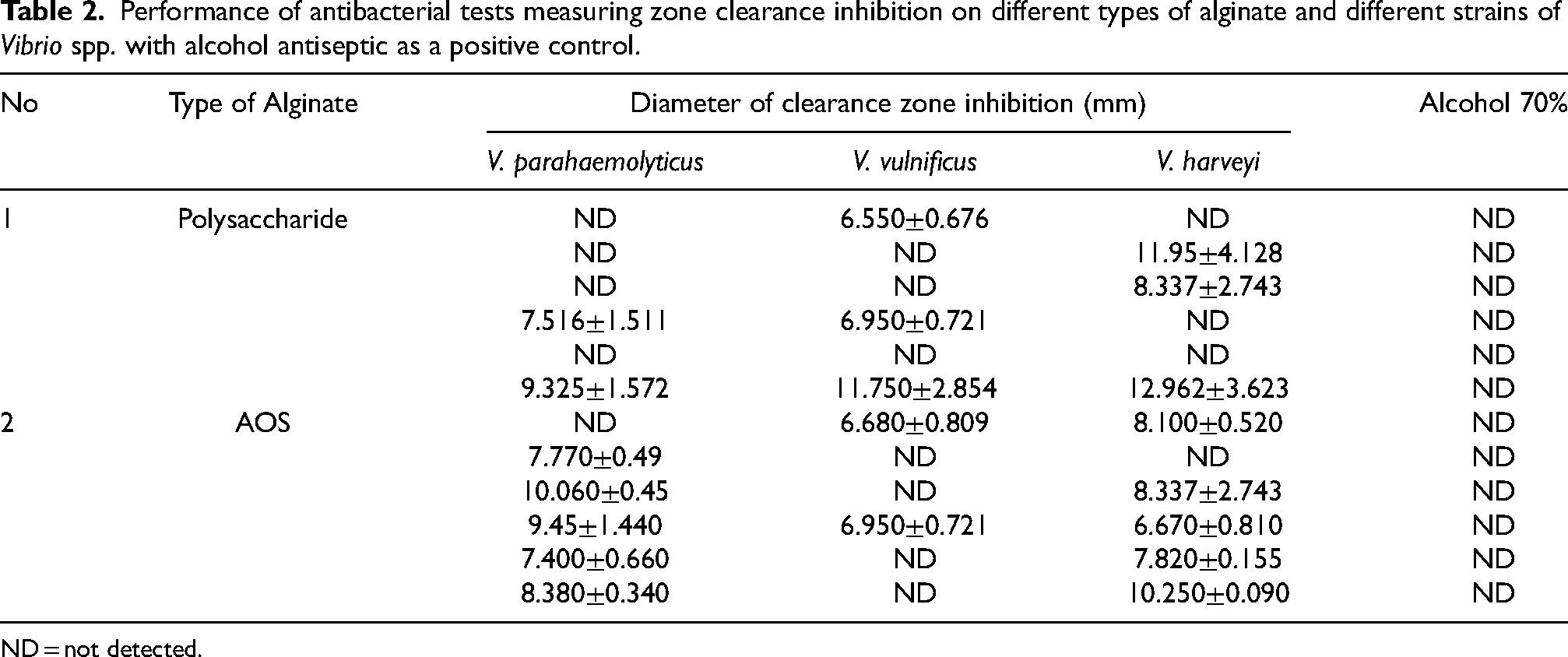
ND = not detected.
Based on the results shown in Table 2, the antibacterial activity test of Sargassum sp. polysaccharide alginate resulted in eight positive results. The highest antibacterial activity was observed against V harveyi at 12.962±3.623 mm. The lowest activity was observed for V. vulnificus, with challenge test results measured at 6.550±0.676 mm. Intermediate antibacterial acitivity was observed from polysaccharide alginates against V. parahaemolyticus, V. vulnificus, and V. harveyi,.
The antibacterial activity of AOS samples gave 12 positive results. The highest antibacterial activity came from the V. harveyi challenge test (10.25±0.09), while the lowest was obtained from the similar V. harveyi test (6.670±0.810 mm). Other results measured from the oligosaccharide alginate to antibacterial test were against V. parahaemolyticus, V. vulnificus, and V. harveyi. Zhang et al. [3] reported that AOS is more attractive with respect to biological activity because of its low MW. The MW of AOS in this study was relatively low, at 217.5 KDa, suggesting that this gives better antibacterial activity compared to alginate polysaccharides [2].
Research by Hu et al. [6] reported that 14 out of 19 tests gave positive results against V. vulnificus and V. harveyi. Conversely, V. parahaemolyticus had negative test effects. Antibacterial activity can be increased by synthesing the extracts into nanoparticles at certain combinations. For example, sodium alginate synthesis using the microwave-assisted method from synthesized silver nanoparticles has significant antibacterial activity against two types of Gram bacteria [52]. Silver nanoparticles alginate (AgNPs) show potent antibacterial activity against Staphylococcus aureus and Escherichia coli, as they cause cell death by increasing membrane permeability and compromising bacterial wall integrity [53]. The chitosan/sodium alginate (CS–ALG) combination increases the antibacterial activity against E. coli and S. aureus. Carboxymethyl chitosan–ZnO (CMCS–ZnO) nanoparticles and sodium alginate (SA) developed with a new bio-based film combined has managed to exhibit distinct antibacterial activity against S. aureus and E. coli as well as increase tension for better moisture resistance [55].
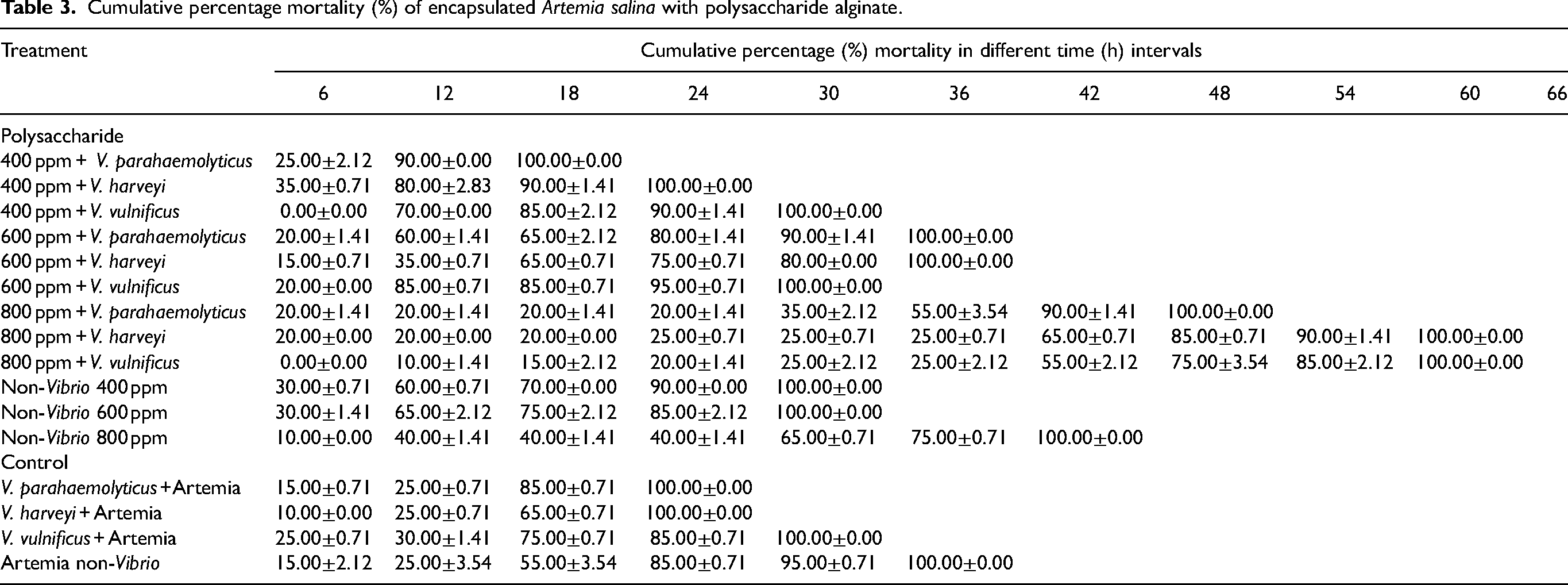
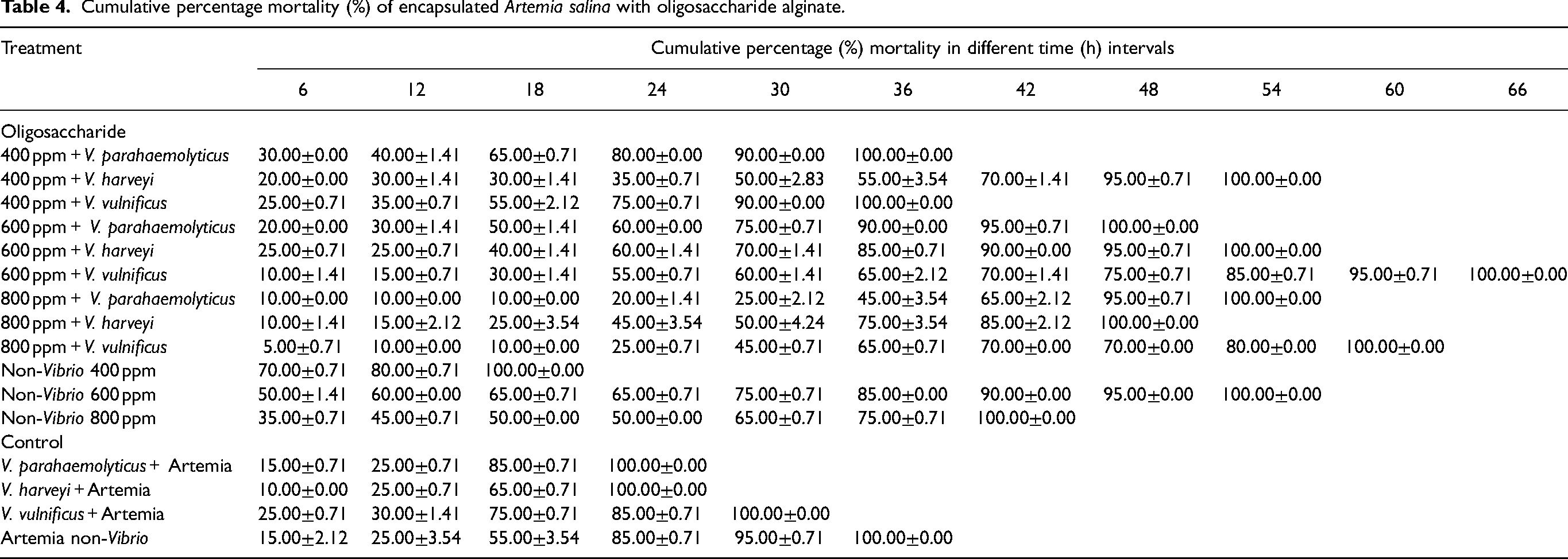
Tables 3 and 4 show the results of Artemia salina encapsulated with alginate polysaccharide and AOS challenged against V. parahaemolyticus, V. harveyi, and V. vulnificus. The results show a difference in the mortality percentage in polysaccharide alginates (Fig. 1A) and AOS (Fig. 1B). Challenged uncapsulated samples gave the highest mortality, at 30% mortality after 12–18 hrs and 18–24 hrs and 100% mortality at the 36 hr time point. The challenged Artemia against V. parahaemolyticus, V. harveyi, and V. vulnificus had the highest mortality (60%, 40%, and 45%, respectively) at similar periods (12–18 hrs). The challenged Artemia against V. parahaemolyticus and V. harveyi resulted in 100% mortality at 24 hrs while the challenge against V. vulnificus had 100% mortality at 36 hrs.
Cumulative percentage mortality (%) of encapsulated Artemia salina with polysaccharide alginate.
Cumulative percentage mortality (%) of encapsulated Artemia salina with oligosaccharide alginate.

Percent mortality of alginate polysaccharide (A) and oligosaccharide (B) capsulated Artemia, challenged and unchallenged against Vibrio parahaemolyticus, Vibrio harveyi, Vibrio vulnificus.
In alginate polysaccharide treatment, the highest mortality percentage (30% –70%) occurred at different hours post infection in different Vibrios, with 100% mortality (at 18–60 hrs). In AOS tretment, the highest mortality (20% –70%) occurred at different hours post infection in different Vibrios, with 100% mortality at 18–66 hrs. Rico-Mora and Voltolina [44] reported that Artemia franciscana in five non-pathogenic isolates of Skletonema costatum were challenged. The Artemia were also challenged with Vibrio parahaemolyticus and Vibrio alginolyticus. They found that 100% mortality of Artemia for five isolates occurred on the fourth day. Results showed that 100% mortality against Vibrio parahaemolyticus occurred at 48 hrs and against Vibrio alginolyticus at 24 hrs.
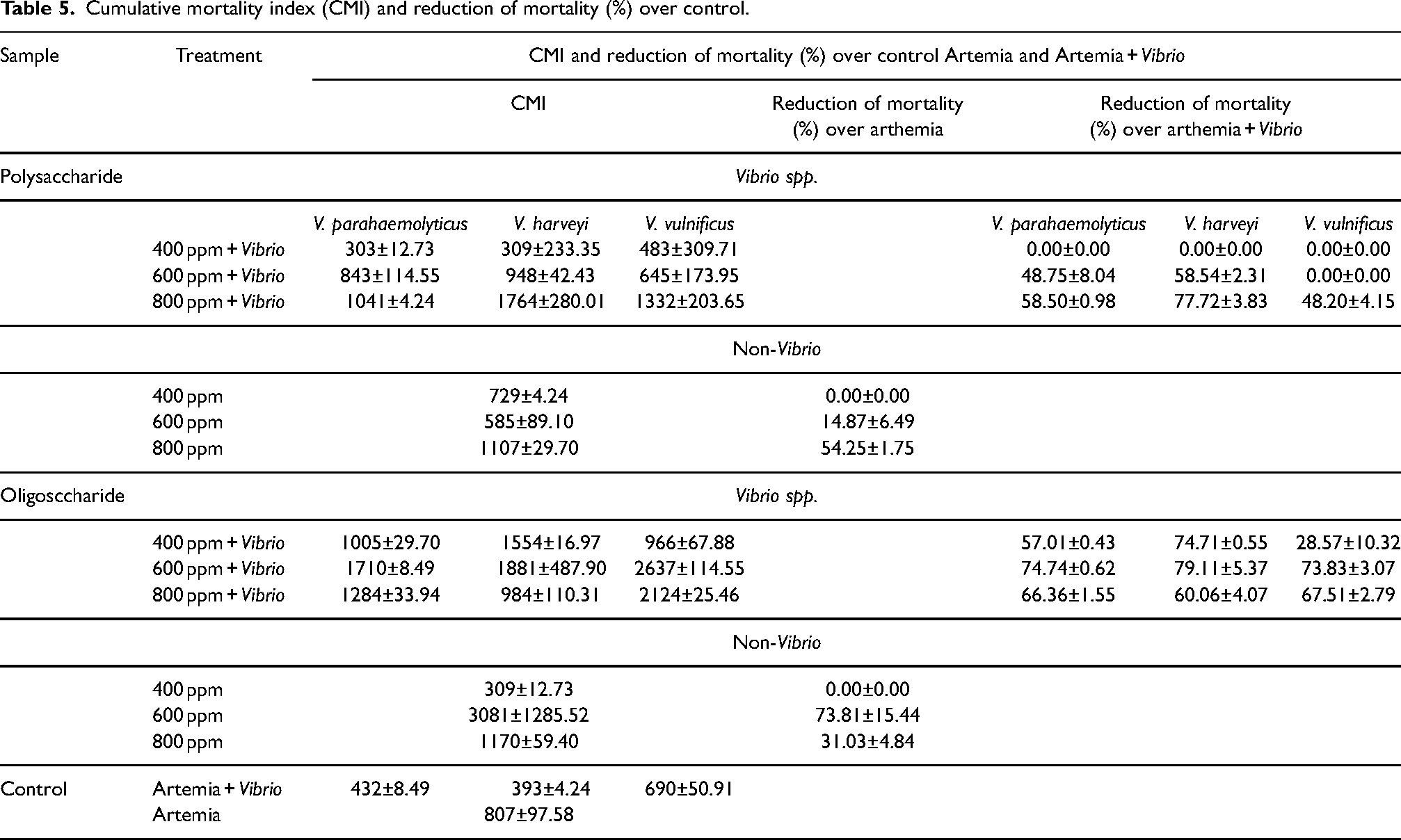
Based on Table 5, the unchallenged Artemia’s CMI was 807±97.58, the challenged Artemia was 432±8.49 (V. parahaemolyticus), 393±4.24 (V. harveyi), and 690±50.91 (V. vulnificus). The alginate-non-Vibrio treatment had varying CMI values and a reduction of mortality (%). The CMI of the polysaccharide was 303–1764 with a 0–77.72% reduction. The oligosaccharide was 966–2637 with a 28.57%–79.11% reduction. The highest CMI and reduction mortality was observed for polyssacharide treatments at 800 ppm concentration against V. harveyi (1764±280.01 with 77.72% ±3.83% reduction). Evidently, the oligosaccharide treatments gave different result. The highest CMI was from 600 ppm concentration against V. vulnificus (2637±114.55) with 79.11±5.37 reduction of mortality (%) against V. harveyi. The percentage reduction of mortality from oligosaccharide treatments was bigger and higher compared to the polysaccharides. Our data shows that all nine polysaccharides test exhibited a reduction in mortality, while only five reductions were observed for the oligosaccharides.
Cumulative mortality index (CMI) and reduction of mortality (%) over control.
Artemia is a well-known model for the study of resistance to pathogenic bacteria [45]. Some research concerning the antibacterial effect of caprylic acid in the culture system of Artemia franciscana nauplii inoculated with shrimp pathogens Vibrio harveyi and V. parahaemolyticus revealed that the mortality of Artemia nauplii reduced to 20.61 against V. parahaemolyticus and 16.30% against V. harveyi challenged groups [47]. High levels of antioxidant compound, such sodium ascorbate, was able to protect Artemia franciscana against Vibrio harveyi HSP70 infection [58].
The brominated furanones compound can interrupt the autoinducer 2 (AI-2) quorum sensing signal, and this was observed to control virulence from pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus to protect Artemia franciscana. The b-glucan compound managed to increase the survival rate of Artemia franciscana challenged with pathogenic Vibrio campbellii [60] and Vibrio harveyi [61].
Artemia can be applied as feed through encapsulation [57] to enhance the resistance of pathogenic bacteria [62]. Natural live feed Artemia franciscana encapsulated with 90% mannuronic acid polymer from brown algae Durvillaea antarctica acted as an immunomodulator of Halibut (Hippoglossus hippoglossus L.) larva challenged with Vibrio anguillarum. O2 succeeded in inducing immunostimulation and a 38% reduction in mortality was observed at high doses (55±1.4%). In addition, the encapsulated sulfated galactans in Artemia salina resulted in an improved the immune system of Litopenaeus vannamei shrimp [42].
Research from Gomez-Gil et al. [63] demonstrated that encapsulated Artemia franciscana with Vibrio parahaemolyticus and Vibrio alginolyticus have succeeded. This research had different data from different bacteria. In Vibrio parahaemolyticus, the quantity of encapsulated bacteria increased after 30 min and declined after 8 h. Finally, at 24 h, the bacteria level increased again but all nauplii were died. Some observations concerning Vibrio alginolyticus showed that the peak growth occurred at 45 min, the concentration of bacteria per nauplius decreased slowly to reach a minimum at 24 h, and there were no dead nauplii. In addition, Plant and LaPatra [64] confirmed that after removing the nauplii from the bacterial broth (at 30 min), bacterial concentration declined 10-fold at the next measurement (45 min). The reduction continued until it reached a minimum level at 8 h, with a slight increase observed at 24 h. This result can be beneficial in the application or oral administration of vaccines.
Artemia was successfully used for vaccine development approaches by encapsulating the pathogenic bacteria [65]. Researchers used Artemia encapsulated with a formalin-inactive culture of Vibrio anguillarum (109 cell mL–1) incubated for 1.5 hrs and implemented for Seabass’ larvae (Dicentrarchus labrax) oral feed. They concluded that the vaccine application method was safe and that an oral vaccine could be used to improve the food conversion ratio and growth performance. Similar reports from other groups also successfully used Artemia encapsulated with vaccine. These have applications for juvenile carp (Cyprinus carpio) and gilthead seabream (Sparus aurata) with Vibrio anguillarum. bioencapsulation. Zebrafish was fed with bioencapsulated Edwardsiella tarda phage (ETP-1) [66], and common carp (Cyprinus carpio var. Jian) larvae was fed with bioencapsulated recombinant Saccharomyces cerevisiae [67]. Moreover, some research has also combined Artemia with fenbendazole [45] in addition to erythromycin [68]. Encapsulation can also be applied to probiotic bacteria [5].
The alginate capsules contain a probiotic model of the Lactobacillus plantarum strain that serve as different surfactants. The treatment aimed to maintain survival during the original simulated gastric (SGF) and intestinal fluid (SIF) journey within 1 hour of exposure to SGF and two hours in SIF. This most likely related to their gradual penetration within the microcapsules, as shown by a confocal microscope, and their antimicrobial effects [69]. Other research by Akman et al. [70] developed and studied the effect of SA based probiotic Lactobacillus plantarum carrier edible films for the survival of probiotics by characterizing their physicochemical, mechanical, barrier, molecular, and thermal properties. Microencapsulation was applied to EPS as a potential method for protection of probiotics against adverse conditions of the gastrointestinal tract [71].
SA’s dry weight reached 4.21–6.47 grams with 21.05% –32.35% in yield. Polysaccharide and oligosaccharide alginate (AOS) samples had 8 and 12 with positive results, respectively. The highest alginate polysaccharides atibacterial activity (12.962±3.623 mm) was obtained from a challenge against V. harveyi, and the lowest was from V. vulnificus (6.550±0.676 mm). The highest AOS antibacterial activity (10.250±0.09 mm) was obtained from a challenge against V. harveyi, and the lowest one was from similar bacteria (6.670±0.81 mm). The polysaccharide and oligosaccharide alginate encapsulated against V. parahaemolyticus, V. harveyi, V. vulnificus and the non-challenged tests without any Vibrio spp. addition resulted in the best concentrations at 800 ppm for the polysacchride and 600 ppm for the oligosaccharide. Quantitatively, AOS were more effective than polysaccharides, and both alginates have potential antibacterial and immunostimulant properties against infection by V. parahaemolyticus, V. vulnificus and V. harveyi. Exclusively, AOS has the potential to be superior as an antibacterial agent and immunostimulant.
Footnotes
Author contributions
R.O.A., E.Y. and M.J.A. conceived and designed the project. N.A., W.S.A. and G.S. prepared the tests samples. All authors helped in analysing and interpreting the data. The paper was written with input from all authors and then they read and approved the final manuscript.
Funding
This research was partially funding by Ministry of Research and Technology, Indonesia via Superior Applied Research scheme (contract no. 184-64/UN7.6.1/PP/2021) and the Faculty of Fisheries and Marine Science, Diponegoro University through non APBN scheme with contract no: 75/UN7.5.10.2/PP/2021. So, therefore, the authors would like to give an enormous gratitude.
Sample availability
Samples and data used during the current study are available from the corresponding author.
Declaration of competing interest
The authors declares that they have no conflict of interest.
Acknowledgments
The authors would like to express their deepest thank to the Faculty of Fisheries and Marine Science at Diponegoro University for technical and support.