
Abstract
OBJECTIVE:
Pregnant non-Hispanic blacks (NHB) have increased vaginal microbiome diversity compared to non-Hispanic whites (NHW) which may contribute to increased preterm birth. Cervical microbiome diversity is poorly characterized in pregnancy, therefore our objective was to correlate cervical microbiota diversity with cervico-vaginal inflammation by race and delivery timing.
STUDY DESIGN:
Pregnant women were recruited in the first and second trimesters. A sterile cervical swab and saline lavage were collected at a single time point. Using 16S rRNA sequencing, Chao1 and Shannon Diversity (SDI) indicies were measured and compared by race and delivery timing (preterm vs. term delivery). Cervico-vaginal inflammatory markers were also compared by race and delivery timing. Spearman correlation coefficients between cervical microbiome diversity and cervico-vaginal inflammatory markers were calculated.
RESULTS:
Of the 51 subjects, 39 (76%) were NHB and 12 (24%) were NHW. Cervical microbiota SDI was significantly higher in NHB compared to NHW (0.5 vs. 0.1; p = 0.03). However, there were no difference in Chao1 diversity or cervico-vaginal inflammatory markers by race or delivery timing.
CONCLUSION:
Our findings suggest the cervical microbiota diversity during pregnancy differs by race. Larger cohort studies will further determine if altered cervical diversity is part of the pathogenesis of PTB and explains race disparities.
Introduction
Non-Hispanic Black (NHB) women have nearly double the risk of preterm delivery (birth before to 37 weeks of pregnancy) compared to non-Hispanic White women (NHW) [1, 2]. In 2015, the overall preterm birth (PTB) rate in the US was 9.6%, while the rate among NHB women was 13.4% [1]. Despite decades of research, the disparity remains poorly understood. Controlled studies indicate that social determinants of health such as income and health habits do not fully account for the racial disparity in PTB [3–6]. Therefore, attention has turned to other factors that could further explain the PTB disparity.
Race differences in genital tract flora is an area of growing interest that may play a role in the PTB disparity. Large population studies show 2–3 fold higher rates of bacterial vaginosis (BV) in black women [7, 8]. BV is associated with an increased cervico-vaginal inflammation and PTB; however, it is unclear if there is a link between the increased rates of BV and PTB in the NHB women [9, 10]. Previously, the diversity of the vaginal flora was defined by culture and microscopy approaches; however, next generation sequencing technology has revealed that much of the vaginal microbial community, called the microbiota, is poorly or non-cultivatable [11–14]. Consistent with the BV data, studies utilizing sequencing technology demonstrate increased vaginal microbiome diversity in NHB compared to NHW during pregnancy, however it is unclear if this increased diversity is correlated with preterm birth [15, 16].
A contemporary model of PTB incorporates a pathogenic or dysbiotic vaginal microbiota that alters PTB risk via increased inflammation and ascension from the vagina to intrauterine compartment via the cervix. Missing from this model, however, is a robust understanding of the cervical microbiota as the gatekeeper between the vaginal and uterine compartments. We hypothesized that the cervical microbiota diversity and cervico-vaginal inflammatory markers are increased in NHB women compared to NHW and in preterm compared to term birth. We further hypothesized that cervical microbiome diversity is correlated with cervico-vaginal inflammatory markers. In this study, we addressed these hypotheses in a prospective cohort with these aims: 1) To identify differences in cervical microbiome diversity in NHB compared to NHW pregnant women and in term compared to preterm delivery timing and 2) To measure correlations between the cervical microbiome diversity and cervico-vaginal inflammatory markers.
Materials and methods
We conducted a prospective cohort study comparing NHB and NHW pregnant women. The Duke University IRB (Pro00052778) approved the study prior to enrollment of any participants. Women with a history of spontaneous preterm birth before 34 weeks (high risk) and with a history of prior term delivery (low risk) were recruited between 6 0/7 weeks and 25 6/7 weeks. Additional inclusion and exclusion criteria are detailed in Table 1. After informed consent, a sterile speculum exam was performed for sample collection. The external os of the cervix was sampled with sterile dacron swab for cervical microbiome analysis. Following swab collection, a cervico-vaginal lavage was performed with 10 ml of sterile saline instilled into the posterior vaginal fornix and re-aspirated for cervico-vaginal inflammatory marker analysis. Swabs were stored at –80°C and bacterial DNA was extracted on all samples at the conclusion of enrollment to avoid batch effects. The cervico-vaginal lavage fluid was centrifuged at 400 RCF for 10 minutes, and the supernatant was stored at –80°C for batch analysis at the conclusion of enrollment.
Eligibility Criteria
Eligibility Criteria
The cervical sample processing, sequencing, analysis steps were as previously published [17]. Briefly, total genomic DNA was extracted from cervical swabs using a commercial bead-beating method (Zymo Research Soil Microbe DNA Kit, Irvine, California). The V4 region of the 16S rRNA gene was amplified in duplicate independent reactions using 12 nucleotide barcode-indexed primers and previously described standardized PCR conditions for the Earth Microbiome Project [18]. PCR amplicons were pooled in equimolar concentrations, and the pooled amplicon of interest was purified by gel extraction. Sequencing was performed using a 2×250 paired end (PE) V2 Miseq protocol (Illumina).
Sequences were split, quality-trimmed, demultiplexed, and chimera-reduced using QIIME tools (V1.9; [19]. High-quality sequences sharing≥97% nucleotide sequence identity were clustered into operational taxonomic units (OTUs) using USEARCH (version 6), aligned to the Greengenes database (version 13.8.99), and representative sequences were given a taxonomic assignment using BLAST against the SILVA bacterial database (Release 111) [20–22]. Sparse OTUs with <3 occurrences in at least 20% of samples were removed from subsequent analyses with the rationale that they were unlikely to significantly contribute to the overall composition and inferred metabolic capacity in any one sample. However, for alpha diversity calculations, only singleton OTUs were excluded. Counts were normalized using the cumulative sums scaling approach (percentile p = 0.5, determined using the ‘cumNormStatFast’ function) in the metagenomeSeq package (version 1.12.0) [23].
Microbial composition and diversity were determined using tools within the phyloseq (version 1.14.0) and metagenomeSeq packages (version 1.12.0; [23, 24] All analyses were performed using R statistical software. Statistical significance was considered P < 0.05, with Benjamini-Hochberg correction for multiple testing.
Cervico-Vaginal inflammatory markers analysis
Luminex multiplex assay was performed on cervico-vaginal lavage specimens to measure concentrations of a battery of cervico-vaginal cytokine and chemokine inflammatory markers known to play a role in PTB. Lavage cytokine and chemokine concentrations were assayed in duplicate using a 10-analyte multiplex assay panel (Milliplex MAP human cytokine/chemokine magnetic bead multiplex assay panel (HCYTOMAG-60K-10) performed according to the manufacturer’s recommended protocol and read using a Bio-Plex 3D Suspension Array reader (Bio-Rad). Data were analyzed using Bio-Plex Manager software (Bio-Rad).
Statistical methods
Demographic characteristics and birth outcome data were compared by race using Mann-Whitney test or Fisher’s exact test. Median and IQR of SDI, Chao1, and cervico-vaginal inflammatory marker concentrations were calculated and compared by race (NHB vs. NHW) and delivery timing (term vs. preterm delivery) using Mann-Whitney statistical test. Correlations between cervical microbiome SDI and cervico-vaginal inflammatory marker concentration were calculated using Spearman correlation coefficients. Spearman correlation coefficients were compared by race and delivery timing. Analysis was performed in R (version 3.3.1[25]).
Results
Fifty-one pregnant women including 39 NHB and 12 NHW provided samples for analysis. The median gestational age at sample collection was 15.5 weeks and the median gestational age at delivery was 37.5 weeks such that there was a median of 22 weeks between sample collection and delivery. Among the NHBs, 29 (74.4%) were at high risk for preterm birth, while 5 (41.6%) NHW participants were high risk. Demographic characteristics did not vary by race. NHB babies were smaller and born at an earlier gestational age than NHW babies. Importantly, these observations are driven by the fact that all of the 14 preterm births were born to NHB women. There were no differences in other birth outcomes by race (Table 2).
Demographics and Birth Outcomes by Race
Demographics and Birth Outcomes by Race
The median cervical microbiome Shannon Diversity Index (SDI) was higher NHB compared to NHW (0.5 vs. 0.1, p = 0.03; Fig. 1). However, there was no difference in cervical Chao1 diversity by race. There was a trend toward higher cervical SDI in pregnancies that delivered preterm compared to pregnancies that delivered at term; however, this difference did not meet statistical significance (0.9 vs. 0.4, p = 0.06; Fig. 2). There was no difference in cervical microbiota Chao1 by delivery timing (Table 3).
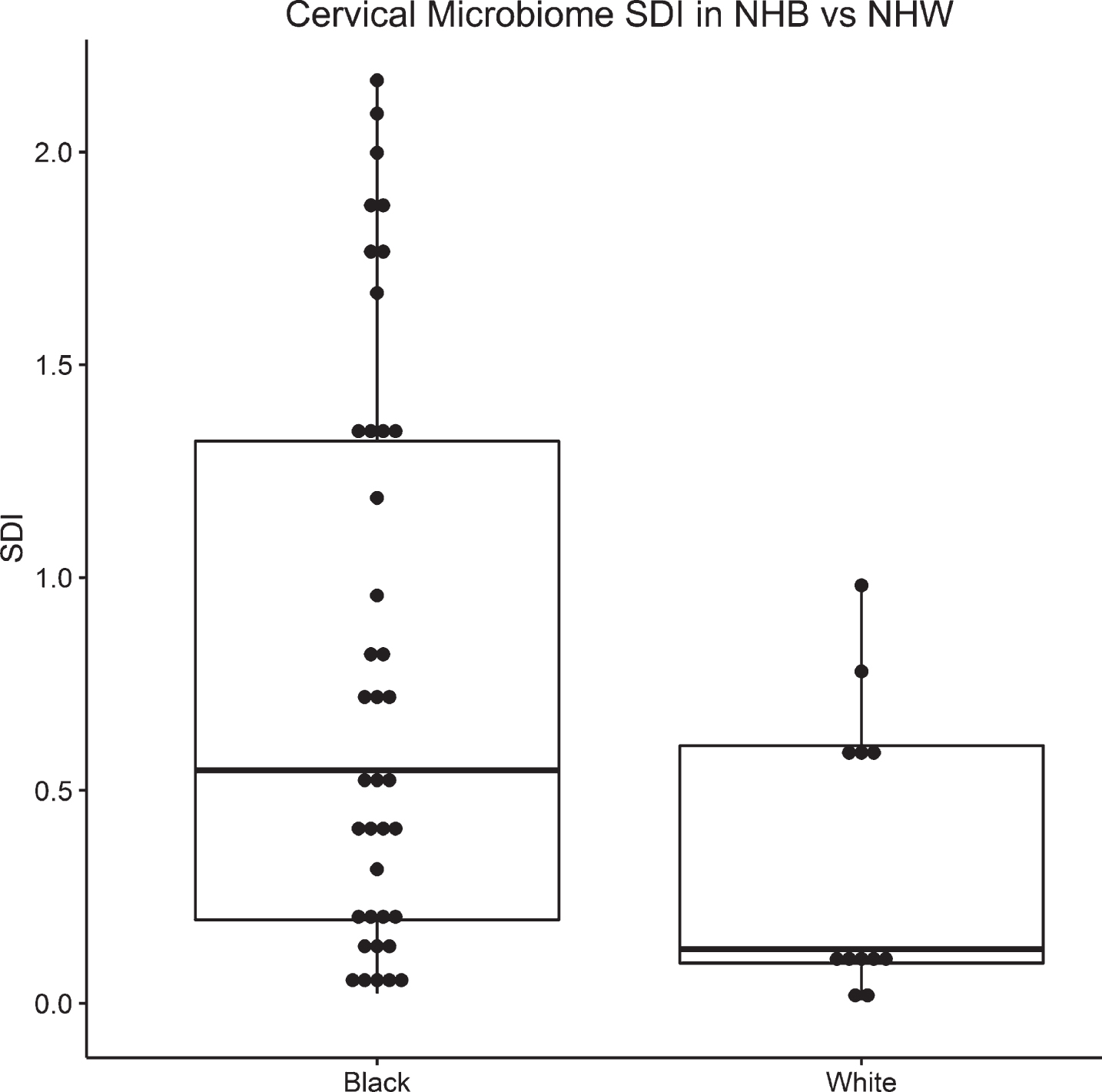
Box plot of SDI by race. Median and IQR depicted via line with the box, top and bottom of box, respectively.

Box plot of SDI by birth outcome. Median and IQR depicted via line with the box, top and bottom of box, respectively.
Median Cervical Microbiome Diversity by Race and Delivery Timing (median, interquartile range)
There was no difference in cervico-vaginal inflammatory markers by race or delivery timing. There was a weak correlation between IL1-β and cervical microbiome diversity (rho = 0.38, p = 0.05). No other correlations between cervico-vaginal inflammatory marker concentrations and cervical microbiota diversity were noted (Table 4).
Cervico-vaginal Inflammatory Marker Concentration (pg/mL) by Race, Delivery timing in correlation with cervical microbiome diversity
The central aims of this study were to identify differences in the cervical microbiota diversity by race or delivery timing to determine if cervico-vaginal inflammatory markers were correlated with cervical microbiota diversity. We conducted a prospective cohort study measuring cervical microbiota diversity and cervico-vaginal inflammatory markers in NHB and NHW pregnant women. Consistent with our hypothesis, NHB had increased SDI cervical microbiota diversity compared to NHW. We also noted a trend in the association between preterm delivery and an increased cervical microbiota SDI diversity. A larger sample size may more conclusively determine if the cervical microbiota diversity is indeed increased before preterm birth and further understand how race differences in the cervical SDI may play a role in race disparities in PTB. When comparing comparing preterm and term, in order to detect a 0.5-point increase in SDI, we would need at least 25 patients in each group. To detect a 2.5-point increase in Chao 1, we would need at least 263 patients in each group.
We detected a difference in the cervical SDI diversity by race but not in the Chao1 index. The discrepancy between SDI and Chao1 diversity measures is likely due to the characteristics of these indices. Chao 1 is an unweighted measure diversity that reflects the number of distinct organisms identified [26], while the SDI is a weighted measure that incorporates not only the number of distinct organisms but also their proportionality [27–29]. Our findings suggest the race and birth timing differences in cervical microbiota diversity may be driven by relative abundance of microorganisms rather than differences in the absolute number of species.
Also contrary to our hypothesis, cervico-vaginal inflammatory markers were not different by race or delivery timing. Inflammation is theorized as an important mechanism by which altered flora leads to PTB. However in our study, cervico-vaginal inflammatory markers did not correlate with race or birth timing. This finding suggests that any link between cervical microbiota diversity and race or birth timing may be mediated by mechanisms outside inflammatory pathways measured herein.
Our findings must be viewed in the light of several additional important limitations. Due to cost and practicality, we applied 16S rRNA amplicon sequencing, providing limited OTU and taxonomic information. Genomic variation among species or even clones within species may contribute to functional diversity in the cervical niche. Thus, greater differences in the cervical microbiomeby race and delivery timing in our cohort may exist that will be identified through future shotgun DNA and RNA sequencing. The current observations are also limited to the analysis of samples froma single time point. Ascending flora and inflammation may have occurred after specimen collection altering the cervical microbiota later in pregnancy. During the non-pregnant state, the vaginal microbiota undergoes natural variation that stabilizes during pregnancy [30]. The extent of stability of the cervical microbiota during pregnancy cannot be inferred from the current study.
The generalizability of our findings is limited by a cohort that only includes NHB and NHW women in the Durham NC area. Much of the literature surrounding race differences in preterm birth is centered on NHB and NHW women; however, PTB affects all races. Due to practical limitations and to correlate our work with prior studies, we focus on women who are self-described as NHB or NHW for the current analysis. We recognize that focusing our investigation to these groups limits the generalizability of our findings to a narrow window of patients. Additionally, in an era where there is increasing ethnic mixing relying on patients to self-identify their race could lead to misclassifications. However, the studies that demonstrate higher rates of preterm birth in NHBs are based on self-report therefore using this method to assign race is most consistent with the literature. Despite these important limitations, our work provides novel data on the cervical microbiota, a part of the pregnancy microbiota that was previously very poorly characterized. Our work also suggests further longitudinal studies evaluating the cervical microbiome and PTB disparities are needed.
Understanding the etiology of the disparity in preterm birth will not only help narrow the gap in birth outcomes among NHB babies but also yield important insights into the pathogenesis of preterm birth in general. Although important advances have been made in PTB, currently our most powerful tool for risk stratification is pregnancy history, and our only preventive therapies are progesterone and cerclage. Cervical microbiota analysis is a promising avenue of research that may reveal additional important insights into the PTB disparity and ultimately lead to more precise risk stratification and targeted PTB prevention.