
Abstract
BACKGROUND:
Children born prematurely (<37 gestational weeks) are at risk for a variety of adverse medical events. They may experience ischemic and/or hemorrhagic events leading to negative neural sequelae. They are also exposed to repeated stressful experiences as part of life-saving care within the neonatal intensive care unit (NICU). These experiences have been associated with methylation of SLC6A4, a gene which codes for serotonin transport proteins, and is associated with anxiety, depression, and increased incidence of autism spectrum disorders.
The purpose of this study was to examine the effects of altered serotonin levels on behavioral and neuroanatomical outcomes in a neonatal rodent model with or without exposure to hypoxic-ischemic (HI) injury.
METHODS:
Wistar rat pups were randomly assigned to either HI injury or sham groups. Pups within each group were treated with a chronic SSRI (Citalopram HBr) to simulate the effects of SLC6A4 methylation, or saline (NS). Subjects were assessed on behavioral tasks and neuropathologic indices.
RESULTS:
HI injured subjects performed poorly on behavioral tasks. SSRI subjects did not display significantly greater anxiety. HI + SSRI subjects learned faster than HI+NS. Histologically, SSRI subjects had predominantly larger brain volumes than NS.
CONCLUSION:
SSRI treated subjects without injury showed patterns of increased anxiety, consistent with theories of SLC6A4 methylation. The paradoxical trend to improved cognition in HI+SSRI subjects relative to HI alone, may reflect an unexpected SSRI neuroprotective effect in the presence of injury, and may be related to serotonin-induced neurogenesis.
Introduction
Hypoxic-ischemic (HI) brain injury is a perinatal complication associated with 25% of neonatal deaths globally [1]. In developed countries, 30% of all neonatal encephalopathy is related to HI and that figure rises to nearly 60% in developing countries [2]. HI occurs when blood and/or oxygen flow to the brain is impaired, leading to energy failure and cell death [3–5]. In premature infants, the fragile, immature neurovascular system increases the risk of HI from intraventricular/periventricular hemorrhages, as well as inflammation and oxidative stress following ischemic reperfusion [6, 7]. Most HI injuries in preterm infants result in white matter damage, though volume reductions of gray matter in cortical and subcortical regions are also seen, particularly in late preterms [8, 9]. HI injuries in late preterm infants (34–37 weeks’ GA) are less common, with resulting pathology similar to the deep nuclei grey matter damage seen in term infants with HI [6, 10–13].
Premature infants are also exposed to numerous stressors including excessive light and noise levels, parental separation and high levels of pain-related stress that are part of routine life-saving care within the neonatal intensive care unit (NICU) [14, 15]. Chronic stress can induce long-lasting effects on neuroendocrine and behavioral responses [16, 17]. One pathway for these effects involves stress-induced epigenetic changes that can enhance or repress gene activity [18]. Recent research suggest that stress-induced epigenetic modifications may have a role in neurodevelopmental outcomes. One particular candidate involves DNA methylation of SLC6A4, a gene which encodes for serotonin transport proteins. Since the serotonergic system is critical in socio-emotional stress response [19, 20] and serotonin receptors are found throughout the central nervous system –appearing early during gestation and then rapidly developing after birth, it is speculated that epigenetic mechanisms regulating SLC6A4 transcription may be associated with early adversities and degraded behavioral and neurodevelopmental outcomes [21]. Serotonergic pathways are found throughout the central nervous system and develop early and rapidly [22]. This system is managed by feedback mechanisms through the serotonin transport protein coded by SLC6A4 [22]. Allelic variations of SLC6A4 can affect serotonin reuptake, and also modulate socio-emotional outcomes and emotionality in early life [23]. The purpose of the current study was to determine the effects of chronic stress and altered serotonergic tone on behavioral and neuroanatomical outcomes in a neonate rodent model with or without exposure to HI brain injury. HI or sham injuries were induced following established experimental methods. This was then followed by repeated saline or selective serotonin reuptake inhibitor (SSRI) treatment. Chronic exposure to SSRI injections was used to mimic epigenetic alterations associated with chronic stress, based on evidence that chronic early SSRI exposure downregulates long-term serotonin transport (similar to SLC6A4 methylation) [24].
Methods
Subjects
Eighteen time-mated female Wistar rats were de-livered to the Bousfield vivarium at University of Connecticut from Charles River Laboratories (Wilmington, MA, USA): 6 on embryonic (E) day 4, 6 on E5 and 6 on E6. Dams were housed in single cages with a 12-hour light/dark cycle and food and water provided ad libitum. Pups were born on approximately E22. On postnatal day 6 (P6), pups were culled to 10 litters of 4 males and 4 females and weaned on P21 to be housed in pairs of gender and treatment, and finally single-housed at P49 (adulthood) for behavioral testing.
Experimental design
On P6, an age that corresponds to a 32–35 weeks gestational age in premature infants, pups were randomly submitted to a HI insult or sham (SH) procedure and, were also assigned to chronic SSRI or saline administration. Therefore, four groups of 20 animals each (10 males and 10 females) were studied: HI chronic SSRI, HI saline, sham chronic SSRI and SH saline.
Induction of Hypoxic-Ischemic Injury
Pups were anesthetized with isoflurane 2.5% and local bupivacaine subcutaneously (SC). A vertical incision was made on the neck at midline. The right common carotid artery was identified and cauterized to restrict blood flow to the right cerebral hemisphere. The incision was sutured, and footpads tattooed for ID. Sham animals underwent a similar surgical procedure without cauterization, and with comparable duration of isoflurane (maximum 9 minute exposure in all groups to avoid confounding neuroprotective or neurotoxic effects). Pups in the chronic SSRI condition were injected SC with 10 mg/kg of citalopram HBR diluted in 9% normal saline (NS) to a volume of 1/100 their weight (15 grams = .15 mL diluted citalopram HBR) immediately after surgery and then daily until P21. Shams were injected subcutaneously with a volume of 1/100 of their weight of 9% NS daily until P21.
Post-surgery, pups were returned to dams to nurse for 60 minutes. After feeding, hypoxic conditions were induced in the HI group via an airtight container on a warming tray (maintained at nest temperature) containing 8% oxygen with a nitrogen balance for 90 minutes. Sham pups were placed in an open container on a warming tray for 90 minutes. Subjects were then returned to the dams (Fig. 1).

Description of animal randomization into 4 conditions.
Sensorimotor task: Rotarod (P35–39)
The rotarod task was used to measure balance, motor coordination and motor learning. Animals were placed on a rotating rod gradually accelerating from 4 rotations per minute (rpm) to 40 rpm, during a 5-minute period. Animals were given four trials per day for three days. Latency to falling off the rod and rpm at time of fall was recorded per trial (in seconds) and averaged for later analysis.

Sequence of tasks animals underwent.
2.3.2.1. Elevated Plus Maze (EPM) (P54–P58): The EPM was used to assess general anxiety behavior. Rats were transported to the testing room inside their home-cages to minimize distress and remained in their cages until testing. The EPM was constructed of black plastic and elevated 50 cm above the floor. The apparatus consisted of four arms, 50 cm long and 10 cm wide aligned perpendicularly. Two arms were enclosed by 30 cm high walls and the other two arms were exposed. The exposed arms each had a 0.9 cm lip to prevent subjects from falling off. The maze was placed in the center of a quiet room with only ambient lighting.
Each experimentally naïve subject was placed in the center of the platform, facing an open arm and were allowed to freely explore for 5 min. After each observation, the EPM was cleaned with Virkon disinfectant cleaner to remove debris and scent cues left from the preceding subject. Each subject was re-corded through a video camera and video files stored in a secure computer. Two variables of interest were analyzed separately: a) ratio of entries into open arm versus closed arm and b) difference of time spent on open arm versus on closed arm (measurement of the inclination to stay on the open arm).
2.3.2.2. Open Field (P61–65): The Open Field task is a simple sensorimotor test used to determine general activity levels, gross locomotor activity, and exploration habits in rodent models of CNS disorders [25]. Assessment took place in a square Plexiglas box. The field was marked with a grid and squares with a center square and a middle square surrounding the center discernable by different colored tapes and an outside square reaching the walls of the box. The animal was placed in the center arena and allowed to freely move about for 10 minutes while being recorded by an overhead camera. The footage was then analyzed by an automated tracking system for the following parameters: distance moved, velocity, and time spent in pre-defined zones.
2.3.2.3. Marble-burying (P66–69): The marble-burying behavioral task is another measure of anxiety or obsessive-compulsive traits. Rodents were placed for 30 minutes in a standard cage filled with wood chip bedding 5 cm deep. There were 10 marbles evenly spaced in the cage. After the study period, the number of marbles buried was measured.
Learning and memory assessments
2.3.3.1. Water escape task(P70–74): Screening for general visual or motor deficiencies was conducted in a one-day water escape task. This task is completed prior to the Morris Water Maze to ensure that all animals are capable of swimming as well as seeing the escape platform. The task used an oval tub (40.5×21.5 in.) filled with room temperature water, with a visible escape platform at the end opposite from where the animal was introduced to the water. Once placed in water, animals were timed until they reached the platform or 45 seconds had elapsed. Those who were not able to locate the platform during the 45-second swim were led to it and allowed to detect cues for 2 seconds. Cues included colored tape and brighter lighting next to platform location.
2.3.3.2. Morris Water Maze (MWM) (P75–79 & P82–86): The MWM task was used to assess spatial learning and memory [26, 27]. Animals were divided into two matched cohorts containing comparable numbers from all groups, due to the large sample size and testing limitations. Cohort 1 was tested between P75–79 and cohort 2 between P82–86. The MWM requires animals to use spatial cues outside the maze (large black geometric shapes painted on the surrounding walls, lighting and the experimenter) to locate a platform. Testing was conducted in a 48-in. diameter inflexible black tub that contained a 6-in. diameter escape platform submerged 1 in. below the water line, making it invisible to the animals. The escape platform was located in the same quadrant of the tub for each trial. There were no intra-maze cues, requiring subjects to use external room-based spatial strategies. Animals participated in four trials per day where the start position varied (North, South, East or West). Start position was not repeated in the same day, and the order of start positions was not repeated on subsequent days. Each animal’s attempt was recorded with a Sony Digital 8 video camera connected to a Dell Dimension E21 computer with SMART Version 2.5 tracking software recording animal latency (measured in s) to reach the platform, as well as average swim speed (measured in cm/s). During each trial the animal was given 45 s to locate the submerged platform. If the animal failed to reach the platform it was guided to the platform and allowed to sit and survey the room for 5 s assessing cues including a shape painted on the particular wall near the platform or, lighting.
Histological examination
After behavioral testing was complete, rats were anesthetized with ketamine (100 mg/kg) and xylazine (15 mg/kg), and transcardially perfused with 0.9% saline solution followed by 10% buffered formalin. Brains were extracted from the skulls and placed in 10% formalin. Due to the large n, only shams were quantified specifically to determine the effect of the SSRI versus normal saline to brain volume. Brains were sliced coronally using a Leica VT1000 vibratome at a thickness of 60μm. Every other slice was mounted on a chrom-alum subbed slide followed by Nissl staining. To measure gross structural volumes of the structures associated with emotion (caudate putamen, globus pallidus, amygdala, hippocampus), a Stereo Investigator Microbrightfield software and an Axio 2 Zeiss Microscope were utilized. Volumes were quantified using 2.5x magnification with Cavalieri’s Estimator software and a grid overlay. Fewest number of sections were counted to achieve a coefficient error of less than 0.05 which was considered stereologically valid. Every third mounted slice was examined and all measurements were performed blind to treatment group.
Statistical analysis
Statistical analyses were conducted in SPSS 24.0 software (IBM, Armonk, NY, USA) and R [28]. Figures were produced using the package ggplot [29]. Data were collapsed across sex as appropriate for some tasks when there was not enough evidence showing sex effect. Rotarod task was analyzed by using a 2 (days)×4 (tests per day) repeated measures ANOVA. The 4 trials performed in day 1 were treated as the warm-up period and therefore eliminated in the statistical analysis. For the variables of interest in the EPM task a linear regression was used. The Open Field task was analyzed using one-way ANOVA and the Marble Burying task by a linear model with certain box-cox transformation applied. Morris Water Maze was analyzed by linear mixed-effect model where models of surgery/treatment interacting with days were considered since we are interested in different performance in learning rate (slope). Histology measures were analyzed using independent t-tests, since only sham SSRI and sham NS were examined. Results are reported based on the statistical analysis methods previously mentioned followed by specific comparisons, using the variables Sex (between, 2 levels; M/F) and Injury (between, 2 levels; HI/Sham), and Treatment (between, 2 levels; SSRI/NS). Interactions between Injury and Treatment, if exists, were also reported.
Results
Seventy-two animals (40 female and 32 male) were tested on a battery of behavioral paradigms. In some tasks, there were significant differences between sex, those that showed no significant difference were collapsed together. Based on a priori hypotheses, results are provided for HI vs. sham within saline and SSRI groups separately, as well as between SSRI and saline groups with sham injury.
Sensorimotor (Rotarod)
For this analysis, data were collapsed across gender because no statistically significant effect of sex was observed (t = 1.301, df = 70, p = 0.198). Then, no significant overall injury (t = 1.319, df = 69, p = 0.191) or treatment effects (t = –0.899, df = 69, p = 0.372) were noted (Fig. 2). Animals in the Sham-NS group showed higher latency to fall than those in the Sham-SSRI group but this difference did not reach statistical significance (Fig. 3; t = –1.36, df = 68, p = 0.178).
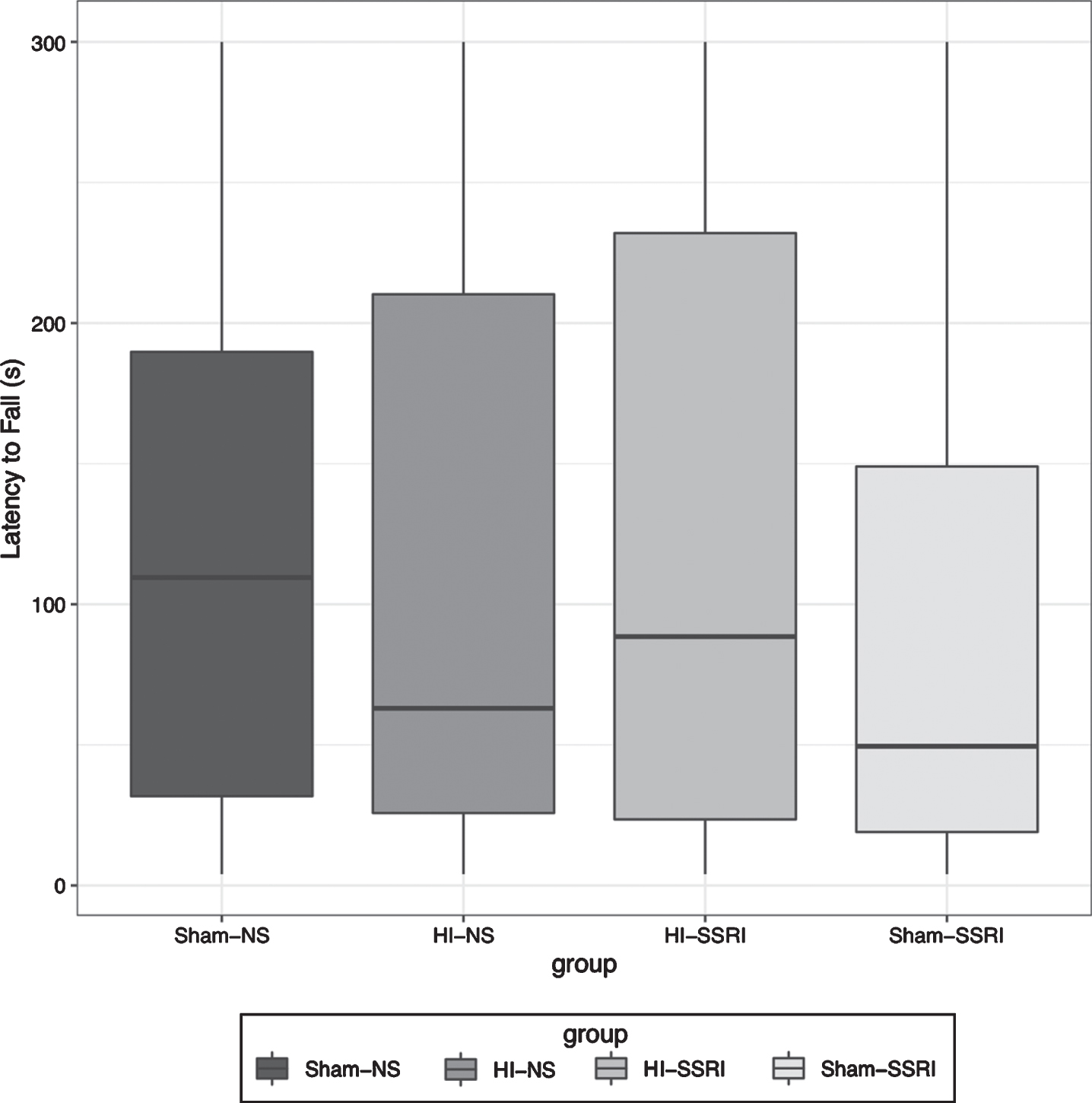
Rotarod, Boxplot of latency to fall after dropping day 1 and treating day 2-3 as repeated measures. (n = 72; Sham-NS n = 21; HI-NS n = 15; HI-SSRI n = 17; Sham-SSRI n = 19).
Females exhibited less anxious behavior compared to males on the EPM. Specifically, they spent more time in open versus closed arms (t = 7.96, df = 70, p < 0.001), and showed a higher ratio of open v. closed entries (Fig. 4, t = 5.76, df = 70, p < 0.001). Sham-SSRI group exhibited non-significantly more anxious behavior compared to Sham-NS group since they spent less time in open versus closed arms (t = –1.556, df = 67, p = 0.124), and showed a lower ratio of open versus closed entries (t = –1.467, df = 67, p = 0.147) but this was not statistically different.

EPM, Boxplot for ratio of entries into open arm against closed arm (#OpenEntry/#ClosedEntry) (Total n = 72; Female n = 40; Male n = 32).
In the Open Field (OF) test, none of Sex, Injury and Treatment was statistically significant in any zone, and there is no difference between Sham-NS group and Sham-SSRI group. In the Marble Burying task, box-cox transformation (lambda = 0.3) was used to make the residuals normally distributed. Female animals buried fewer marbles than males (Fig. 5, t = 2.301, df = 70, p = 0.024), but neither Injury nor Treatment reaches significance. Also, there is no difference between Sham-SSRI and Sham-NS group (t = 0.254, df = 67, p = 0.800).
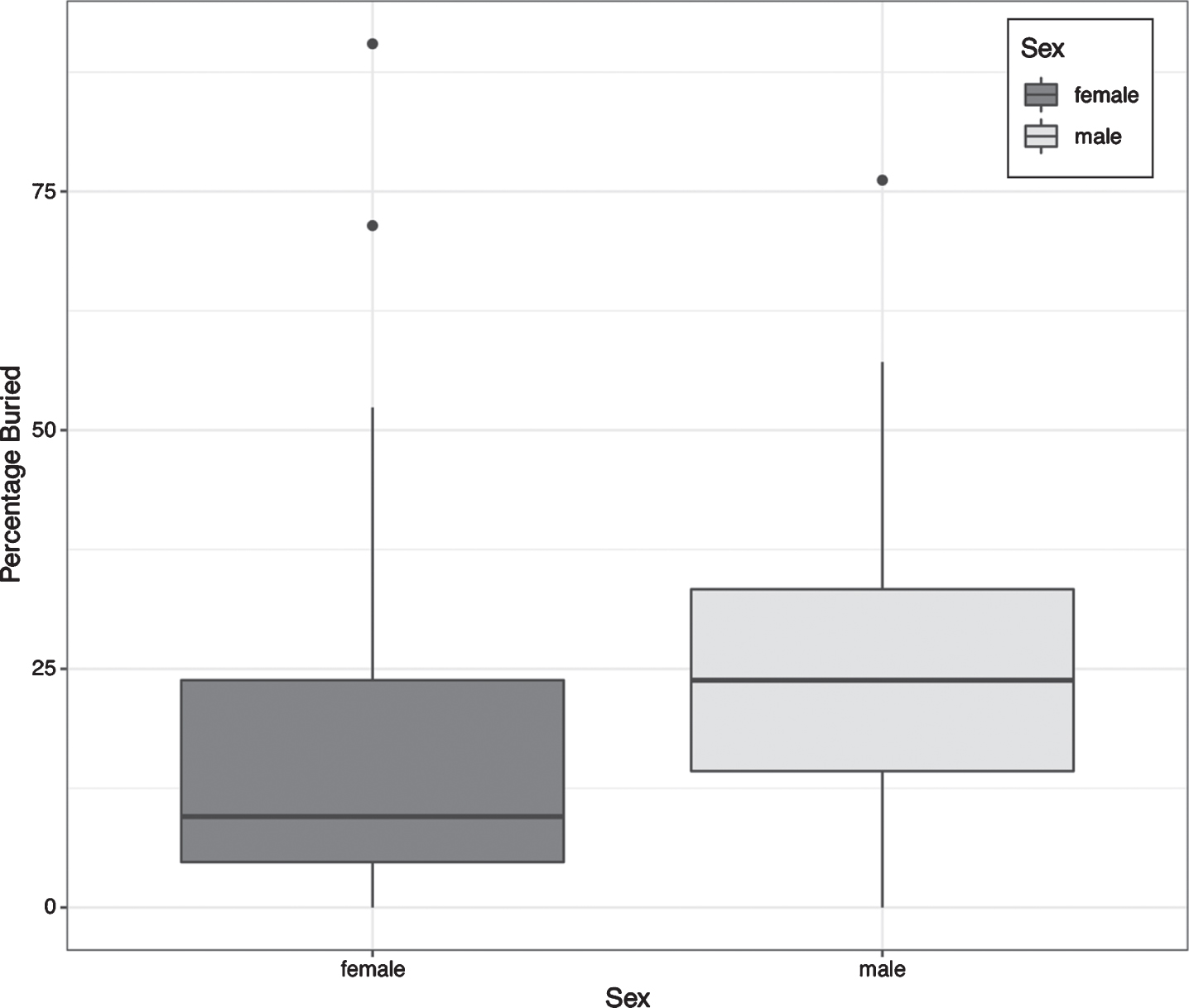
Marble Burying, Boxplot of percentage buried across sexes (Total n = 72; Female n = 40; Male n = 32).
Animals subjected to HI injury had more difficulty on the MWM when compared to Sham animals (t = –2.42, df = 33, p = 0.021) (Fig. 6). As clearly shown in Fig. 7, HI groups perform worse at day 1. At day 5, HI-NS group still performed much worse than Sham groups, but the gap between HI-SSRI group and Sham groups was no longer as considerable. Subjects with both HI and SSRI learned more quickly than those with only the HI injury (t = –1.601, df = 284, one-sided p-value = 0.055). Specific comparison between Sham-SSRI and Sham-NS groups revealed no statistical difference (t = 0.43, df = 284, p = 0.668)
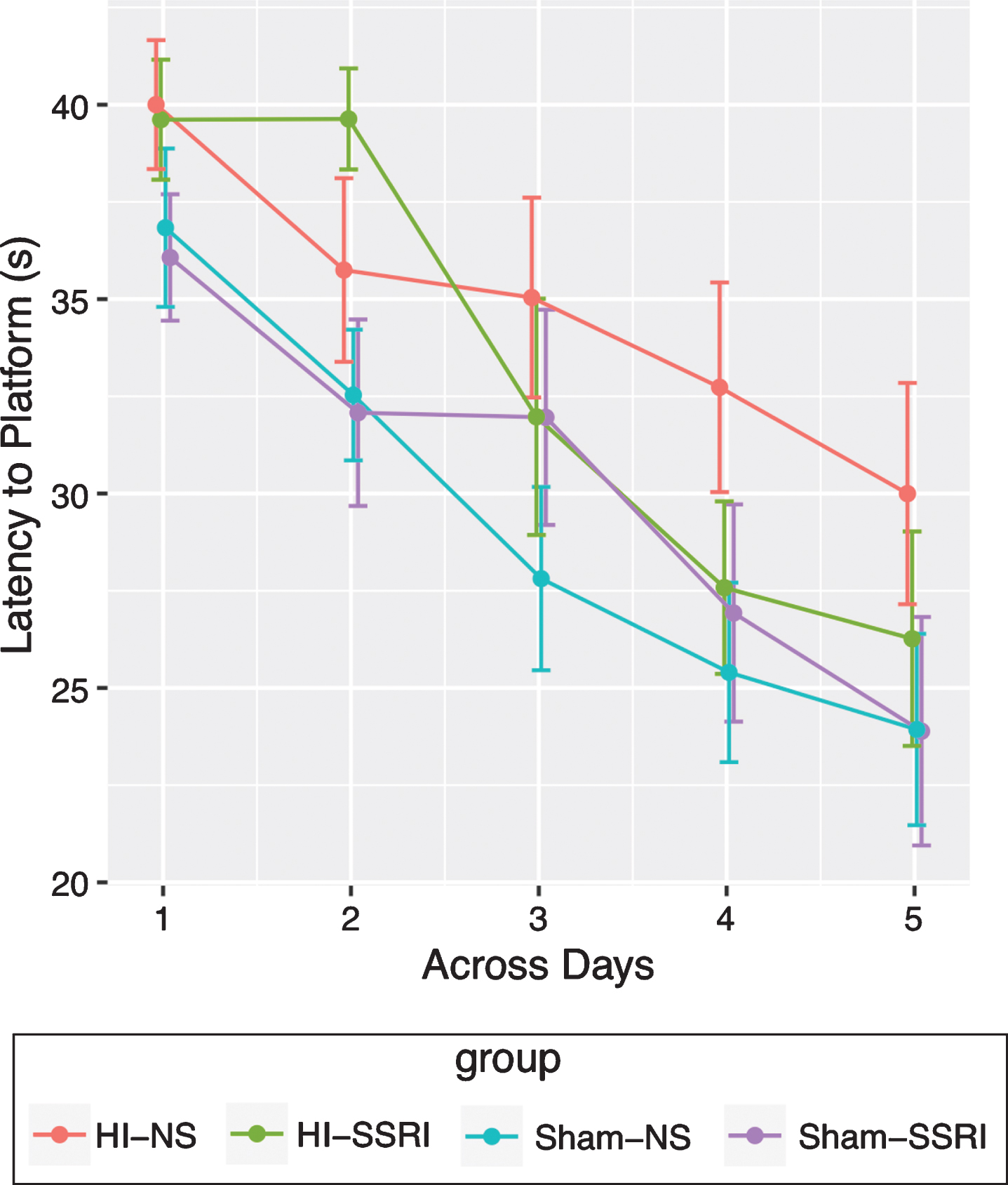
Morris Water Maze (mean level for each group across days with error bar representing standard error) (n = 72; Sham-NS n = 21; HI-NS n = 15; HI-SSRI n = 17; Sham-SSRI n = 19).
Since the treatment effect of interest related to the comparison between SSRI and NS, only surgical sham rats were examined histologically. No significant treatment effect of the Citalopram for either left or right side in any of the structures examined (caudate putamen and anterior cingulate gyrus, globus pallidum, dorsal hippocampus or the amyg-dala) was detected. It is important to note that structural integrity of tissue varied, and as such, not all structures related to emotionality could be examined. In addition, the number of samples varied per structure. Of interest is that although these volumes did not reach statistical significance, those who received the Citalopram trended toward larger brain volumes in striatal and limbic structures assessed (except for the hippocampus).
Discussion
Recent research suggests that infants who undergo numerous stressful events per day including skin-breaking procedures while in the NICU demonstrate reduced serotonin expression later in life [21]. The current study sought to examine the effect of altered serotonergic tone in rodents who underwent a hypoxic-ischemic injury at P6, an age that mimics a moderately preterm infant.
Our findings demonstrate that exposure to SSRIs during vulnerable stages of neurodevelopment may evoke neuroprotective effects on brain development and behavior. Previous studies have found that neonatal exposure to SSRIs results in alterations in behaviors associated with emotion, such as increased depressive-like behavior, decreased exploration and increased anxiety [30]. Perinatal exposure to citalopram results in behavioral alterations that persist into adulthood [31]. While the behavioral effects of the hypoxic-ischemic injury were consistent with previously reported studies, the results following chronic administration of citalopram in combination with the injury in this study were novel and suggest a trend toward possible neuroprotective effect of SSRI in the presence of HI injury. This could reflect beneficial evolutionary effect of early chronic stress in the presence of injury, even though the effects of chronic SSRI (mimicking chronic stress) were deleterious in healthy (sham) subjects.
Specifically, rodents injected with citalopram from P6–P21, an age in which 5-HT ontogeny and synaptogenesis occurs [32] displayed learning impairment compared to sham saline rodents. In contrast, subjects who sustained an HI injury and received the citalopram trended toward learning more quickly on the MWM than subjects with an HI injury who received saline. Thus, it is possible that the SSRI may have acted as a neuroprotectant with a p-value 0.055. Additional research may prove useful in determining neuroprotective capacity of SSRIs in learning and memory tasks for those with brain injuries.
As previously reported, we observed some significant differences between males and females, on specific tasks. Indeed, females displayed less anxiety on the EPM than males (p < 0.001) by spending more time in the open than the closed arms. While in the marble-burying task, females displayed less perseveration than males (p = 0.024). Previous studies in this lab have also indicated a sex difference in the severity of subsequent behavioral deficiencies.
Histologically, there were no statistical significance in brain volume differences between Sham-NS subjects and Sham-SSRI subjects. However, it is interesting to note that with the exception of the dorsal hippocampus, SSRI subjects trended toward larger brain volumes than NS subjects in striatal and limbic structures associated with anxiety. Our findings mirror others which indicate that exposure to SSRIs spurs neurogenesis, creating larger structural volumes related to emotionality [33]. Limitations of this study include, that we were not able to conduct polymerase chain reactions to assess true methylation status; the small sample size for histology and that structural integrity of tissue varied. As such, not all structures related to emotionality were examined; in addition, the number of samples varied per structure.
Conclusion
In conclusion, significant associations between serotonergic anomalies were discovered that have also been associated with chronic stress, and an HI injury typical of preterm/term populations. HI subjects displayed decreased learning and memory and a trend towards increased anxiety. Chronic SSRI treatment showed to be protective with a trend of faster learning. Thus, it appears that physiological adaptations to HI, by altered serotonin expression, may be beneficial. Further research might elucidate whether chronic SSRI can exert some neuroprotection against hypoxic-ischemic insults.
Footnotes
Acknowledgments
We wish to thank Dr. Roslyn H. Fitch and the Fitch Lab for providing the space, animals and lab equipment to conduct this study. We wish to thank Fitch Lab students Ruth McLeod and Peter Perrino, MS for their assistance in running behavioral tasks and perfusing the animals.
Disclosures
Supported by the National Institute of Nursing Research of the National Institutes of Health (NIH-NINR) under award number 1F32 NR018591-01 (S. Casasvant) and award number 5R01NR016928-02 (X. Cong), the University of Connecticut Institute for Brain and Cognitive Sciences (S. Casavant), University of Connecticut Research Excellence Award (R.H. Fitch, T. Rosenkrantz).
Animal research statement
The authors affirm that all procedures were approved by the University of Connecticut’s Institutional Animal Care and Use Committee (IACUC).