
Abstract
BACKGROUND:
Intermanual transfer of learning is an important movement basis for a keyboard and instrument playing movement. However, the issue of where neural plastic mechanism occurs in the brain after intermanual transfer training remains both controversial and unresolved.
OBJECTIVE:
The aim of present study is to investigate the neuroplastic mechanism associated with the interlimb transfer learning from non-dominant hand to dominant hand.
METHODS:
Twenty healthy right-handed adults were classified into either the control group (no-training) or the experimental group (training serial button-press motor task, SPMT), 5 days a week for two consecutive weeks. SPMT involved pressing the numbers 1, 2, 3, and 4 in a random sequence, which was presented in the monitor screen. Outcome measures included movement accuracy (MA), movement time (MT), and the fMRI data using a 3T MRI scanner. Repeated measures of analysis of variance (ANOVA) and non-parametric tests were used at p <0.05.
RESULTS:
Motor performances in the MA and MT were significantly more improved in the experimental group than in the control group (p <0.05). Neuroimaging data revealed a distributed subcortical and cortical motor network including the SMA–thalamus (VL/VL)–basal ganglia–cerebellum loop, suggesting a differential and time-dependent neural network utilized during intermanual transfer learning.
CONCLUSION:
Pre-training intermanual transfer learning involved a form of declarative (or explicit) motor learning, which was primarily mediated by the cortical motor network, whereas post-training involved a form of procedural knowledge, which activated subcortical and cortical motor network regions, including the SMA–thalamus (VL/VL)–basal ganglia–cerebellum loop.
Introduction
Intermanual transfer, such as serial keyboard-pressing, instrument playing and writing, is a phylogenetic learning process in normal motor development and skill acquisition. Intermanual transfer of skill acquisition, also known as simple cross-transfer learning, is defined as when training for a certain task with one hand can be positively carried over to the other hand (Imamizu & Shimojo, 1995). This concept has been explored by behavioral studies that examined the intermanual transfer of learning concepts within various contexts including motor coordination, force, sensation, and functional tool use learning (Charron et al., 1996; Japikse et al., 2003; Schulze et al., 2002). Clinically, an understanding of intermanual transfer of skill acquisition can inform clinical decisions regarding evaluation and motor retraining following interhemispheric/callosal pathology, ipsilesional brain damage, or dominant hand amputation (Jarus, 1994).
Despite these clinical implications, there is a dearth of neuroscientific evidence regarding neural control mechanisms and plasticity associated with intermanual transfer training (Mecklinger, 2002; Taylor & Heilman, 1980). The issue of where such neural plasticity occurs in the brain after intermanual transfer training remains both controversial and unresolved (Schulze et al., 2002). It has been theorized that a dual motor memory (“engram”) is formed in both hemispheres via interhemispheric fibers in the corpus callosum when a novel skill is acquired (Muellbacher et al., 2001; Teixeira & Caminha, 2003). In an attempt to investigate the cortical areas associated with intermanual transfer of skill acquisition, two neuroimaging studies used positron emission tomography (PET). One study was conducted in normal subjects and PET was performed to detect cortical activation areas by means of measuring regional cerebral blood flow (rCBF) while the subjects were instructed to draw figures (Thut et al., 1997b). Intermanual transfer of learning was associated with increased rCBF in the left medial prefrontal cortex and right prefrontal convexity, whereas individual rCBF changes in the right prefrontal cortex were inversely correlated with individual changes in transfer learning performance. This finding suggests the possible existence of dual facilitatory and inhibitory mechanisms, which may modulate intermanual transfer of learning (Thut et al., 1997b). Conversely, the other neuroscience study into primate tool-use skill and transfer of training to the opposite hand revealed increased cortical activation in the bilateral prefrontal cortex, bilateral intraparietal sulci, and cerebellum contralateral to the trained hand (Obayashi et al., 2003). This seemingly implies a facilitatory mechanism mediated by modification of a motor engram specific to the original learning in monkeys (Obayashi et al., 2003). Although both PET studies provided neuroscientific evidence that intermanual transfer of skill acquisition is attributed to the efficiency of interhemispheric modulation and the presence of activation areas, the precise nature of the mechanisms or neural network involved remains unclear (Rauch et al., 1998; Stoddard & Vaid, 1996; Taylor & Heilman, 1980).
Event-related fMRI provides a powerful tool for accurately localizing brain regions in which there are block-by-block neuroplastic changes in intermanual transfer of learning-related activity. This advantageous functional capability in fMRI has important ramifications for differentiating learning-specific changes from non-specific neuroplastic changes (Ramnani, 2006). The purpose of this study is to investigate the enhancement of performance through 2-week intermanual transfer training and demonstrate how the neural network differently reacts in transfer learning from non-dominant to dominant hand via fMRI analysis.
Methods
Subjects
Twenty healthy right-handed adults (10 females, mean age, 23.9 ± 3.5 years) volunteered to participate in this study. Professional typist, musicians were not included. All subjects provided written informed consent prior to their participation. Handedness was determined by the modified Edinburgh inventory (Oldfield, 1971). Subjects were then randomly assigned to either the experimental group (n = 10, mean age, 23.4 ± 4.1 years) or the control group (n = 10, mean age, 24.3 ± 2.9 years). The experimental group received serial finger button-press motor task training whereas the control group did not receive any training.
Motor practice training
The motor practice training involved serial button-press motor task (SPMT) training, which was given 50 minutes/day, 5 days a week for two consecutive weeks. The customized SuperLabPro 2.0 (Cedrus Co., NY, USA) system was used to design the motor practice paradigm. Each button was labeled with a number that represented what finger to press the button with 1, the index finger; 2, the middle finger; 3, the ring finger and 4, the little finger in the keyboard. The computer monitor was placed 50 cm from the subject who was comfortably seated in a chair. The square shape buttons with 7-digit number random combination of 1, 2, 3 or 4 were arranged horizontally on the monitor for 3 seconds. The subject was then instructed to repeatedly press the four corresponding buttons as accurately and quickly as possible using the non-dominant (left) hand within 3 seconds. The subjects performed 10 repetitive pressing tasks within 30 seconds, which is defined as ‘task block’. The subjects were then instructed to repeat 8 task blocks, which is defined as ‘training run’ (a total of 560 presses) (Fig. 1(b)). The resting interval was provided for 15 seconds after each task block. None of subjects reported feeling uncomfortable and every subject completed motor practice training during experimental period without any dropout. To control head-movement artifacts during fMRI scanning, a strap was used on forehead of the subject.
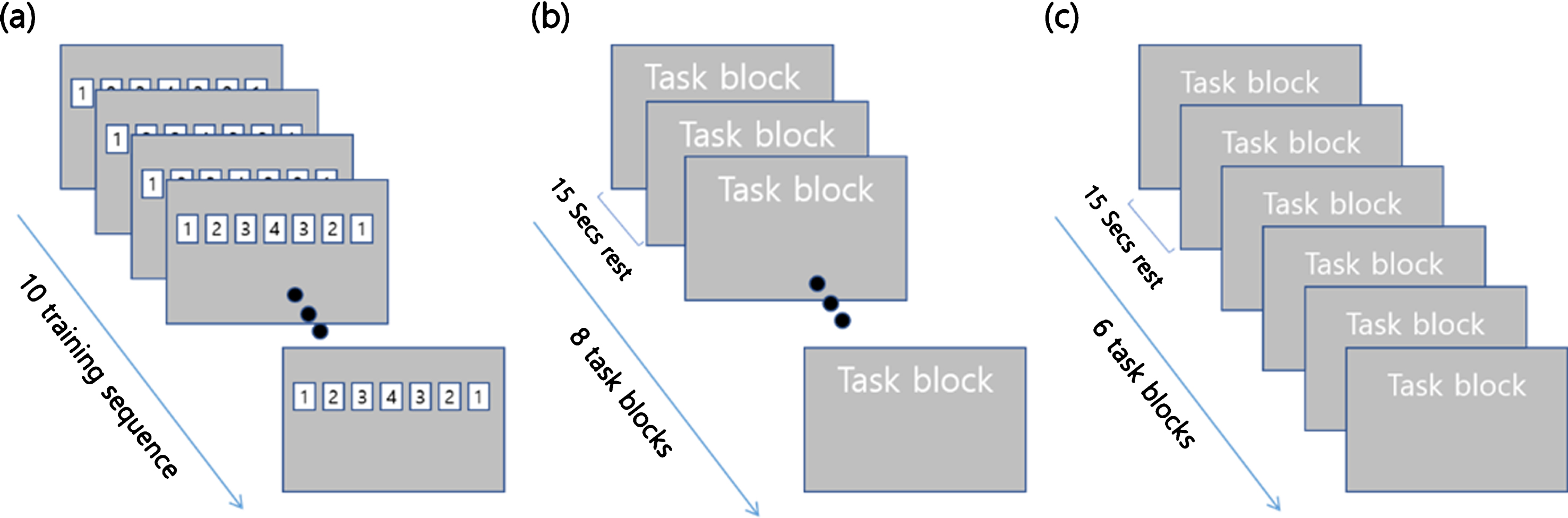
Composition of (a) task block, (b) training run, (c) measurement.
Motor learning performance was assessed by recording movement accuracy (MA) and movement time (MT) with using SuperLabPro 2.0 software in the MRI scanner environment. MA was defined as the total number of correct button-press responses. MT represented the elapsed time from visual stimulation displayed on the monitor to the time to complete a sequential finger pressing motor task. Data were collected three times: pre-training, at 1 week post-training, and 2 weeks post-training (after 5 and 10 training days). Subjects performed task blocks for 6 times with 15 seconds interval rest (a total of 420 presses) (Fig. 1(c)). E-Prime (Psychology Software Tools, USA) and an Integrated Functional Imaging System (IFIS; Invivo, USA) were used to present the stimuli and record behavioral data.
fMRI Data Acquisition
fMRI was performed on a 3T ISOL Forte scanner (ISOL, Rep. of Korea) at baseline, 1 week, and 2 weeks after sequential motor practice for all subjects. In the functional scan, we obtained two single-shot echo-planar imaging (EPI) runs based on the blood oxygenation level-dependent (BOLD) technique. Imaging parameters consisted of 85 volumes of the whole brain (TR/TE = 3000/30 ms, Flip angle 60°, FOV 220 mm, 64'64 matrix, slice thickness 4 mm). Each EPI run included 6 alternating rest and task blocks and 5 dummy scans. A T1-weighted anatomical scan was acquired for each of the subjects (3D MPGR, TR/TE = 11/5.7 ms, flip angle = 10o FOV = 220, matrix = 224 × 256, slice thickness 1 mm). Two scanning runs were acquired during each fMRI session with either the left or right hand in a random order. Each functional run consisted of 125 images including 5 dummy scans at the beginning of each functional run to decrease the effect of non-steady-state longitudinal magnetization. After the functional scan, T1-weighted anatomic images were acquired in transaxial planes parallel to the anterior commissure-posterior commissure line (AC-PC line). The standard fMRI data acquisition procedure was implemented over repeated measurements.
For the functional neuroimaging paradigm, all subjects were secured in the MRI scanner using foam padding to limit head motion within the coil. The MRI-compatible SuperLab Pro 2.0 system was used to project a repeated cycle of 15 seconds of rest and 30 seconds of task stimuli onto the LCD visual display of the Integrated Functional Imaging System (IFIS, MRI Devices Co., USA). As with the motor practice paradigm, the subject was instructed to perform the SPMT task presented for 3 seconds. During the task block, subjects pressed the representative response buttons horizontally arranged on the non-magnetic response box repeatedly and as quickly as possible for 30 seconds. MT and MA were concurrently recorded.
Statistical analysis
Behavioral data analysis
Statistical analysis included mean and standard deviation, repeated measures of analysis of variance (ANOVA), and non-parametric tests. Repeated measures ANOVA was conducted to determine any significant differences (inter-manual transfer effect) in the MT and MA scores of both hands among pre-training, 1 week post-training, and 2 weeks post-training time points between the control and experimental groups. The Wilcoxon test was used to compare MT and MA scores across the three time points. The Mann-Whitney test was used to compare MT and MA scores across the three time points between the two groups. All statistical significance was set at p <0.05. For statistical analysis of the data, SPSS (IBM Corp., Armonk, NY, USA), version 20.0 for Windows, was used.
fMRI data analysis
All individual functional imaging data were realigned, coregistered, normalized, smoothed, and then statistically analyzed using SPM-99 software (Wellcome Department of Cognitive Neurology, University College of London). Brain activation in the experimental and control groups was analyzed using random effects group analysis of both hands. Two-way ANOVA was used to analyze one between-subject factor (group) and two within-subject factors (hand side and training). All clusters with more than 3 contiguous voxels were considered significant with an individual voxel height threshold of p <0.001 (uncorrected) with a cluster-level threshold of Pb0.05 (False Discovery Rate [FDR] corrected) unless otherwise observed.
Results
Behavioral motor performance
The pre-training test revealed similar MA and MT scores between groups, indicating similar baseline motor performance. However, after training, significant differences were observed in MA and MT scores between the experimental and control groups (p = 0.0001). Moreover, in within-subject factor analysis, there were no significant differences in MA (p = 0.334) or MT (p = 0.528) scores between hands in the experimental group, whereas a significant difference was observed between hands in the control group (p = 0.001) (Tables 1 and 2).
Comparison of mean MA scores (standard deviation, SD) (N = 20) (Maximum score = 420)
Comparison of mean MA scores (standard deviation, SD) (N = 20) (Maximum score = 420)
*: p <0.05 on non-parametric Wilcoxon matched pair tests between the left and right hands.; **: p <0.05 on non-parametric Mann-Whitney tests between the experimental and control groups.; 1-ANOVA: one-way ANOVA, r-ANOVA: repeated measures ANOVA.
Comparison of MT scores (ms). Numbers in the cells represent mean (SD)
*: p <0.05 on non-parametric Wilcoxon matched pair tests between the left and right hands.; **: p <0.05 on non-parametric Mann-Whitney tests between the experimental and control groups.; 1-ANOVA: one-way ANOVA, r-ANOVA: repeated measures ANOVA.
One-way ANOVA revealed significant improvements in the MA and MT scores of the left hand in the control group as well as both hands in the experimental group (p <0.05) (Figs. 2A and 2B). The Wilcoxon test showed significant differences in MA and MT scores between pre-training, 1 week post-training, and 2 weeks post-training time points with the exception of MA score between the pre-training and 1 week post-training periods. The Mann-Whitney test revealed a significant difference between groups, with the experimental group showing greater performance in both MA and MT scores as a function of the intervention (p <0.05).
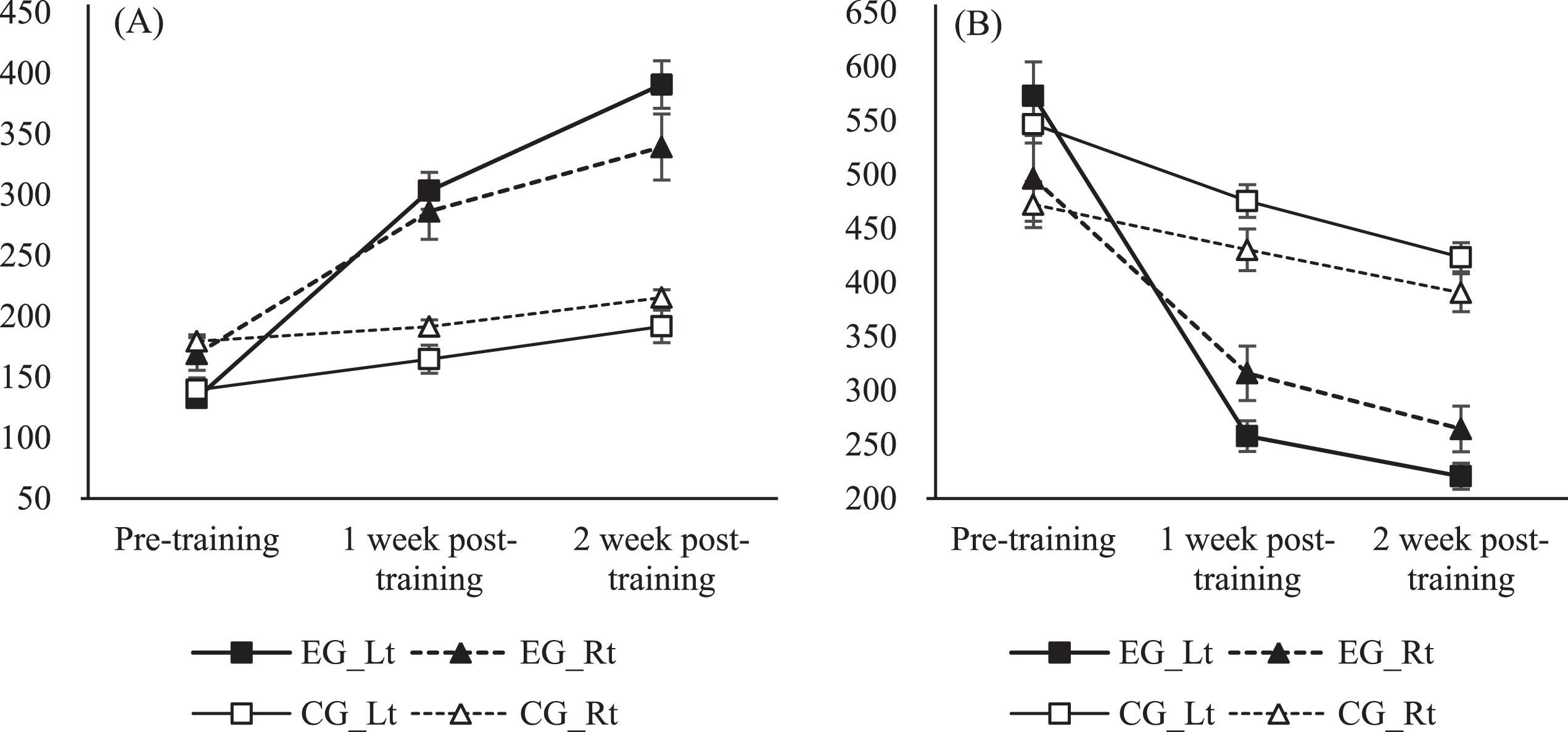
Changes in (A) movement accuracy (MA), (B) movement time (MT) scores pre-training and 1 and 2 weeks post-training. MA scores improved significantly in both hands in the experimental group and the left hand of the control group. MT score was reduced significantly in both hands in the training and control groups. On repeated measures ANOVA, changes in MA and MT scores in the experimental group were significantly different from those in the control group.
Group comparison between pre-and post-training periods
Baseline (pre-training) fMRI data were comparable between control and experimental groups. Specifically, cortical activation areas included the contralateral primary sensorimotor cortex (SM1), bilateral pre-motor cortices (PMC), supplementary motor areas (SMAs), bilateral secondary sensory areas (S2), left dorsolateral prefrontal cortex (DLPFC), bilateral parietal cortices, and bilateral cerebellum during left hand SPMT. In the experimental group 1 week post-training, all activation in these areas had decreased, with the exception of relatively increased activation in the SMA and bilateral cerebellum. Furthermore, the DLPFC area was completely deactivated. In the control group 1 week post-training, activation in most of the previously activated areas had decreased, but to a much lesser extent when compared to the experimental group who underwent left hand SPMT. However, during right hand SPMT, the experimental group demonstrated activation in similar areas to left hand SPMT, which encompassed the contralateral SM1, bilateral PMC, SMA, parietal cortices, and right cerebellum in pre-training fMRI data. After post-training, the medial cerebellar area was predominantly activated along with new activation of the contralateral cerebellum, whereas the SM1, PMC, and parietal cortices were deactivated. However, in the control group, the medial and ipsilateral cerebellar areas were activated in the pre-training test, with increased activation on the post-training test and additional activation of the contralateral cerebellum (Fig. 3).
Brain activation upon sequential finger movement of the trained (left) and untrained (right) hands in the experimental (n = 10) and control groups (n = 10) pre-training and 2 weeks post-training (uncorrected p <0.001).
Left hand SPMT training significantly increased cortical activation in the right DLPFC, SMA, temporooccipital junction, left post-central sulcus, and left cerebellum during left hand serial button-press test in the experimental group. However, the left posterior parietal lobe, bilateral fusiform gyri, left parahippocampal gyrus, and bilateral thalamus were relatively less activated (Table 3, Fig. 4).

A and B represent brain fMRI images related to the group main effects of sequential motor training (uncorrected p <0.001). (A) Areas more activated in the experimental group compared to the control group upon movement of the trained (left) hand. (B) Areas less activated in the experimental group compared to the control group upon movement of the trained (left) hand. Cbll, cerebellum; TO, temporooccipital region; PFC, prefrontal cortex; PCG, precentral cortex; SMA, supplementary motor area; PHG, parahippocampal gyrus; Th, thalamus; BG, basal ganglia; M1, primary motor cortex; PPC, posterior parietal cortex.
Brain areas activated by sequential finger movement of the left hand in the experimental and control groups before and after 2 weeks of training
Left hand SPMT training increased cortical activation significantly more in the right SMA, medial frontal region, and bilateral insula during the right hand serial button-press test in the experimental group compared to controls. However, the bilateral thalamus bilateral parahippocampal gyri, left posterior parietal and right inferior parietal regions were less activated (Table 4, Fig. 5).
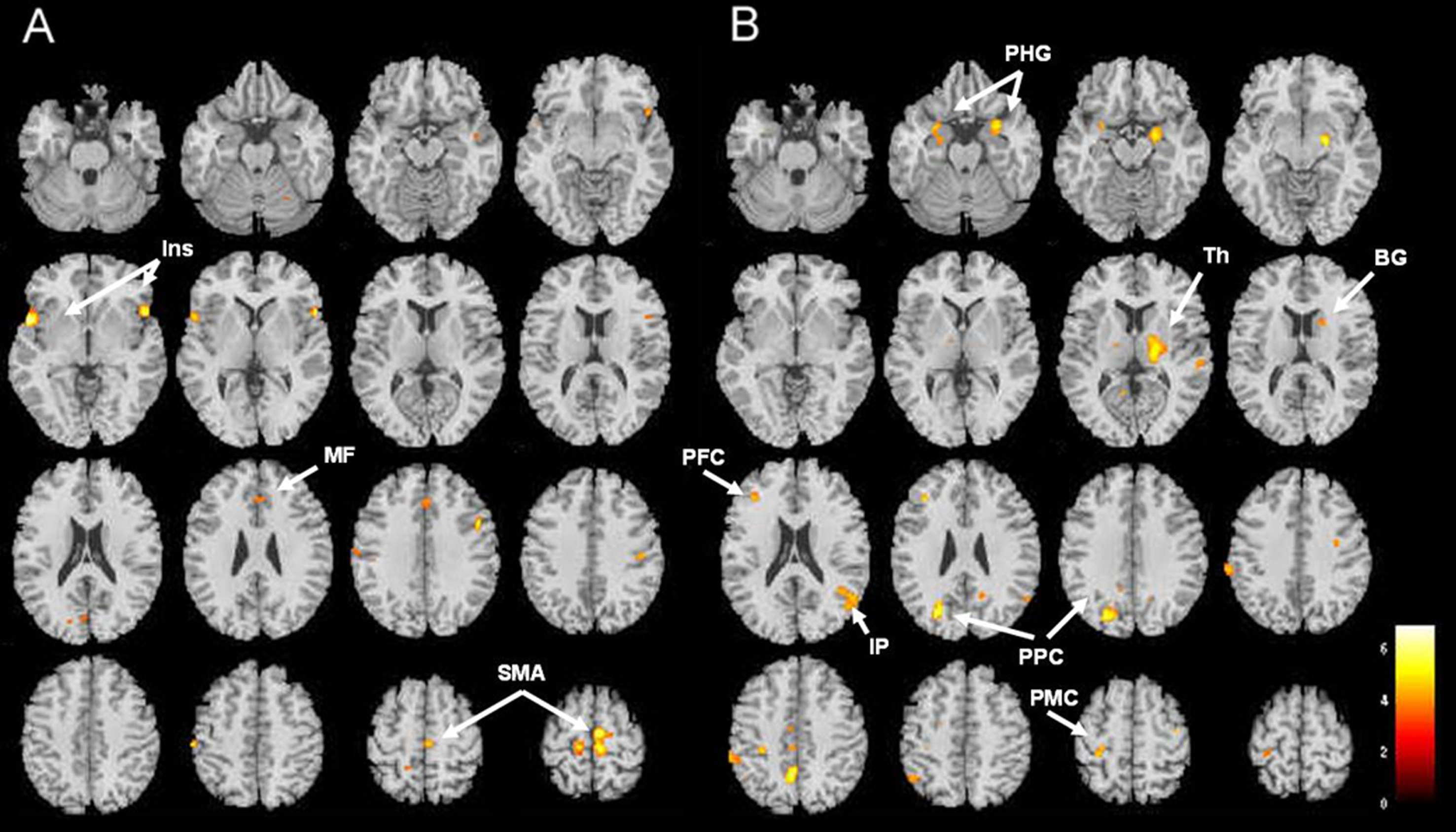
A and B illustrate brain fMRI images related to intermanual transfer of training (uncorrected p <0.001). (A) Areas more activated in the experimental group compared to the control group upon movement of untrained (right) hand. (B) Areas less activated in the experimental group compared to the control group upon movement of untrained (right) hand. Ins, insula; MF, medial frontal cortex; SMA, supplementary motor area; PHG, parahippocampal gyrus; Th, thalamus; BG, basal ganglia; IP, inferior parietal cortex; PPC, posterior parietal cortex; PMC, pre-motor cortex.
Brain areas activated by sequential finger movement of the right hand in the experimental and control groups before and after 2 weeks of training
Investigation of non-dominant hand training-induced intermanual transfer effects and the involved neuroplastic mechanisms in the sensorimotor network has important theoretical and clinical implications. The present study supports evidences of non-dominant hand training-induced intermanual transfer effects and associated cortical activation in the sensorimotor network. To examine intermanual transfer effects from non-dominant hand to dominant hand, we investigated non-dominant-hand training-induced fMRI activation during SPMT. As predicted, intermanual transfer influenced the sensorimotor network as evidenced by fMRI data and behavioral motor changes.
Interlimb transfer training-induced activation was associated with areas such as the contralateral SMA, cerebellum, thalamus, putamen, and contralateral S2, which became less activated, while the ipsilateral cerebellum, left DLPFC, fornix, and right inferior frontal gyrus (IFG) became more activated. Such cortical plastic changes were noted during the interlimb transfer training phase in previous fMRI studies (Halsband & Lange, 2006; Anguera et al., 2007; Mecklinger et al., 2002; Oldfield, 1971; Ramnani, 2006). Anguera et al. investigated sensorimotor adaptations associated with dominant-to-non-dominant hand intermanual transfer tasks in 25 healthy adults and demonstrated activation in the bilateral middle frontal gyrus, right IFG, right pre-SMA, and right inferior parietal lobule and bilateral precuneus (Anguera, 2007). Similarly, Obayashi et al. demonstrated fronto-parieto-cerebellar network interactions (i.e., PFC and cerebellum) underlying intermanual transfer of learning associated with tool-use between hands by means of detecting regional cerebral blood flow in two monkeys on PET (Obayashi et al., 2003). Cerebellar projections to the contralateral dorsal PFC may be involved in the cognitive procedural motor learning process, which utilizes prior-practiced memory (Obayashi et al., 2003) and may mediate intermanual transfer learning by modifying prior memory. Moreover, DLPFC activation may play an important role in accessing stored motor cues or searching memories, which could facilitate transfer learning and performance of the opposite hand by means of interhemispheric interaction (Halsband & Lange, 2006). In fact, Sakai and colleagues documented that the DLPFC and frontal pre-motor areas can process interlimb transfer of learning more efficiently by accessing and retrieving stored motor program memories during sequential finger movement tasks (Sakai et al., 2002). Additionally, cortical activation of the right IFG was identified during the transfer learning process, which confirms the role of the IFG in intermanual transfer learning in executing sequential movement. Recent neuroimaging studies reported that the right IFG was important in grasping and hand motor sequence learning (Grafton et al., 2002; Johnson et al., 2003; Koski et al., 2002; Mecklinger et al., 2002).
On transfer, cortical activation was primarily observed in the M1, SMA, thalamus, basal ganglia, and cerebellum. Our fMRI results demonstrated that the magnitude of intermanual transfer of SPMT was associated with both the SMA and VLa thalamic nucleus areas. SMA has strong transcallosal commissural networks with both the contralateral M1 and pre-motor areas. Empirical research from primates revealed that the internal segment of the globus pallidus in the basal ganglia thalamus releases GABAergic neurotransmitters to the VLa nucleus, which is correlated with increased magnitude of intermanual transfer learning. This association of VLa and SMA activation was well established in successful intermanual transfer elsewhere, which confirmed the associated neuronal role for the VLa–SMA–basal ganglia network in intermanual transfer learning of procedural knowledge (Perez et al., 2007). It is plausible that the thalamic VLa nucleus modulates the intermanual transfer learning process while interacting with the cortical (SMA, M1) and subcortical targets (basal ganglia, and cerebellum) in both cerebral hemispheres in the selection of correct motor responses to visual cues. On the other hand, the VLa, which receives strong afferent input from the cerebellum regulates the adjustment process via motor error estimation and correction of intermanual transfer learning of procedural knowledge, which was evidenced by post-training fMRI correlation with transfer in the current study. A recent Resting-state (rs)-fMRI and diffusion weighted imaging (DWI) research demonstrated increased cortical activation in bilateral supplementary motor areas (SMAs) after unilateral ballistic wrist flexion training. Such resting neural activation was correlated with more improved performance of the untrained limb (83.6%) when compared with that observed for the trained limb as well as that of a control group (no training). The finding was consistent with the present neuroimaging data which showed increased interhemispheric interactions between bilateral SMA, which further support an important neuromotor control role in intermannual transfer (Ruddy et al., 2017).
A couple of study limitations should be considered in future research. The present results support the dominant intermanual transfer effects for rehabilitation in a neurological (e.g., hemiparetic hand) or sport context (e.g., athletes with arm immobilization). However, further investigation should be implemented to explore the role of task complexity and attention in relation to skill level in pathological populations as mentioned above. The other limitation is that it may be of great interest to compare an active intramanual transfer training mode with a passive observational feedback intramanual transfer training mode (Reissig et al., 2015) or active left hand, motor imagery, and bilateral training or left-handed participants have not been examined in the present study. In this line, a higher sample size is needed to examine potential high variability of skill levels due to baseline characteristics and intermanual performance gained through the active dominant intramanual transfer training.
Conclusion
The present investigation provided an underlying neurophysiological mechanism for the involvement of cortical and subcortical motor control networks utilized during pre-training and post-training intermanual transfer learning of the SPMT task. Pre-training intermanual transfer learning involved a form of declarative (or explicit) motor learning, which was primarily mediated by the cortical motor network, whereas post-training involved a form of procedural knowledge, which activated subcortical and cortical motor network regions, including the SMA–thalamus (VL/VL)–basal ganglia–cerebellum loop. Neuroimaging results also suggested that differential and time-dependent neural neuroplasticity was involved during intermanual transfer learning phases of the SPMT task. Finally, the current evidence contributes important knowledge regarding the neuroplastic mechanisms associated with intermanual transfer learning of the SPMT task in healthy humans, which is used to apply sequential hand motor skills acquired with the dominant hand to the contralateral hand during daily life activities.
Author contributions
All authors contributed to the conceptual framework of the study, experimental design, data collection, analysis, and manuscript preparation. Dohee Jung and Ji-Won Park participated as co-first authors. Yun-Hee Kim and Joshua H. You participated equally as corresponding authors.
Conflict of interest
The authors declare that there is no competing or conflict of interests regarding the publication of this paper.
Footnotes
Acknowledgements
This study was supported by grants from the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIP) (NRF-2017R1A2A1A05000730).