
Abstract
Diabetes mellitus (DM) is a chronic metabolic disorder and one of the most challenging health problems worldwide. Left untreated, it may progress causing serious complications. Genetics, epigenetics, and environmental factors are known to play an overlapping role in the pathogenesis of DM. Growing evidence suggests the hypothesis that the environment induces changes in the early phases of growth and development, influencing health and disease in the adulthood through the alteration in genetic expression of an individual, at least in part. DNA methylation, histone modifications and miRNAs are three mechanisms responsible for epigenetic alterations. The daily diet contains a number of secondary metabolites, with polyphenols being highest in abundance, which contribute to overall health and may prevent or delay the onset of many chronic diseases. Polyphenols have the ability to alter metabolic and signaling pathways at various levels, such as gene expression, epigenetic regulation, protein expression and enzyme activity. The potential efficacy of polyphenolic compounds on glucose homeostasis has been evidenced from in vitro, in vivo and clinical studies. The present review is designed to focus on epigenetic regulation exerted by polyphenolic compounds in DM and their complications, as well as to summarize clinical trials involving polyphenols in DM.
Keywords
Introduction
Diabetes mellitus (DM) is a chronic metabolic disorder characterized by hyperglycemia that results from defects in insulin secretion, insulin sensitivity or both. It is one of the most challenging health problems worldwide, and evidence suggests that it is epidemic in most developing countries [1]. According to a report published by the World Health Organization (WHO) in 2016, about 422 million people were living with DM in 2014 around the globe. Furthermore, a slight increase in the annual mortality rate of DM has been seen in 2016 (1.6 million) when compared to 2012 (1.5 million) [2]. This increases the economic burden of individuals on the healthcare system. The International Diabetes Federation (IDF) reported the direct annual cost of diabetes for people in the age range of 20 to 79 years to be approximately 153 billion dollars internationally [1].
The most common mechanisms thought to cause DM include insulin resistance, associated with numerous factors (visceral obesity, oxidative stress and chronic inflammation), and decreases in insulin secretion associated with β cell destruction. Impairment in insulin action and secretion may occur concomitantly in most cases, though the mechanism responsible for hyperglycemia is yet to be entirely understood [3–5]. Different factors are known to play an overlapping role in the pathogenesis of DM such as genetics (PPAR-γ, KCJN11, HNF4A, ENPP1, GCKR, IRS1, PCSK1 and G6PC2), epigenetics (a molecular connection between environmental factors and DM) and environmental factors (including age, sedentary lifestyle, obesity, unhealthy diet and certain medications) [6–8]. When left untreated, DM can progress to serious complications which may affect patient’s quality of life and reduce life expectancy. Acute complications, referred to as diabetic emergencies, develop more suddenly and include diabetic ketoacidosis (DKA) and hyperosmolar hyperglycaemic state (HSS), while chronic complications develop gradually over time and include nephropathy, retinopathy, peripheral neuropathy, autonomic neuropathy, peripheral arterial disease (PAD), atherosclerotic cardiovascular disease and cerebrovascular events [9–11].
Recent decades have borne tremendous improvement, with the development and approval of new agents for the treatment of DM and associated complications, focusing on insulin resistance and insulin deficiency [12]. The foremost issue currently faced by physicians is the high cost of available therapeutic options, as it increases economical burden on patients and may affect adherence to the therapy [13]. The incidence of DM is also continuously rising among very young individuals, but the available therapeutic options for this cohort are limited. Only two anti-diabetic agents, insulin and metformin, are currently approved by the US Food and Drug Administration (FDA) for use in children and youths [14, 15]. Plant extracts and isolated compounds can be a source of alternative treatments in such cases, as supported by the traditional usage of plant extracts for the treatment of DM [16, 17]. Modern literature also supports the benefits of plant extracts as a whole or as isolated components against diabetes and other metabolic abnormalities, as evidenced in experimental studies [18].
Polyphenols are the largest group of plant secondary metabolites, they contain a benzene ring with hydroxyl (OH) moieties and range from simple flavonoids and phenolic acids to structurally complex procyanidins. Extensive research has been carried out on this group of compounds to explore their therapeutic potential [19–25]. They are the most abundant phytochemicals found in the human diet, possessing many capacities such as cardioprotective, anti-diabetic, antiasthma, antimicrobials, anxiolytic, antidepressant, anticarcinogenic, neuroprotective, to name a few. Their antioxidant properties make them valuable agents in the prevention and treatment of age related diseases [26]. Table 1 presents the main classes of polyphenols with their basic structures and respective examples. The objective of the present paper is to review the research progress on the significant role of polyphenols in the prevention of diabetes and related complications through modulation of epigenetic pathways.
Main classes and examples of polyphenols [165]
Main classes and examples of polyphenols [165]
The word epigenetic is of Greek origin, which means above the genes and was coined by Conrad Hal Waddington, who defined it as “the causal interactions between genes and their products, which bring the phenotype into being” [27]. Epigenetic mechanisms are vital for regulating gene transcription and genomic stability, without making changes to the underlining DNA sequence. Later on Russo et al. (1996) defined epigenetics as “mitotically and/or meiotically heritable changes in the gene function, that cannot be explained by change in the DNA sequence” [28]. In other words, epigenetics deals with the change in DNA expression without being an alteration in the structure of the DNA.
After extensive research in the field of epigenetics, a hypothesis on the developmental origins of health and disease was proposed, stating that the environment induces changes in the early phases of growth and development (from conception to early childhood), with long term effects on later health and disease. It is believed that these changes may be caused by alteration in the genetic expression of an individual [29]. Scientists have now identified several biomarkers related to the alteration of genetic expression, associated with a number of chronic pathologies like autoimmune and inflammatory diseases, cancer, neuropsychiatric disorders, diabetes, obesity, and cardiovascular disorders. These biomarkers could serve as therapeutic targets [30–35]. The mechanisms that serve their role under the umbrella of epigenetics include DNA methylation, histone modification and miRNAs, and these mechanisms provide means for the organism to react upon environmental cues [36].
DNA methylation
DNA methylation is a covalent modification of DNA, catalyzed by DNA methyltransferases (DNMTs), which involves the transfer of a methyl group from S-adenosyl-L-methionine (SAM) to position 5 of cytosine (5’ cytosine), followed by guanine residues (CpG sequences). DNA methylation is associated with gene silencing, genomic imprinting, X-chromosome inactivation and regulation of transcription events [37–39]. Properly regulated DNA methylation is essential for maintaining genome stability and transcriptional regulation of nearby genes [37]. Three members of the DNMT family are well-known in vertebrates for their role in DNA methylation: DNMT1, DNMT3A and DNMT3B. DNMT1, also known as maintenance methylase, maintains the DNA methylation process as it copies the pre-existing methylation mark from parent to daughter strands during replication, however literature also support the function of DNMT1 as de novo DNA methyltransferase. DNMT3A and DNMT3B are responsible for de novo methylation of previously unmethylated CpG sequences, which normally occurs in early embryonic stem cells or cancer cells [40]. Alterations in DNA methylation (hypo- or hyper-methylation) have been linked to numerous chronic disorders like cancer, imprinting disorders (neurodevelopmental and neurobehavioral disorders), aging and atherosclerotic disorders [41]. FDA has approved DNMT inhibitors for the treatment of several chronic disorders due to their effectiveness [42].
Histone modifications
Histones are basic proteins that reduce the length of DNA by packing and ordering the structural units called nucleosomes. The structure of nucleosome involves eight histone proteins, two pairs each of H2A, H2B, H3 and H4, around which DNA is supercoiled [43]. Histones are subjected to posttranslational modifications (acetylation, phosphorylation, ubiquitination, and sumoylation) which can affect gene expression by altering chromatin structure, which modulates the access of nuclear proteins to specific sections of DNA [44, 45]. Important and well-known regulators of histone modifications are histone acetyltransferase (HATs), histone methyltransferase (HMTs), histone deacetylases (HDATs) and histone demethylases (HDMTs). Newly synthesized histones get acetylated by HATs through the transfer of negatively charged acetyl groups from acetyl coenzyme A (acetyl Co-A) to positively charged lysine residues, resulting in weakening the interaction between DNA and histones which may be associated with an increase in transcriptional activity. HDACs remove an acetyl group from lysine residues, transferring it back to acetyl Co-A, resulting in inactivation of transcriptional activity. HMTs perform methylation of lysine and arginine residues by transferring a methyl group from SAM, which may result in transcriptional activation or repression depending on the degree of methylation and methylated residues. HDMTs counteract the action of HMTs by demethylating histone residues [46–48]. SIRT1 is a member of sirtuins superfamily of HDACs, which deacetylates many transcription factors such as NF-κB, p53 and histone proteins and thus may regulate cell survival, metabolism, inflammation, aging and immune function [49].
Micro RNAs
Micro RNAs (miRNAs) are small, 21–25 nucleotides long, non-coding RNAs, mainly involved in post-transcriptional silencing and messenger RNA (mRNA) decay [50]. Biogenesis of miRNA starts in the nucleus with synthesis of primary miRNA by the enzymatic action of RNA polymerase, which is further cleaved by the microprocessor complex and transported to cytoplasm where it is processed by a dicer to produce mature miRNA [51, 52]. Lin-4 was the first member of the miRNA family to be discovered, in Caenorhabditis elegans in 1993 by Lee and colleagues, which led scientists to identify several other members of this family, and now more than 2500 miRNAs have been discovered in humans to date [53–56]. MiRNAs are known to regulate over 60% of protein encoding genes across the entire human genome. They are supposed to bind target mRNA, causing its degradation or inhibiting its translation [57–59]. Furthermore, a single miRNA can regulate more than one mRNA as they do not required perfect complementarity for target recognition. However in some cases a single mRNA may also be regulated by multiple miRNAs [60]. The 5' region of miRNAs is associated with target binding, by base pairing to sequence motifs in the 3' untranslated region (UTR) of mRNA [61]. Altered expression of miRNAs can be associated with the pathogenesis of numerous chronic diseases, including cancer, metabolic disorders, myocardial infection and hepatitis C. The over-expression or under-expression of miRNAs in particular diseases gives rise to a signature miRNA pattern [62]. The particular role of miRNAs in diabetes remains unclear, though it can be used as biomarker for disease diagnosis and prognosis. They may be related to the fate of pancreatic β-cells, insulin secretion and insulin sensitivity. Moreover a cluster of miRNAs could be involved in DM. rather than single miRNA [63]. MiRNAs could be a potential target by polyphenols in disease management as demonstrated by Devi et al. (2017) [52].
Epigenetic regulation of diabetes and complications by polyphenols
Nutritional genomics deals with the interaction between bioactive food components and the genome, and includes studies of nutrigenetics, nutrigenomics and epigenetic modifications caused by food components [64]. The daily diet contains a number of secondary metabolites, with polyphenols being the most abundant, that contribute to overreach health and may prevent number of chronic diseases [65]. Since DM is a multifactorial disease, it is imperative to find therapeutic options that can target multiple mechanisms. Polyphenols have the ability to alter metabolic and signaling pathways at various levels such as gene expression, epigenetic regulation, protein expression and enzyme activity [66]. They can modulate glucose metabolism, inhibition of α-amylases or α- glucosidases, improve insulin resistance, attenuate hyperglycemia and dyslipidemia, decrease oxidative and inflammatory stress markers and improve adipose tissue metabolism, the gut microbiota and inhibit advanced glycation end products [64, 67–69]. The effects of polyphenols on carbohydrate metabolism and glucose homeostasis in DM have been summarized in Fig. 1. Table 2 has listed the epigenetic targets of polyphenols in relation to diabetes or diabetic complications.
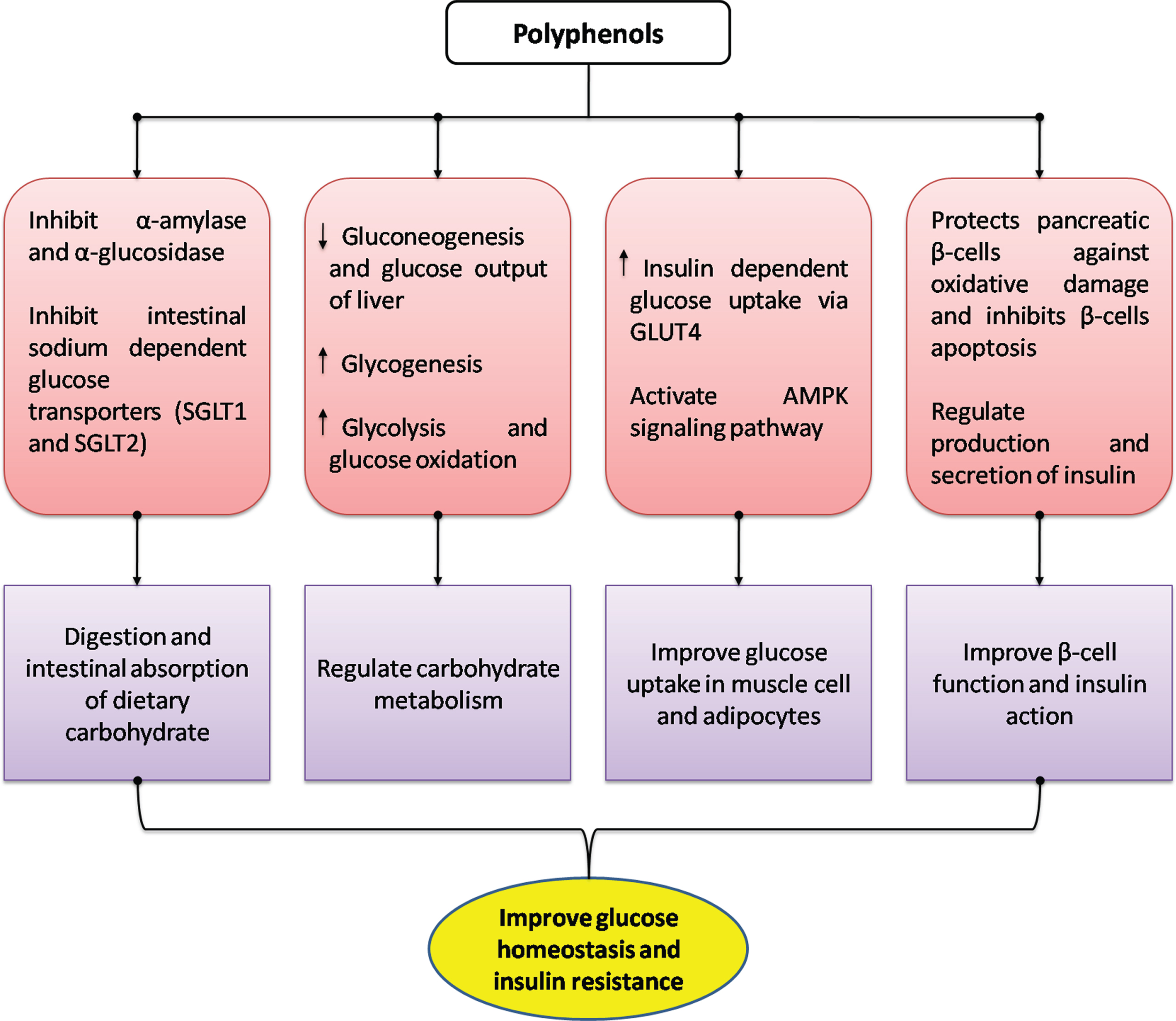
Effects of polyphenols on glucose homeostasis in management of diabetes mellitus. Modified from Bahadoran et al. (2013) [67].
Epigenetic targets of polyphenols in relation to diabetes or complications
Epigallocatechin gallate (EGCG), histone deacetylase (HDAC), DNA methyltransferase (DNMT), micro RNA (miRNA), histone acetyl transferase (HAT), sirtuin (SIRT).
Epigallocatechin gallate (EGCG) (Fig. 2a) is the most abundant catechin present in green tea. Chemically it is a flavan-3-ol phenolic compound containing eight free hydroxyl groups, which make it a bioactive compound with a wide range of pharmacological effects, including diabetes, metabolic syndrome, cardiovascular disorders, cerebral ischemic stroke and many more [70]. Studies have shown that EGCG may regulate gene expression and molecular signaling pathways [71]. Consumption of green tea can increase insulin sensitivity, improve insulin resistance, regulate insulin signaling pathways, protect islet β-cells, and ameliorate oxidative and inflammatory stresses [72]. Some of these effects are mediated by epigenetic regulations by EGCG, as it inhibits HDAC activity and HDAC2 expression in regulatory T cells (Treg) and decreases nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) [73]. A gallate derivative methyl-3-O-methyl gallate may protect against oxidative damage in chronic disorders by reduction of miR-17–3p expression [74].
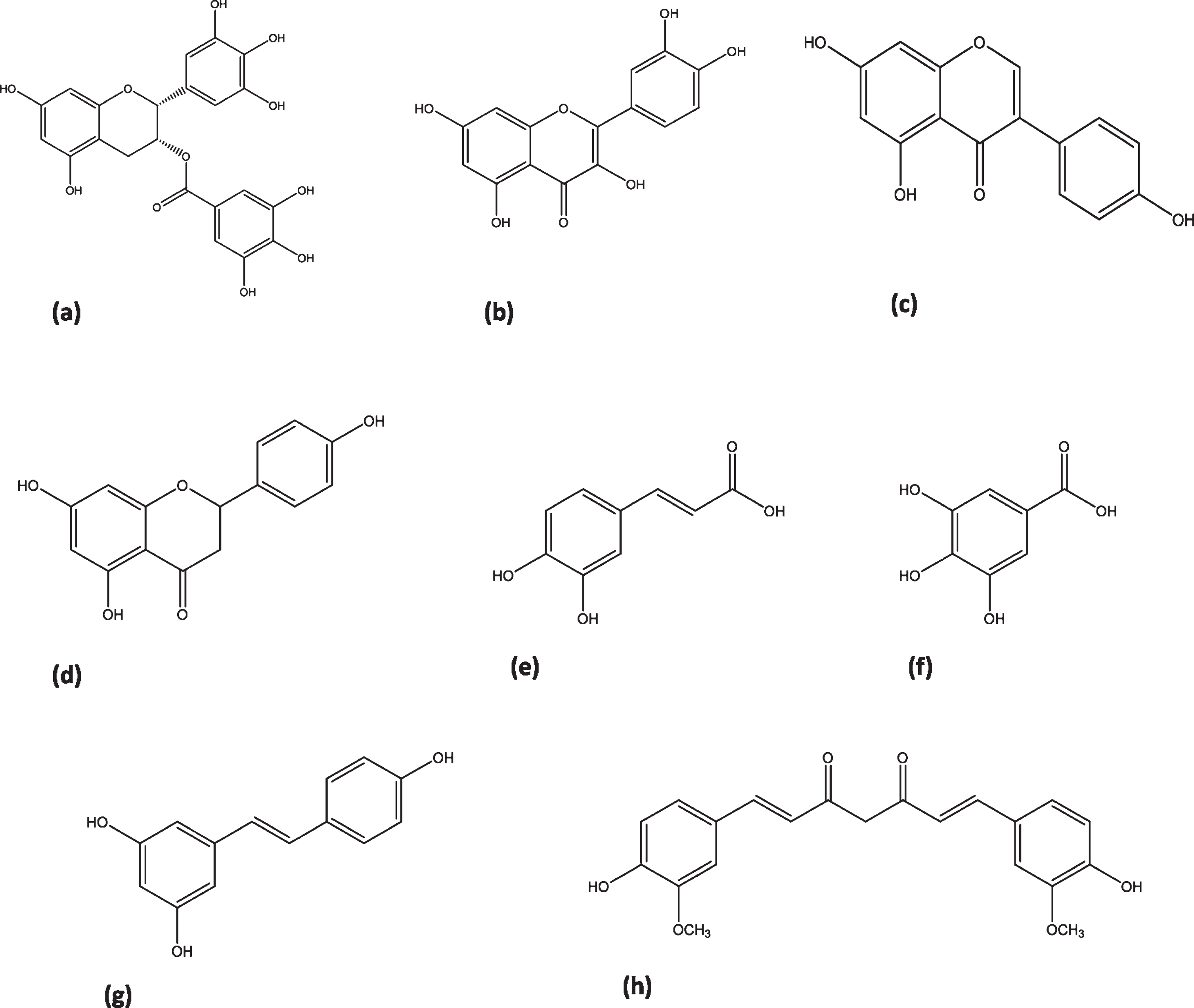
Chemical structures of natural polyphenols. (a) EGCG, (b) quercetin, (c) genistein, (d) naringenin, (e) caffeic acid, (f) gallic acid, (g) resveratrol, (h) curcumin.
An in vivo study on C57BL/6J male mice showed the inhibition of enzymatic DNA methylation via DNMT mediated DNA methylation through increased formation of S-Adenosyl-L-homocysteine (SAH), a potent inhibitor of SAM mediated reactions, and through alteration of the availability of methyl groups which leads to prevention of high-fat diet induced changes, when treated with EGCG in daily dose of 25 mg/kg body weight for 4 months [75]. It has been shown to alleviate maternal diabetes induced neural tube defects in diabetic pregnant mice by inhibition of DNA hypermethylation, treated with 10μM, while no such effects were seen at dose of 1μM [76]. Inhibition of DNMT by EGCG may be associated with decreasing reactive oxygen species (ROS), reducing glucose levels and enhancing glucose tolerance in animals [77]. Baselga-Escudero et al. demonstrated from in vitro study, that EGCG could regulate miRNA-33a and miRNA-122, where both of these miRNAs are related to the development of DM and metabolic syndrome [78]. Furthermore EGCG reduced the expression of prostaglandin E2 (PGE-2) and cyclooxygenase-2 (COX-2) through stimulating hsa-miRNA-199a-3p in interleukin (IL)-1b-stimulated human OA chondrocytes [79].
Quercetin (Fig. 2b) belongs to the flavonols, a chemical class of flavonoids, and is a potent antioxidant of plant origin. It exerts many different pharmacological effects including antioxidant, anti-infective, anticancer, anti-inflammatory, hepatoprotective, cardio-protective, anti-diabetic, anti-obesity and neurological effects [80, 81]. The estimated average intake of quercetin is 10 –16 mg/day, while the recommended intake of quercetin aglycone as a dietary supplement is 1 gram per day [82, 83]. It possesses anti-diabetic effects, mainly by stimulating glucose uptake through an mitogen-activated protein kinase (MAPK) insulin dependent mechanism [84]. It also improves renal functioning in diabetic nephropathic rats by blocking the expression of connective tissue growth factor (CTGF) and transforming growth factor-β1 (TGF-β1) [85].
Moreover in vitro studies have revealed the effective inhibition of aldose reductase and prevention of polyol accumulation by quercetin [86]. It helps to decrease the seriousness of the numbness, jolting pain and irritation of diabetic neuropathic patients [87]. Quercetin is a natural epigenetic regulator, as it inhibits DNA methylation by DNMTs, decreases expression of DNMT1, demethylates gene promoter regions, suppresses HAT activity and inhibits HDACs [88]. It has been evidenced that a sucrose solution containing quercetin and resveratrol attenuated metabolic dysfunction in male Wistar rats by altering the expression of histone deacetylates SIRT1 and SIRT2 in white adipose tissues, when supplemented with resveratrol (10 mg/kg/day) plus quercetin (0.19 mg/kg/day) or resveratrol (50 mg/kg/day) plus quercetin (0.95 mg/kg/day) daily for 4 weeks [89]. Quercetin reduced DNMT activity and DNA methylation of the nuclear factor erythroid-2-related factor-2 (Nrf2) promoter in male ICR mice treated with 40 mg/kg/day or 80 mg/kg/day quercetin for 20 days, leading to protection against inflammatory response [90].
Genistein
Genistein (Fig. 2c) is an isoflavone compound well-known for its estrogenic effects. It is present in soy based foods. It exhibits multiple therapeutic benefits ranging from cardiovascular protective activity [91, 92] to anticancer activity [93] through its antioxidant, antiproliferative and proapoptotic effects [94, 95]. Genistein intake is beneficial in DM and other metabolic diseases, more probably though increasing insulin secretion, alleviating insulin resistance, and reduction of low grade inflammation, ROS and adipocyte tissue mass [96, 97]. Soy foods are thought to exert beneficial effects during embryogenesis due to their regulation of DNA methylation and histone modification. Thus, increasing evidence suggests that it possesses long term effects on health and disease in childhood [98, 99].
Genistein may alter the epigenome of offspring in utero, and a report has shown that genistein induced DNA hypermethylation is likely to persist in adulthood, leading to alteration of coat color distribution and protection against metabolic diseases [100]. A study using nonhuman primates demonstrated that genistein improved body weight, insulin sensitivity and lipid profiles, and altered DNA methylation in the liver and muscles [101]. Although genestein influences histone acetylation and methylation in various cancer cell lines, data does not exist in diabetic models [102].
Naringenin
Naringenin (Fig. 2d) belongs to the flavanone class of flavonoids, widely distributed in citrus fruits such as oranges and grapefruits. It has received a considerable attention in the therapeutics of human disorders, with special attention on antioxidant, anti-inflammatory, antibacterial, antiviral, anticancer, antiadipogenic and cardioprotective effects [103, 104]. Naringenin possesses the potential to prevent against metabolic diseases, via numerous mechanisms including modulation of antioxidant enzymes, phosphoinositide 3-kinase/protein Kinase B/nuclear factor (erythroid-derived 2)-like 2 (PI3K/Akt/ Nrf2), NF-κB, nuclear factor (erythroid-derived 2)-like 2/antioxidant responsive element (NRf2/ARE), MAPK, 3-hydroxy-3- methyl-glutaryl-coenzyme A (HMG-CoA) reductase, PPAR, and nitric oxide-cGMP-protein kinase G-induced KATP channel (NO-cGMP-PKG-KATP) [105].
In DM, it has been reported to improve serum glucose and insulin levels, alleviating oxidative and inflammatory stresses, improves insulin growth factor (IGF), restoring diabetes related renal and hepatic damage as well as improving nerve growth factor (NGF) expression of the sciatic nerve, and sciatic nerve histology [106–108]. Naringenin is a potent DNMT inhibitor, and at nutritionally relevant doses it may alter the expression of hundreds of genes in the liver, aorta or lungs [109, 110]. It has been shown to modulate gene expression involved in glycolysis and gluconeogenesis in the liver [111, 112].
Studies have linked the antidiabetic effects of naringenin with the activation of HDAC enzyme SIRT1 protein deacetylase, resulting in SIRT1 activation, potentially reversing hyperglycemia and repairing damaged pancreatic cells [113, 114]. Grapefruit extract (0.5 g/kg body weight), rich in naringenin-7-O-rutinoside and kaempferol-rutinoside, improved hyperglycemia in diabetic mice treated for 6 weeks, through regulation of the hepatic glucose metabolism and reduction of pro-inflammatory gene expression (monocyte chemotactic protein-1, TNF-α, COX-2 and NF-κB) in the liver and visceral fats. The therapeutic effects were attributed to reduced miRNA expression of pro-inflammatory genes and increased DNA methylation [115]. Furthermore naringenin could possess antioxidant effects in CaCo-2 cells treated in dose range of 1 to 100μg/mL, due to reduction of miRNA expression (miRNA-17-3p, miRNA-25-5p) [116].
Anthocyanidins
Anthocyanidins are molecules with a flavonoid structure, imparting red and blue colors to fruits, flowers and leaves. They are useful for therapeutic purposes, owing to their antioxidant and anti-inflammatory properties [117, 118]. Anthocyanidins have showed anti-diabetic effects through enhancing insulin secretion, upregulating insulin signaling, decreasing gluconeogenesis, increasing expression of PPAR-α and glucose transporters (GLUT4) in the liver and downstream regulation of lipogenic gene expression [119]. Maize extract, rich in anthocyanins and ferulic acid, ameliorated high fat induced obesity in mice by altering SIRT1, AMPK, and IL-6 related metabolic and inflammatory pathways. SIRT1 was up-regulated and genes related to glucose metabolism, gluconeogenesis, lipogenesis, adipogenesis, inflammation and β-oxidation were modulated in animals treated with the extract [120].
Phenolic acids
Phenolic acids, such as chlorogenic, caffeic, vanillic, ferulic, ellagic, cinnamic acid, gallic and p-coumaric acids, are a specific class of polyphenols that is involved in mechanisms of defense against biotic and abiotic stresses. Phenolic acids are versatile molecules with promising therapeutic applications, thanks to their antioxidant and anti-inflammatory properties [121, 122]. They can influence glucose homeostasis by several mechanisms, including inhibition of glucose absorption and digestion, stimulation of insulin secretion from pancreatic β-cells, modulation of hepatic glucose output, activation of insulin receptors and glucose uptake by tissues [123]. Phenolic acids can also alters gene expression associated with glucose and lipid homeostasis [64]. Coffee is rich in phenolic acids such as chlorogenic acid and caffeic acid, and an in vitro study linked the antioxidant defenses of coffee with down-regulation of microRNA-124-3p [124]. Decreasing oxidative stress might be a potential therapeutic mechanism in protection against diabetes induced complications [125].
Caffeic acid (Fig. 2e) attenuates diabetic nephropathy through modulation of autophagy in diabetic rats supplemented with an oral dose of 40 mg/kg/day for 8 weeks. It improved blood glucose, albumin excretion and decreased renal mesangial matrix extension, with enhanced vacuolation and reappearance of autophagosomes. It up-regulated autophagy genes including microtubule-associated proteins 1A/1B light chain 3(MAP1LC3B), RB 1-inducible coiled coil protein (RB1CC1) and autophagy related gene (ATG-12), with reductions in their epigenetic regulators; miRNA-342, miRNA-133b and miRNA-30a respectively [126]. The latest research has shown amelioration of renal damage in rats via modulation of miRNA-636 using caffeic acid intake. The fasting levels of glucose, cholesterol and triglycerides were significantly decreased under caffeic acid, with down-regulation of miRNA-636 [127].
Caffeic acid phenethyl ester attenuated diabetic retinopathy by increasing the expression of extracellular superoxide dismutase (SOD3) through acetylation of histones by inhibition of HDAC1, within the SOD3 promoter region of human retinal endothelial cells [128]. Gallic acid (Fig. 2f) has been shown to reduce pro-inflammatory cytokine secretion through regulation of HAT and HDAC2 in high glucose induced human monocytes. It suppressed NF-κB activity and cytokine release along with reduced CREB-binding protein/p300 gene expression, acetylation levels and CBP/p300 HAT activity. Moreover HDAC2 expression was found to be significantly increased [129].
Resveratrol
Resveratrol (Fig. 2g) is a stilbene molecule naturally found in red grapes, red wine, peanuts and berries (blueberry, cranberry, black currant, raspberry and strawberry) [130, 131]. The pharmacological effects of resveratrol include improvements of cancer, cardiovascular disease, diabetes, obesity, nervous system disorders, aging, reproductive system diseases and irradiation injury [132, 133]. It decreases blood glucose and insulin levels, enhances β-cell function, improves insulin resistance, increases peripheral glucose utilization and decreases hepatic glucose output [134]. However, while many studies have highlighted the role of resveratrol in altering DNA methylation and histone modification, very few have linked it with metabolic diseases [73].
A study reported a decrease in inflammatory cytokines (IL-1β, IL-6, TNF-α, and IFN-γ) via DNA hypomethylation at promoter regions of cytokine genes, while the expression of anti-inflammatory IL-10 was decreased through DNA hypermethylation by resveratrol in diabetic rat aortas [135]. Resveratrol ameliorates cardiac oxidative stress in diabetes through activation of surtuin-1 (SIRT-1), which results in deacetylation of NF-κB-p65 at lysine 30 and histone 3 (H3) at lysine 9 positions. Activation of SIRT-1 may decrease binding of NF-κB-p65 to DNA and attenuate oxidative stress and cardiac hypertrophy through reduced transcription of NADPH oxidase subunits [136]. SIRT1 activation by resveratrol may also reduce levels of acetylated farnesoid X receptor (FXR) in a mouse model of metabolic disorder and thereby FXR may become activated, regulating glucose and lipid metabolism, and disrupting genes associated with diabetes and hypercholesterolemia [137].
Curcumin
Curcumin (Fig. 2h) is phenolic compound known as diferuloylmethane, isolated mainly from Curcuma longa (Turmeric). It targets several molecular pathways, causing either upstream or downstream regulation of these pathways, which make it a potential choice in the treatment of chronic disorders including cancers, metabolic diseases, diabetes, cardiovascular disorders, neurologic pathologies and infections [138, 139]. Curcumin can be a good choice as an alternative treatment in diabetes and its complications including adipocyte dysfunction, nephropathy, neuropathy, vascular diseases and pancreatic disorders [140].
It affects multiple targets to provide a therapeutic response to DM i.e. alleviation of oxidative stress and the inflammatory cascade in diabetes, activation of PPAR-γ, elevation of plasma insulin level, increasing lipoprotein lipase activity, activation of enzymes associated with glycolysis, gluconeogenesis, and lipid metabolic processes, and activation of Nrf2 function [140–142]. It also increases the expression of glucose transporters (GLUT1 and GLUT3) and thus protects against DM associated cerebral infarction [143].
Curcumin has been widely studied for its effects on epigenetic pathways in various diseases which support curcumin as a DNA hypomethylating agent, histone modification regulator by regulating HATs and HDAC and miRNA regulator [144, 145]. An in vitro study indicated that curcumin decreased high-glucose induced proinflammatory cytokine production in monocytes and suppressed NF-κB binding by reduction of HAT activity, p300 level and acetylated CBP/p300 gene expression while HDAC2 expression was increased [146].
Curcumin decreases ROS production and restores DNMT function in a cellular model of diabetic retinopathy [147]. Studies shows that hyperglycemia induced oxidative stress is involved in the etiology of diabetic retinopathy, where epigenetic regulations indicated decreased DNMT activity in the early phase of the disease, followed by increased DNMT activity and expression as a result of long term oxidative stress [148]. A curcumin analog attenuates diabetes related increase in histone acetylation, HATs activity and p300/CBP HAT expression, which leads to inhibition of the c-Jun N-terminal kinase (JNK) pathway and thus prevents against diabetic nephropathy [149].
Human studies of polyphenols in diabetes mellitus
The potential efficacy of polyphenolic compounds on glucose homeostasis has been evidenced in clinical trials (Table 3)[150]. Knekt et al. (2002), in a prospective study on flavonoid intake, showed that quercetin and myricetin may reduce the risk of DM [151]. Differently, in a Women’s Health Study, the intake of kaempferol, quercetin, myricetin, luteolin and apigenin was found to not be associated with reduction in risk of DM pathogenesis [152]. However, there are many studies showing an inverse correlation between a whole-polyphenol rich diet and the development and/or progression of DM, suggesting that its disease prevention capacity might be ascribed to groups of compounds, rather than to a single compound [153, 154]. A systemic review and meta-analysis by inclusion of 36 controlled randomized trials showed a significant impact of polyphenol supplementation on glycated haemoglobin (HbA1c) in diabetic patients, while no such effect was seen on non-diabetic or pre-diabetic patients, supporting the intake of polyphenols in the prevention of diabetic complications [155]. A clinical trial revealed that the intake of total polyphenols and total flavonoids (especially flavanones and dihydroflavonols) and stilbenes are associated with reduced risk of DM in elderly patients at high risk of cardiovascular disorders [156].
Evidence of polyphenols effectiveness in diabetes mellitus from clinical trials
Evidence of polyphenols effectiveness in diabetes mellitus from clinical trials
Increase (↑), decrease (↓), high density lipoprotein (HDL), low density lipoprotein (LDL), malondialdehyde (MDL), reduced glutathione (GSH), insulin-like growth factor-binding protein (IGFBP), fasting plasma glucose (FPG), body weight (BW), blood pressure (BP), peripheral vascular resistance (PVR), Chemokine ligand 3 (CCL3), interleukin 1-beta (IL-1β), tumor necrosis factor-alpha (TNF-α).
The effects of a polyphenol rich food supplement on oxidative stress and lipid peroxidation has been evaluated in type 2 diabetic patients with no complications [157]. 114 male and female non-smoker patients (age ranging from 40 to 65 years) with diabetes were recruited in the study and were treated with placebo (n = 58) or a food supplement containing polyphenols (n = 56) which consisted of three main ingredients, pomegranate extract (500 mg), green tea extract (300 mg), and ascorbic acid (60 mg), in addition to standard diet and exercise for 3 months. The patients treated with the polyphenol rich antioxidant supplement were found to have decreased low density lipoprotein (LDL) levels, increased high density lipoprotein (HDL) levels, decreased plasma malondialdehyde (MDA) levels, and increased total plasma levels of reduced glutathione (GSH), which suggested that this food supplement can antagonize the effects of oxidative stress and lipid peroxidation in type 2 diabetic patients.
Strawberry and cranberry polyphenols (anthocyanidins, proanthocyanidins, and phenolic acids) improved insulin sensitivity in insulin resistant, non-diabetic adults in a parallel double-blind, controlled and randomized clinical trial [158]. The experimental group was supplemented with a Strawberry and cranberry polyphenol beverage (containing 333 mg polyphenols) daily for 6 weeks while the control group was supplemented with flavor-matched control beverage without polyphenols. Of the 50 eligible human subjects, 46 (20 men and 26 women, age 40 –70 years) completed the trial. The polyphenolic supplementation resulted in improving insulin sensitivity in insulin resistant human subjects.
Borges et al. (2016) investigated the therapeutic effects of green tea polyphenols in treating residual albuminuria in diabetic nephropathy [159]. 42 diabetic subjects with albumin-creatinine ratio greater than 30 mg/g were recruited for the study, and were treated with green tea polyphenols (EGCG 800 mg) or placebo for 12 weeks. Urinary albumin-creatinine ratio was reduced by 41 % in patients treated with green tea polyphenols while a 2 % reduction was noted in the placebo group. A reduction in podocyte apoptosis via activation of the Wnt signaling pathway was the mechanism suggested for this effect.
Olive (Olea europaea L.) leaf polyphenols improved insulin sensitivity in middle-age overweight men in a randomized, placebo-controlled, crossover trial [160]. 46 men aged 46.46±5.5 years with body mass index (BMI) 28.062.0 kg/m2 were randomized to receive capsules containing olive leaf extract or placebo for 12 weeks, crossing over to the other treatment after a 6-week washout. Improvement in insulin sensitivity and pancreatic β-cell secretary capacity was observed with olive leaf polyphenol supplementation. Moreover the fasting concentrations of interleukin-6 (IL-6), and Insulin-like growth factor-binding protein 1 and 2 (IGFBP-1 and IGFBP-2) were increased with olive oil polyphenols.
Resveratrol supplementation can improve insulin sensitivity and postprandial glucose levels in patients with impaired glucose tolerance [161]. A pilot study was conducted and 10 older adults aged 72±3 years were selected for a4 week open-label study. The patients were treated with resveratrol at a daily dose of 1, 1.5 or 2 grams. As indicated by the results, fasting plasma glucose concentration was unchanged, however the peak post-meal and 3-hour glucose concentrations declined with improvements in insulin sensitivity. The body weight, blood pressure and lipids remained unchanged. Wong et al. (2016) reported that an acute intake of resveratrol improves neurovascular coupling capacity in adults with type 2 DM in randomized clinical trial [162]. 36 diabetic patients aged 40 –80 years were randomized to receive a single dose of resveratrol (0, 75, 150 and 300 mg) at weekly intervals. Results showed that 75 mg resveratrol significantly improved neurovascular coupling capacity while enhanced performance on a multi-tasking test battery was also observed with 75 and 300 mg doses. It was concluded that a single dose of 75 mg resveratrol can improve neurovascular coupling and cognitive performance in diabetic patients.
Fuchs et al. (2016) demonstrated the impact of flavonoid rich black tea and beetroot juice on postprandial peripheral vascular resistance and glucose homeostasis in obese, insulin-resistant men [163]. 16 obese, insulin-resistant males were selected for a randomized, controlled, cross-over study, and each consumed a single dose of 100 mL black tea, beetroot juice or placebo, combined with 75 mg glucose. A single dose of black tea effectively decreased peripheral vascular resistance across upper and lower limbs accompanied with lowering insulin resistance. Beetroot juice showed less consistent effects on peripheral vascular resistance and no effects on glucose homeostasis.
Tomé-Carneiro et al. (2013) investigated the impact of one-year supplementation of grape extract containing resveratrol on inflammatory related miRNAs and cytokine expression in peripheral blood mononuclear cells of diabetic and hypertensive patients with coronary artery disease in a randomized placebo-controlled, triple-blind, trial [164]. 35 male subjects were randomized to receive grape extract containing 8 mg resveratrol, grape extract without resveratrol or placebo for 1 year. The effect of the grape extract with or without resveratrol was not significant on body weight, blood pressure, serum glucose concentration, and HbA1c and lipid levels. The expression of proinflammatory cytokines chemokine ligand 3 (CCL3), IL-1β and TNF-α was reduced significantly in patients supplemented with resveratrol containing grape extract. In addition resveratrol containing grape extract alters miR-21, miR-181b, miR-663, miR-30c2, miR-155 and miR-34α.
Literature showed strong association between alterations in epigenetic pathways and the pathogenesis of DM or related complications. Therefore, targeting epigenetic pathways to regain normal metabolic homeostasis is an ideal approach for the discovery of protective agents. A large body of evidence supports that polyphenols can regulate the epigenome. The epigenetic studies of various individual polyphenols such as flavan-3-ols (EGCG), flavonols (quercetin), isoflavones (genistein), flavanones (naringenin), anthocyanidins, phenolic acids (caffeic acid and gallic acid), stilbenes (resveratrol) and curcumin showed strong preclinical potential in DM. Human studies also revealed the clinical effectiveness of polyphenols in diabetes and its complications. However, the current evidence regarding epigenetic regulation by polyphenols, and the clinical studies performed on their protective activity against DM, are insufficient to draw a definitive conclusion on their role in the treatment of metabolic diseases. Thus, this area of research requires more in depth information on the relative potency and efficacy of these compounds, alone or in combination with other drugs.
Funding
This research received no external funding.
Conflicts of interest
The authors declare no conflict of interest.