
Abstract
BACKGROUND:
Circular economy aims to get added value from byproducts of agriculture or livestock, including beekeeping. Classically in Alzheimer’s disease (AD), β-amyloid plaques and neurofibrillar clews formation have been considered as the main characteristics. However, the role of oxidative stress in the onset and progression of β-amyloid accumulation has been related.
OBJECTIVE:
The present study aims to test the antioxidant and neuroprotective effects of five hydrophilic extracts (sample 1–5; S1–S5) from a mud byproduct obtained from the recycling of beeswax using the Caenorhabditis elegans model.
METHODS:
The antioxidant activity was tested in vitro by DPPH, ABTS and FRAP assays. The content of phenols and flavonoids was evaluated by colorimetric procedures. Antioxidant and neuroprotective activities were investigated in vivo using the C. elegans model.
RESULTS:
The antioxidant capacity of the samples and their content in phenols and flavonoids was demonstrated. Nematodes treated with S1, S2 and S5 showed lower intracellular production of hydrogen peroxide. A neuroprotective activity of S1 extract was demonstrated by a delay in paralysis caused by the accumulation of beta amyloid (Aβ 1–42).
CONCLUSIONS:
S1 extract exhibits important antioxidant and neuroprotective activities in vivo, which could provide a novel source of natural antioxidants and neuroprotective compounds.
Introduction
Beekeeping is the maintenance of honey bee colonies, commonly in hives, by humans [1]. Bee products include honey and other products that the hive produces such as beeswax, propolis, pollen, poison, royal jelly and more. [2]. It is of great interest to study the various products that are obtained from the hive, like beewax [3], to add value to the beekeeping sector and, at the same time, contribute to the study of potential molecules of biomedical interest. The recycling of beeswax produces substances which are, until now, considered as industrial waste, however, they could be of significant value in the biomedical field [4–6]. Recently, it has been discovered that the flavonoid and phenol content in these by-products is very similar or higher than foods commonly considered to be rich in these compounds [4]. Among the components present in these by-products, quercitin derivatives, myricetin derivatives, kaempferol derivatives and other minor compounds, have shown antioxidant and neuroprotective effects [7–11].
Alzheimer’s disease (AD) is a multifactorial neurodegenerative disease of unknown etiology diagnosed for the first time by Alois Alzheimer in 1907, which show a progressive deterioration of cognitive function and memory, as well as dysfunction in language and orientation [12]. Around 44 million people are affected by AD around the world and it is estimated that by 2050 the number of people affected by AD will triple, with the consequent social and economic burdens for the state and their families [13]. Currently, the drugs developed for the treatment of AD are unsuccessful because they only show a slight improvement in symptoms, without actually preventing the progressive neurodegeneration. Therefore, it is urgent to develop more effective drugs to treat AD [14]. According to the amyloid cascade hypothesis, cerebral deposition of amyloid β-peptide (Aβ) is a typical feature of AD. The missense mutations in Amyloid Precursor Protein (APP), presenilin-1 (PS1) or presenilin-2 (PS2) genes increase Aβ production [15], causing Aβ-42 oligomerization and deposition and eventually harm functions of brain [16]. Classically, the formation of β-amyloid plaques and the formation of neurofibrillar clews have been considered as the only main characteristics that precipitate the symptomatic development of AD. However, it has been observed that, in addition to accumulations of β-amyloid and neurofibrillar clews, the brains of patients with AD usually have characteristics of a sustained inflammatory response together with higher levels of tissue oxidation and lower levels of antioxidant enzymes [17]. In this regard, healthy dietary patterns are characterized by high intake of plant-based foods, probiotics, antioxidants, soy beans, nuts, and omega-3 polyunsaturated fatty acids. Moreover, a low intake of saturated fats, animal-derived proteins and refined sugars are typical from these paterns. Overall, the intake of these foods have shown to decrease the risk of neurocognitive impairments and eventually the onset of AD [18]. Therefore, to observe the effects of bioactive compounds present in food against Aβ may be one of the important strategies for AD therapy. Caenorhabditis elegans is a small, transparent, free-living nematode that inhabits wet and warm soils. C. elegans has been developed as an alternative model to explore treatments for neurodegenerative diseases and study the mechanisms of gene expression, neurobiology and aging. The aim of this study was to evaluate the antioxidant capacity of different beeswax recycling by-products and the anti Aβ toxicity effects in C. elegans.
Methods
Sample collection and preparation
During the recycling process of the wax combs, the combs were collected and processed with steam. Sediment with organic and inorganic residues, including pollen, molts, etc., was separated from the wax. This sediment, called MUD 1, represented half of the initial weight of the processed product. Five samples of MUD 1: S1, S2, S3, S4 and S5, were randomly collected and used for hydrophilic extraction. Briefly, 1 g of each sample was diluted in 10 mL of distilled water and filtered through a 45 μm Minisart filter (PBI International). Subsequently, the extracts were dried by Speed vac (SC110 A, New York, USA). Finally, they were resuspended in a solution of KPO4 (1 M, pH 6) to incorporate it into the growth medium in the selected concentrations for each test.
Measurement of total phenolic and flavonoid content
The Folin–Ciocalteu method was used to determine the total phenol content (TPC) of the hydrophilic extracts, as reported by Singleton et al. [19], while flavonoid content (Flavo) of the extracts was determined using a colorimetric method described previously by Lin et al. [20].
Evaluation of the total antioxidant capacity (TAC)
For the determination of the TAC, the following analyses were carried out: the FRAP test, the DPPH free radical method and the TEAC test. For the FRAP test, the protocol described by Deighton et al. [21] was followed, which is based on the ability of the samples to reduce the ferric ion to ferrous. The absorbance measured at 593 nm reflects the magnitude of said reduction. For the DPPH method, the protocol described by Kumaran et al. [22] was followed, it is based on the reduction of DPPH by free radicals and their subsequent measurement through a 515 nm spectophotometric reading. Finally, for the TEAC test, the protocol described by Re et al. [23] was followed, based on the ability of the samples to deactivate the fluorescence of the ABTS radical, thus decreasing the absorbance values measured to 734 nm. All determinations were carried out in triplicate.
Strains and maintenance
C. elegans strains used in this work Including wild type N2 Bristol, CL2355 (smg-1 ts [snb-1/Aβ 1–42 long 3’-UTR]), CL4176 (dvls27 [marker gene mio-3/Aβ minigene + rol-6 (su1006)] X), JV1 [rpl–17p:: HyPer + unc-119 (+)], CL2122 (unc-54 vector + mtl-2:GFP) and CL802 (smg-1(cc546) I; rol-6(su1006) II) were obtained from the Caenorhabditis Genetics Center (Minneapolis, USA). CL2355 strain employs the promoter of the C. elegans synaptobrevin ortholog (snb-1) to facilitate neuronal expression of Alzheimer’s β-amyloid peptide in neurons, which is inducible upon the increase of the temperature from 16°C to 23°C. CL4176 expresses the β-amyloid peptide at the muscular level resulting in a paralysis phenotype which is also inducible by temperature upshift from 16 to 23°C. JV1 [rpl–17p:: HyPer+unc–119 (+)], ubiquitously presents the HyPer YFP sensor for hydrogen peroxide which is particularly useful for in vivo testing of ERO production.
N2 Bristol were routinely maintained in an incubator (VELP Scientifica FOC 120 E, Usmate, Italy) at 20°C, and the rest of the strains at 16°C, in solid nematode growth medium (NGM) with a lawn of Escherichia coli OP50 according to standard protocol. Age-synchronised animals were obtained by isolating embryos from gravid hermaphrodites using bleaching solution.
Lethality assay
To evaluate the toxicity of different extracts, N2 Bristol age-synchronised L4 worms were seeded in NGM without food with different concentrations of hydrophilic extracts. The concentrations used were the same for all extracts (0, 0.1, 1, 10, 50, 100 μg/mL) Then, they were incubated for 24 hours at 20°C before the counting of live and dead animals was carried out by visual inspection through a dissecting microscope (Motic Inc., LTD., Hong Kong, China) [24]. Death was considered when there was no response to mechanical stimulus generated with a platinum wire. At least three different experiments were carried out for each concentration with ten worms in each one. The results are expressed as the survival percentage (%). Lethality test results were used to choose the concentration to use in the following analyses.
Paralysis assay
This assay was carried out to determine the rate of paralysis of the transgenic strain CL4176 and the possible effects of hydrophilic extracts on this phenomenon were observed. The model proposed by Dostal et al. [25] with several modifications. In this assay, nematodes of the transgenic strain CL4176 in L3 stage were seeded in NGM with food and contained or not 10 μg/mL of the different hydrophilic extracts. After 48 hours of incubation at 16°C, the worms were transferred to an incubator at 23°C. After 20 hours, the count of paralysed worms was carried out every 2 hours until the last nematode was paralysed. Paralysation was assumed when there was no response to mechanical stimulus, generated with a platinum wire. At least three different experiments were carried out for each concentration with ten worms in each one. The results are expressed as non-paralysed worm percentage (%).
Chemotaxis assay
Age-synchronised transgenic C. elegans CL2355 were transferred onto NGM plates containing 10 μg/mL hydrophilic extracts or not, cultured at 16°C for 36 hours from L1 stage and moved to 23°C for another 36 hours to induce the expression of neuronal Aβ. Well-fed worms were collected at young adulthood (YA) stage by washing with K buffer (0.032 mol/L KCl, 0.051 mol/L NaCl) to remove the food. Subsequently, worms were conditioned or not for two hours in plates with benzaldehyde (1% in 100% ethanol). Finally, worms were dispensed on a plate without food containing on one side benzalheyde and on the other side ethanol together with sodium azide to facilitate its counting. After one hour, the number of worms located on both sides was counted, as the as described in He et al. [12]. Chemotactic index was calculated using the following formula: (worms paralysed in benzalheyde - worms paralysed in ethanol)/total worms.
Intracellular H2O2 production
This analysis was carried out to determine the effect of the different hydrophilic extracts on the production of intracellular H2O2 in young nematodes of the transgenic strain JV1. In this trial, the protocol described by Back et al. was reproduced with some modifications [26]. In summary, synchronised embryos were seeded throughout life in plates with NGM and food that included or not 10 μg/mL of the extracts. After 72 hours of culture, the nematodes of each extract were divided in two groups (induced and basal) and were washed in a solution of M9 and EDTA independently to remove the remains of E. coli. To facilitate the expression of the H2O2 fluorescent sensor of strain JV1, an induction with 20 mM H2O2 was carried out in one of groups of each extract. Subsequently, a 15-minute reading was performed on a spectrophotometer (Synergy 2 Biotek; Bad Friedrichshall, Germany) at a wavelength of 490 and 405 nm, respectively. The results were expressed as the average of the 490/405 ratio of each group and were normalized by the number of nematodes.
Statistical analysis
Unless otherwise indicated, the results expressed show the values of the mean±the standard error of the mean for a minimum of 3 replicates, or in the case of lethality analysis, for a minimum of 25 worms per treatment. To verify the existence of significant differences between two groups, the statistical tests used were the Student’s t test for chemotaxis and paralysis tests and the Kruskal-Wallis test for the intracellular hydrogen peroxide production test. Statistically significant differences have been considered for a P < 0.05. For statistical analysis, SPSS 24.0 software was used.
Results
Composition of by-products from beeswax recycling process
In this work the total antioxidant capacity, the total flavonoid and the polyphenol content of each individual extract were evaluated. However, the results shown in Table 1 refer to the average of the data obtained from the five extracts. The data shows that the average total phenolic content of the five extracts is 9.77 mM (expressed in mM gallic acid). On the other hand, the total flavonoid content is 3.34 mg/mL (expressed in mg/mL of catechin). With respect to the techniques that analyze the antioxidant capacity, ABTS obtained 1.6 mM (Expressed in mM of trolox), FRAP obtained 1929.78 μM (expressed in μM of trolox) and, finally, DPPH obtained 939.84 μM (expressed in μM of trolox).
Total phenolic and flavonoid contents and total antioxidant capacity of the average of the five samples
Total phenolic and flavonoid contents and total antioxidant capacity of the average of the five samples
Results are mean±SEM. AS shows the average of the five samples.
Lethality
First of all, we evaluated the short-term toxicity in the wild strain N2. As shown in Table 2, none of the extracts produced short-term fatal effects on the nematode at any of the concentrations tested. The concentration of 10 μg/mL was selected as the standard concentration for the rest of the experiments because any of the assayed concentrations caused lethality, because the amount of MUDs was not so high and because 10 was the intermediate concentration.
Percentage of live worms after the treatment with different concentrations (0.1, 1, 10, 100 and 1000 μg/mL) of differents samples for 24 h
Percentage of live worms after the treatment with different concentrations (0.1, 1, 10, 100 and 1000 μg/mL) of differents samples for 24 h
Results are mean±SEM. S1–S5 indicates groups exposed to different extracts.
Intracellular ROS production assay was performed to evaluate the protection of different hydrophilic extracts in C. elegans against intracellular hydrogen peroxide production. JV1 mutant strain of C. elegans has a biosensor sensitive to redox changes in vivo. We used a free radical generator (20 mM H2O2) that results in oxidative stress, thereby investigating the antioxidative effects of the the differents hydrophilics extracts in JV1. As shown in Fig. 1, worms treated with 10 μg/mL of the extracts S1 and S5 during their whole lives had less intracellular production of ROS than the untreated worms at baseline. This effect was maintained when the worms were exposed to 20 mM hydrogen peroxide. S2 could only reduce intracellular peroxide production in the exposed group. No significant differences were found for the other extracts tested. However, significant differences were found when the individual effects of the extracts were grouped (AS) in the exposed group.
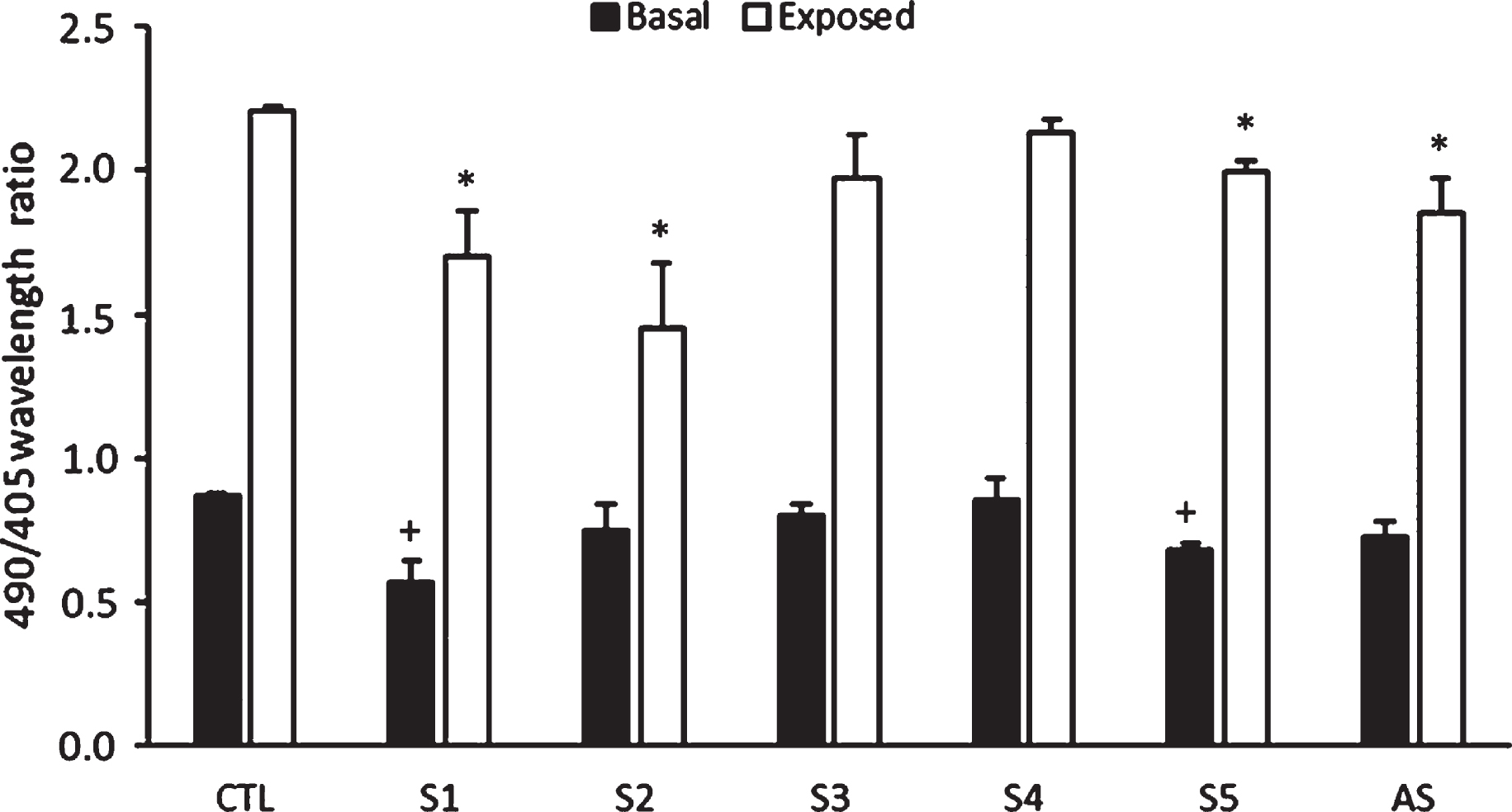
Effects of the samples tested on intracellular hydrogen peroxide production. Treatment with S1 and S5 reduced the intracellular production of hydrogen peroxide in nematodes that were exposed or not to 20 mM of hydrogen peroxide. Treatmen with S2 only reduced the intracellular production of hydrogen peroxide in nematodes that were exposed to 20 mM of hydrogen peroxide. S1–S5 indicate groups treated with different extracts. AS show the average of the effects of the five extracts on the intracellular production of hydrogen peroxide. (+) and (*) mean significant differences respect to basal and exposed control (CTL), respectively. P < 0.05.
The rapid paralysis from Aß expression in the transgenic C. elegans strain is useful for investigating and developing drug effects. CL4176 has a phenotype of Aβ production and aggregation when grown at 23°C, and progressive paralysis subsequently was observed [27]. To identify protective small molecule compounds against Aβ toxicity, we examined the effects differents hydrophylic extracts from by-products beewax recycling on the CL4176 strain. As shown in Fig. 2A, almost complete paralysis in all groups was found within 37 h at 23°C; however, as can be seen at Fig. 2B, the Aβ-induced paralysis rate was significantly delayed in C. elegans CL4176 strain with S1 treatment until 27 h at 23°C. No significant differences were found for the other extracts tested.

Effect of the different extracts tested on the paralysis phenotype in the transgenic strain CL4176. (A) Paralysis timeline of treated and untreated nematodes. (B) Paralysis assay at 27h after the temperature increase. S1 treatment presents less number of paralyzed worms with respect to untreated nematodes at 27 h. S1–S5 indicate groups treated with different extracts. AS show the average of the effects of the five extracts on the paralysis phenotype. * Indicate significant differences with respect to control (CTL). P < 0.05.
The dysfunction in associative learning memory is an AD-like symptom induced by Aβ toxicity. In this case, CL2355 strain employs the promoter of the C. elegans synaptobrevin ortholog (snb-1) to facilitate neuronal expression of Alzheimer’s β-amyloid peptide in neurons. To evaluate the chemotaxis index associated with learning in C. elegans, a 2 hour conditioning process was set before the chemotaxis assay to pair the repulsive situation of food deprivation with attractive situation of the presence of benzaldehyde. As shown in Table 3, the reduction of chemotaxis index between non-conditioned and conditioned worms suggested that the worms without Aβ toxicity successfully associated the benzaldehyde with starvation. Nonetheless, when comparing groups, the reduction in the chemotaxis index was not significant in any of the treated groups, conditioned or not.
Effects of the samples tested on the chemotactic index
Effects of the samples tested on the chemotactic index
Results are mean±SEM. CTL: 2355 control group; CL2122: control strain. S1–S5 indicates groups treated with different extracts. AS show the average of the effects of the five extracts on chemotactic index.
To the best of our knowledge, this is the first study that evaluates the protective effects of by-products from beeswax recycling process against Aβ toxicity in C. elegans. To understand the regulatory mechanisms of oxidative stress and anti-Aβ disposition of our extracts, we performed experiments using C. elegans. We observed that S1 and S5 provided preventive effects for C. elegans intracellular ROS production induced by H2O2 as well as S1 protection against paralysis in C. elegans induced by temperature-up-shifted.
It should be noted that the effects of extracts obtained during the recycling of beeswax on Aβ toxicity have not been evaluated in the nematode. However, there are numerous in vitro and rodent studies that could explain the effects shown through the use of its isolated components. Previously, Giampieri et al. [4] characterized the different extracts. S1 stood out for its high content of quercetin derivatives (quercetin-3-O-rutinoside and quercetin-3-O-rhamnoside) and myricetin derivatives (myricetin-3-O-rutinoside). The mechanisms of the beneficial effects of S1 could be explained through isolated compounds. Quercetin suppresses β-amyloid synthesis in cell-free, cell-based and in silico studies by inhibition of BACE1-mediated cleavage of APP [28]. In addition, Hirohata et al. [29] reported that myricetin, another of the predominant compounds present in S1, exerted anti-amyloidogenic effects in vitro by preferentially binding to Aβ through its hydroxyl arms. In vitro tests conducted by Jagota et al. [30], showed that treatment with 50 mM quercetin decreased aggregation and formation of Aβ oligomers, which have been shown to be more toxic than fibrils [31]. In this context, Regitz et al. [32] observed that exposure to different concentrations of quercetin (10 μM and 100 μM) reduced the paralysis phenotype in the transgenic strain CL2006 of C. elegans due to an activation of proteosomal and macroautophageal degradation, which translates into a reduction in the amount of oligomeric aggregate β-A1–42 and a reduction in paralysis [33]. In addition, quercetin and its metabolites have been reported to act in several signaling pathways, including cAMP response element binding protein (CREB), c-Jun N-terminal kinases, mitogen-activated protein, macroautophagy, calcium homeostasis, proteasomal degradation and GADD34-eIF2α-ATF4, pathways that participate in AD pathogenesis. In addition, quercetin serves as an Sirt1 agonist and acetylcholine esterase inhibitor (AchE) to improve AD phenotypes [7, 34]. Also relevant are some recent findings that show that quercetin improves the pathology of Alzheimer’s disease and related cognitive deficits in a mouse model with triple transgenic Alzheimer’s disease [35].
On the other hand, oxidative stress is recognized as an important factor in a variety of neurodegenerative diseases, as a mediator of the adverse effects of a series of neurotoxins and as a mechanism for age-related degenerative processes. In vitro studies in neuronal cell lines and in primary neurons have shown that quercetin, at micromolar concentrations, antagonizes cellular toxicity induced by various oxidants (e.g., hydrogen peroxide, linoleic acid hydroperoxide) [36]. In accordance with the above, Thabit et al. [37] observed that treatment with Cassia fistula, whose components include quercetin, was able to reduce oxidative stress through upward regulation of SOD-3 expression and upward regulation of HSP-16.2 chaperone protein, which has been found to partially suppress Aβ-induced paralysis by modulating oligomerization of Aβ in C. elegans. This could explain the protective effect of S1 extract on the intracellular production of ROS and its protective effect against the paralysis phenotype. However, not all molecules with antioxidant capacity have direct neuroprotective effects. For example, in our study, S2 and S5 reduced intracellular levels of hydrogen peroxide, but did not exhibit effects on the paralysis phenotype. Similarly, in the study conducted by Wu et al. [38], they observed that the administration of L-ascorbic acid reduced intracellular levels of hydrogen peroxide, but was not effective in suppressing paralysis in the transgenic strain CL4176 of C. elegans. However, the dissociation of the antioxidant effect to delay paralysis does not rule out the contribution of oxidative damage in abnormal Aβ-dependent behaviors.
Finally, Wu et al. [38] observed that the chemotactic behavior does not correlate with the average density of the Aβ oligomers. It is possible that chemosensory neuronal circuits are less sensitive to Aβ toxicity and, therefore, chemotactic behavior may not be the best test to assess amyloid toxicity. However, the paralysis assay correlates well with the density of Aβ oligomers, which could support our hypothesis that S1 and its components alleviate behavioral abnormalities by decreasing levels of toxic Aβ oligomers.
Conclusions
In conclusion, our study demonstrates the potential antioxidant and neuroprotective activities of the S1 extract obtained during the recycling of beeswax in C. elegans. The extract was able to reduce intracellular levels of hydrogen peroxide at baseline as well as when it was exposed to exogenous hydrogen peroxide. In addition, the neuroprotective effects of the extract were shown by a decrease in the paralysis phenotype. Further studies are required to study the exact molecular mechanism by which the extract exerts these effects, and to prove its efficacy and safety in higher model organisms.
Funding
The authors report no funding.
Conflict of interest
The authors have no conflict of interest to report.
Footnotes
Acknowledgments
Jose M. Romero-Márquez and María D. Navarro-Hortal are FPU fellows from the Spanish ‘Ministerio de Educación y Formación Profesional’. The present study was partially funded by the “National Beekeeping Aid Program (Programa Nacional de Ayudas a la Apicultura)”, cofounded by the European Union and assigned to Spanish FEGA and FEAGA agencies (2016).