
Abstract
BACKGROUND:
Action observation describes a concept where the subsequent motor behavior of an individual can be modulated though observing an action. This occurs through the activation of neurons in the action observation network, acting on a variety of motor learning processes. This network has been proven highly useful in the rehabilitation of patients with acquired brain injury, placing “action observation” as one of the most effective techniques for motor recovery in physical neurorehabilitation.
OBJECTIVE:
The aim of this paper is to define an EEG marker for motor learning, guided through observation.
METHODS:
Healthy subjects (n = 41) participated voluntarily for this research. They were asked to repeat an unknown motor behavior, immediately after observing a video. During the observation, EEG raw signals where collected with a portable EEG and the results were later compared with success and fail on repeating the motor procedure. The comparison was then analyzed with the Mann-Whitney U test for non-parametrical data, with a confidence interval of 95%.
RESULTS:
A significant relation between motor performance and neural activity was found for Alpha (p = 0,0149) and Gamma (0,0005) oscillatory patterns.
CONCLUSION:
Gamma oscillations with frequencies between 41 and 49,75 Hz, seem to be an adequate EEG marker for motor performance guided through the action observation network. The technology used for this paper is easy to use, low-cost and presents valid measurements for the recommended oscillatory frequencies, implying a possible use on rehabilitation, by collecting data in real-time during therapeutic interventions and assessments.
Introduction
Even though sensorimotor learning is often associated with repetitive training of motor procedures, there has been a vast group of studies on this topic analyzing alternative methods. These methods describe in general, ways of training motor function by facilitating the systems involved without excessive repetition, increasing efficiency at the same time as resources can be spared (Bassolino et al., 2014). For neurorehabilitation this means an increased focus on cognitive functions related to sensorimotor (re-) learning after injury, leading to reduced recovery time, which also means a decrease of the risk of comorbidities for these patients.
A solid example of that can be found in the work of Seidler et al. (2012), where the authors integrated spatial aspects of working memory to improve sensorimotor adaptability and sequential procedural learning in order to recover motor function in acquired brain injury patients. The authors also discussed an overlapping between spatiality in working memory and the early phases of motor adaptation in relation to the activation of the dorsolateral prefrontal cortex.
The integration or training of cognitive abilities, to improve sensorimotor function in the rehabilitation of neurologic patients as a therapeutic approach is called mental training (Johansson, 2011). This concept can be subdivided in different techniques, according to the main systems involved in facilitating learning, one of them being a technique called action observation. The physiological base of learning through observation is not new and describes that the observation of a motor procedure can influence the motor behavior of a passive observer and, thereby, increase the efficiency and rate of motor learning processes (Cross et al., 2009; Douma et al., 2015).
The physiological system involved in this process was first described with its neuroanatomical components by Rizzolatti and colleagues in 1992 and was named “mirror-neuron system”, but their observations were based on studies with non-human primates. In 2001, Buccino et al. were able to describe a similar system in humans as well, opening the debate not only about the similarities, but also the differences in facilitating motor learning the human brain and other primates.
The theory of the mirror neuron system and its components have been pointed as “weak” in describing this phenomenon in humans (Turella et al., 2009) and a new model, called “action observation network” started taking place, adding new functional pathways to its description (Moriuchi et al., 2014; Nummenmaa et al., 2014).
Spontaneous and task-related electroencephalography studies have shown, that brain oscillations in the gamma frequency band (31 –200 Hz) can be found in almost all neocortical sites and subcortical structures under somatosensory stimulation (Uhlhaas et al., 2008). This modality of brain wave activation can be observed in a variety of higher cognitive processes, involving perception and information analysis. In processes involving attention, working memory and the planning of a motor action, these waves showed a similar function as observed by alpha frequencies (8 –12 Hz) in the selection of the information intake, inhibiting distractors (Lachaux et al., 2007).
The facilitation of the communication between distant cortical areas in perceptual processes had already been discussed by Jensen and Maraheri (2010) through the observation of elevated cortical excitability and gamma band synchronization in a task-related context. Hermann et al. (2015) also described the event related synchronization of gamma frequencies being independent on the type of sensory stimulation and primary kinetic state of the subject, implying the importance of these waves in processes of communication between cortical areas during input analysis.
The link between gamma activity, especially between 30 and 70 Hz, and complex cognitive processes of signal analysis in humans and non-human primates had already been mentioned in a study by Fries et al. in 2001 and confirmed in 2013 by Saxena et al. on the analysis of unknown objects, through the visual and tactile systems. This shows the potential of these waves as EEG markers for perceptual processes initiated by observation like the ones taking place in the action observation network.
The use of EEG data can provide evidence about neural processes, which can give therapists and patients a valuable feedback on performance. As described by Daly and Wolpaw (2008), with the use of portable and affordable equipment (software and hardware), so as a better understanding about potentials and deficits of people with sensorimotor impairment and the respective neural networks responsible for those, real time neurofeedback can be a valuable tool in neurorehabilitation.
The intent of this work is to provide data about the importance of gamma oscillations, especially in the range between 41 to 49 Hz, in monitoring neural processes during motor learning, as a marker for predicting the success of sensorimotor performance in a therapeutic context.
Methods
Population
Forty-five healthy participants, students of the second, third or fourth year of a Physiotherapy program in Munich (Germany), in ages between 18 and 35 participated on this project. All had the same level of education and no diagnosed sensorimotor or cognitive deficit. A mental-task-rotation was done to evaluate the spatial reasoning ability in the population, in order to exclude subjects significantly below or above the population average.
Procedure
The participants were asked to observe an instructional video on how to construct a three-dimensional-puzzle, according to the correct order of components. During the observation of the video, raw EEG patterns where registered, using a portable, one-channel EEG, with dry electrodes positioned in Fp1 and Fp2, following the international 10–20 system for location of scalp electrodes. The video was presented once, and the participants where located in an isolated room, having only a 140 × 60 cm table in front of them, with the 9 puzzle pieces and a 22,63 zoll monitor on it. They all wore noise suppression headphones during the observation. The video showed in 100 seconds the correct order and positioning of the puzzle pieces. Immediately after that, the participants were asked to put the nine pieces together (action-phase).
Material
The chosen motor procedure was the construction of a three-dimentional-puzzle, since this represents an unknown motor task, reducing the risk of bias. It was shown that experience and procedural memory can influence the neurophysiological process (Schütz-Bosbach & Prinz, 2007), and by that matter, the neurological pathway used in a context of action observation (Calvano-Merino et al., 2006; Fiorio et al., 2010; Novembre et al., 2014).
The hardware chosen for the recording of raw-EEG patterns in all frequencies, was the BrainLink® from the company Macrotellect® (Shenzhen, China). This one channel, portable EEG uses TGAM (Think Geartm AM) technology –the same used in most commercial portable EEGs, also employed in research. The effectivity of this technology for research proposes was investigated and found to be equivalent to a P300, not showing significant deviation for frequencies between 7,5 and 49,75 Hz (Chen et al., 2014; Grozea et al., 2011).
Statistical analysis
The raw-EEG data was evaluated first through a comparative analysis, where the separate values for each frequency band were compared with the results of the motor activity. The participants were allocated into two groups, according to their outcome in the execution of the task. For the comparison between the two groups, the Mann-Whitney U test for non-parametrical data was done for each observed frequency band with a level of significance set with an alpha of 0,05%.
Data protection
All participants were voluntaries and had to sign a consent form before joying this study. They were informed that the consent could be retrieved at any point during or even after the research, without the need of plausible explanation. Immediately after the signing of the consent, each acquired an anonymous identification number that would be present in all forms instead of their names. This procedure served the blinding of the researchers involved. No physical nor emotional harm for the participants was expected.
Results
Demographic data
Forty-one healthy participants (25 males, 16 females) took part in this study. They were all voluntaries between 19 and 33 years old, with the average age being 22,55 years old and same educational background with very similar social and financial statuses. None of them presented a physical nor cognitive impairment and they could have retreated their consent at any time during or after the study.
None of the 42 participants were able to complete the motor task –construction of the puzzle –in the given time of 15 minutes, still the results could clearly divide the population into two groups: (1) participants that were able to complete 67 –90% of the task, by having left only 1 to 3 pieces of the puzzle to be completed; and (2) participants that were not able to complete more than 33% of the puzzle. Seventeen participants could be placed in the first group and 24 could be placed in group 2, based on their results (Fig. 1a and 1b). No analysis regarding gender versus result was made, since this was not the purpose of this study.
Following the results the population was divided in two groups: (1) 17 participants that were able to complete 67–90% of the task, by having left only 1 to 3 pieces of the puzzle to be completed; and (2) 24 participants that were not able to complete more than 33% of the puzzle.
The recorded raw-EEG values provided by the TGAM system were arranged in 8 categories according to the frequency, as presented in the Table 1. The analysis showed a direct relation between the ability to reproduce the observed motor behaviour and the neural activity recorded during the observation of the video. Significant results were found for alpha and gamma frequency bands, between 7,5 –11,75 Hz (α) and 41 –49,75 Hz (γ).
Frequency bands in Hz for all measured oscillation patterns, showing the equivalent frequencies between TGAM analyzed data in comparison to EEG data found in literature
Frequency bands in Hz for all measured oscillation patterns, showing the equivalent frequencies between TGAM analyzed data in comparison to EEG data found in literature
In the comparison between groups 1 and 2, a positive relation was present in the alpha frequency band during observation of a motor task and motor outcome. This relation seems to be stronger when considering low-wave activity –between 7,5 and 9,95 Hz –then alpha frequencies above 10 Hz. Considering raw-EEG values, group 1 showed an average of 16290 (SD±16035) while group 2 an average of 8593 (SD±7059) (Fig. 2). In a confidence interval of 95%, the Mann-Whitney U test for non-parametrical values showed a p of 0,0149 comparing low-alpha activity between the two groups, and a p of 0,0416 for high-alpha activity (Table 2).
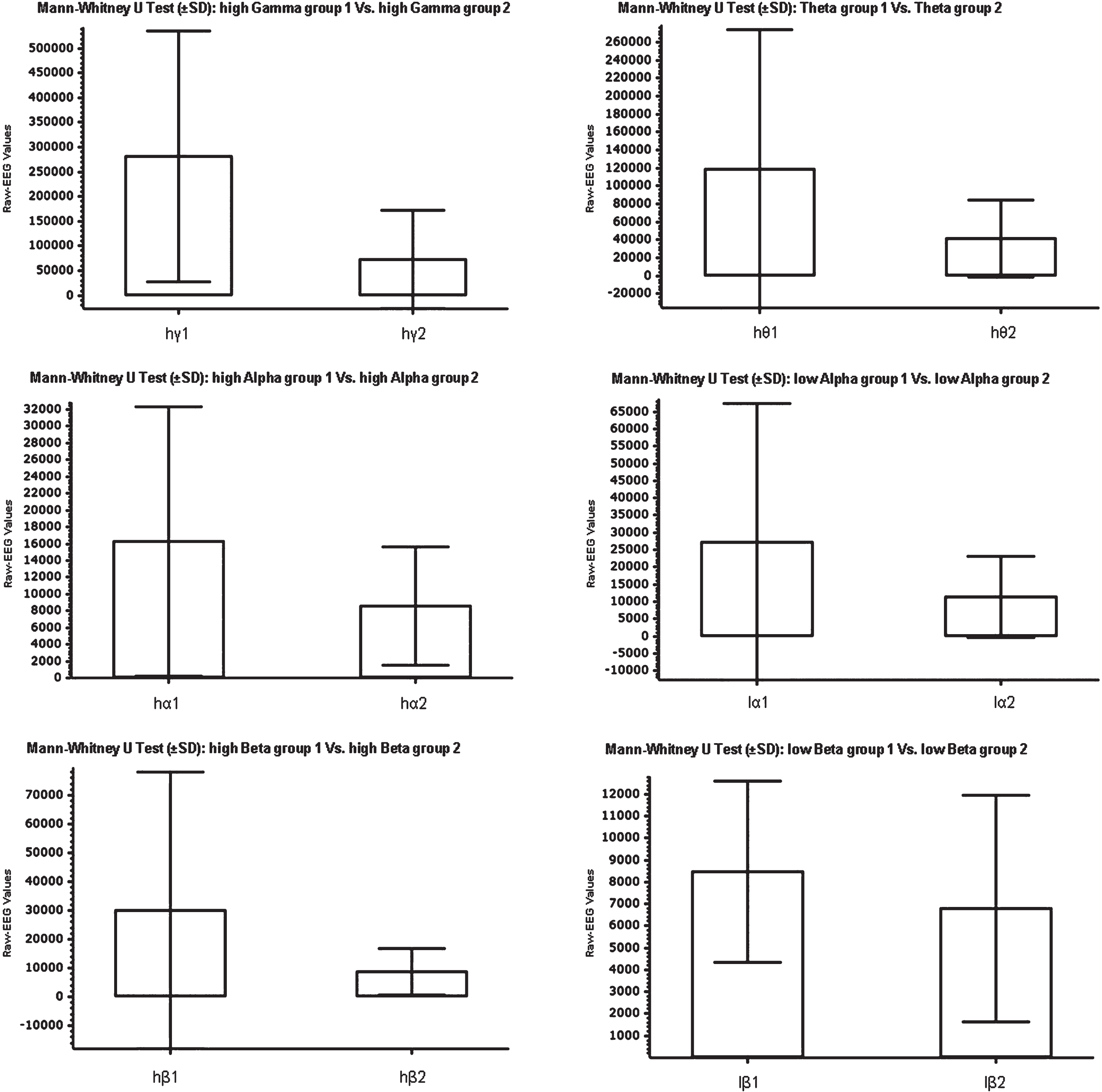
Mann-Whitney U test plot showing the comparison between groups 1 and 2, related to the activity in the Gamma band between 31 und 49,75 Hz (hG), in the Theta frequency band between 3,5 und 6,75 Hz (T), in the Alpha band between 10 und 11,75 Hz (hA), in the Alpha band between 7,5 und 9,25 Hz (lA), in the Beta band between 18 und 29,75 Hz (hB), and in the Beta band between 13 und 16,75 Hz (lB).
Presentation of the p values, calculated with the Mann-Whitney U test for non-parametrical values, for alpha, beta and gamma frequency bands
In the gamma frequency band between 41 –49,75 Hz, the comparison between the two groups showed an even stronger relation between gamma activity during the observation of the task and a positive outcome. Taking the raw-EEG values into account, group 1 showed an average value of 280402 (SD±252981), while group 2 presented an average value of 72227 (SD±99289) (Fig. 2). The p value in this case shows the strongest significance (p = 0,0005) (Table 2).
Other frequency bands (delta, theta, beta)
No significant difference between groups could be observed in delta and beta frequencies. In beta frequency bands between 13 and 16,75 Hz and between 18 and 29,75 Hz, the p values were 0,2136 and 0,1009 (Table 2) considering a confidence interval of 95%. In comparison, theta activity between 3,5 and 6,75 Hz showed a strong correlation –even stronger than the one found in the alpha frequency bands –, with a p of 0,0134. Still, since the applied hardware seems to show a significant deviation in the values for delta and theta oscillations, when compared with the gold standard, these results were not considered for further investigation in this study.
Discussion
The main porpoise of this study was to evaluate the type of neural activation involved in the process of learning though observation, thus making use of the action observation network to acquire knowledge over an unknown motor task. Here, a portable EEG was used to provide information about encephalic oscillation patterns involved in this process. The choice of the device was done based on the criteria of the practicality and accessibility of a portable device (Daly & Wolpaw, 2008) that uses a TGAM technology to produce valid data for investigation (Grozea et al., 2011; Liao et al., 2012; Ahn et al., 2014; Chen et al., 2014).
The role of alpha oscillations in the action observation network
Other authors already considered alpha brand synchronization in the investigation of neural activity over stimulation in the action observation network (Quandt et al., 2013; Muthukumaraswamy et al., 2010). The reason for that is that oscillations are mentioned in literature as being related to processes involving working memory and attention –in specific selective/divided attention –(Linkenkaer-Hansen et al., 2004; Busch & Hermann, 2003). Even though the origin of oscillations in this frequency band is widely spread around the cerebral cortex, as some diencephalic sites, two main sites can be named during visual stimulation during analysis of space and object: the thalamus and visual areas, especially the primary and secondary visual cortices (Ulhaas et al., 2008).
One of the main components of facilitating learning by using the action observation network, is the ability of the subject to investigate and maintain a variable amount of information about space and object for a short period of time. Busch and Hermann (2003) showed that the amount of information given during a working memory task is directly proportional to the activity of alpha oscillations in a range around 10 Hz. In a similar experiment, but with focus on attentional processes, Linkenkaer-Hansen et al. (2004) showed a direct correlation to the demanded attention for a task, thus complexity of the task, and the production of alpha frequencies in the 10 Hz range.
A possible explanation for the distinction of alpha oscillations in these cases, is the inhibitory function of alpha by supressing non-relevant information, so that the cortical pathways involved can be employed for analysis and interpretation of task relevant sensory input (Sauseng et al., 2007). This cohesion between alpha activity and the ability of reproducing a motor procedure immediately after observation was also observed here.
The role of gamma oscillations in the action observation network
A considerable body of studies already confirmed the role of gamma activity in processes involving selective attention, working memory, sensory integration and perception (Hermann et al., 2005; May Tan et al., 2013; Muthukumaraswamy et al., 2010; Jensen & Mazaheri; 2010; Fries et al., 2001). Brain oscillations in the frequency band between 31 and 70 Hz can be found practically everywhere in the neocortex since they are produced by GABA-ergic neurons (Ulhaas et al., 2008).
Many authors discuss the possibility of gamma band oscillations being involved in a neural communication network, in order that distant cortical and subcortical sites can exchange information between and within each other without the need for a structural component (Tan et al., 2013; Muthukumaraswamy et al., 2010).
The results of the present study confirm what anatomical and fMRI studies state about the role of the communication between a great variety of cortical sites in processes of perception, so as their value for motor learning (Buccino et al., 2001). The activity of so called “mirror neurons” in a proposed “mirror neuron system”, presented by Rizzolatti et al. (1996) could explain the cortical activity in non-human primates during processes of motor learning through observation. Still, the same activity in humans was described by many other researchers as being “weak” (Buccino et al., 2001; Turella et al., 2009), implying that more complex structures and pathways, more related to perceptive systems than to the primary and supplementary motor cortices themselves could be involved in motor learning processes, guided through observation, thus the action observation network.
The overlap between cortical sites involved in the execution of movement and perception/interpretation of the same movement has been shown in various fMRI studies (Buccino et al., 2001; Rizzolatti & Craighero, 2004). Also, psychosocial studies support the idea that the attentional and somatosensory components of behaviour observation play a main role in social interaction through movement and posture (Nummenmaa et al., 2014).
These results indicate the existence of a complex neurological system behind the facilitation of movement in a motor learning context, like the one involving the action observation network. This complex system is dependent on the communication and interaction of multiple cortical areas and subcortical sites to stablish a functional neural perceptual system (Nummenmaa et al., 2014; Kreysers et al., 2010; Caspers et al., 2010).
Current literature confirms the importance of the visual system and the activity of gamma oscillatory patterns in motor learning processes that depend on spatial and object recognition, so as sustained and selective attention (Muthukumaraswamy et al., 2010; Lachaux et al., 2007). Also, the theory that gamma activity might form a possible communication network for cortico-cortical and subcortical-cortical sites could be observed in the present study, since the most significant value (p = 0,005) was observed in the frequency range of 41 –49,75 Hz.
The role of beta frequencies in this study
No relation was found regarding beta activity and the motor performance in this work, even though Wrobel (2000) and other authors reported an increase in beta event-related synchronization during visual stimulation and “visual attention”. Besides the consideration that beta activity does not have a significant influence in processes guided through action observation, two other factors could contribute to these results in this study: (1) involuntary movements during the observation of the video and, (2) the stress level of the participants.
All participants were instructed not to move during the observation of the instructional video, however to diminish the number of distractors, they were left alone in the room during this part of the activity. In other observational studies, like from Thompson et al. (2014), it was shown that beta oscillations possess an akinetic aspect, meaning that even though they increase during the planning of a movement, they also show a tendency to decrease during the execution of movement itself. The variance in motor behaviour of the participants (conscious or sub-conscious) during the observation of the video would explain an unpredictable variance in beta synchronization in this phase and therefore, a possible bias for this frequency band in this study.
Another important aspect to consider when discussing the synchronization of beta rhythms is the emotional state of the person. A link between increased beta activity and a negative emotional state was shown in many studies investigating these oscillations (Güntekin et al., 2014; Güntekin & Basar, 2007; Wrobel, 2000). For this matter, it is to expect that again a behavioural discrepancy between participants, this time at an emotional level would produce a bias in this investigation.
Practical use of acquired information
Many portable, low-cost EEG devices, like the BrainLink® (Macrotellect, Shenzhen, China) here used, can provide reliable data for certain aspects applied in mental training. The main goal of the current devices and programs is the training of attention using general markers. With a different setting of the software, in this case to give the user a visual or auditory feedback during gamma activity between 41 and 49,75 Hz, these same devices could eventually serve in the rehabilitation of patients with motor deficits caused by brain damage. For that matter, more feasibility studies would be necessary, and some special considerations should be done regarding a number of conditions, like (1) patients with perceptive disorders or in (2) acute stroke, due to neurophysiological and pathological processes that would create a bias when working with EEG markers. Perceptive disorders: schizophrenic patients –just like patients with other neuropsychological or psychiatric disorders –, present a biodynamic correlation between disfunction and synchronization in gamma band frequencies (Hunt et al., 2017; Demiralp et al., 2007), being the reduction of anterior-posterior coherence between 30–50 Hz the probably most investigated aspect of it. This can be clearly observed by schizophrenic patients in formative tasks (Spencer et al., 2003), where the purpose involves the recognition of objects and faces. Acute stroke: a reduction in the synchronization in all frequency bands can be observed immediately after the decrease in cerebral blood-flow, especially in the first 3 hours after a stroke (Krnjevic et al., 2008). Starting after the first 24 hours, a physiological elevation in delta and theta bands can be observed, followed by a reduction in alpha, beta and gamma bands, ipsilateral to the brain injury (Rabiller et al., 2015; Kim et al., 2014). These discrepancies do not necessarily imply that frequencies in the gamma band would be an inappropriate marker, but their expression (expected event related values) would need to be adapted according to the population.
Limitations of this study
Brain oscillations in delta and theta frequency bands –0,5 to 2,27 Hz and 3,5 to 6,75 Hz in the TGAM, respectively –were not considered for this work, even though theta do play a major role in working memory and attentional tasks, as shown in many studies (Hoffmann et al., 2015; Canolty et al., 2009; Buzsaki, 2002). Their “exclusion” in the data analysis here is due to the fact that the applied hardware in this study shows a significant deviation for these two frequency bands, when compared to the gold standard. Nevertheless, a tendency for theta was observed (p = 0,0134) and a deeper investigation of the relationship between gamma and theta in this context could be promising.
Due to the use of a one-channel EEG, information regarding origin and more specific neuroanatomical sites could not be retrieved. The akinetic role of beta oscillations in this type of observational task, involving perception and interpretation, and their role in the sensorimotor rhythm could be better defined and even show a –maybe weak, but still existent –relation between action observation and beta band activity, like non-human primates show.
Conclusion
The focus of this study was to purpose an EEG marker that would predict the accomplishment of a motor task guided through action observation, using a portable and accessible hardware. Here, gamma oscillations between 41 and 49,75 Hz proved to be a good alternative for a marker, showing the highest level of significance (p = 0,005) when comparing the performance of two groups immediately after the observation of a task.
The use of 41 to 49,75 Hz oscillations as an EEG marker for motor learning based on action observation can open new possibilities to integrate neurofeedback methods in the rehabilitation of patients with motor deficits after encephalic injuries.
Also, the use of accessible technologies in neurorehabilitation that can facilitate movement and motor learning is of importance. With the results presented in this paper, the possibility for real-time data collection using a practical and affordable method can assist therapists during interventions and assessments, by allowing these professionals to understand the effect of their approaches and adapting their measures if necessary.
Other acknowledgements were achieved, regarding the action observation network itself and how this network can influence motor behaviour in humans, in contrast to non-human primates. This study provide evidence that support the idea that more complex pathways are involved in human motor behaviour, compared to other primates. This could open a discussion the true value of the investigation of these complex systems in non-human primate subjects as animal models, when applying this knowledge to understand the functioning of human neurofunctional systems and their influence in motor behaviour.
Deeper research, investigating the neural activity during observation and its impacts on human motor behaviour still need to be done in order to obtain more specific data of these processes and the pathways involved. Also, considering metabolic changes affecting the electromagnetic activity of the brain, such as observed in different populations of psychiatric and acute patients, is important before setting specific functional EEG markers to be used in therapy.
Conflict of interest
The authors declare no competing interests.
Footnotes
Acknowledgments
The company Macrotellect® provided the devices used in this project, for the purpose of supporting research, without any monetary, commercial or otherwise profit.