
Abstract
BACKGROUND:
Improvement of functional movements after supratentorial stroke occurs through spontaneous biological recovery and training-induced reorganization of remnant neural networks. The cerebellum, through its connectivity with the cortex, brainstem and spinal cord, is actively engaged in both recovery and reorganization processes within the cognitive and sensorimotor systems. Noninvasive cerebellar stimulation (NiCBS) offers a safe, clinically feasible and potentially effective way to modulate the excitability of spared neural networks and promote movement recovery after supratentorial stroke. NiCBS modulates cerebellar connectivity to the cerebral cortex and brainstem, as well as influences the sensorimotor and frontoparietal networks.
OBJECTIVE:
Our objective was twofold: (a) to conduct a scoping review of studies that employed NiCBS to influence motor recovery and learning in individuals with stroke, and (b) to present a theory-driven framework to inform the use of NiCBS to target distinct stroke-related deficits.
METHODS:
A scoping review of current research up to August 2023 was conducted to determine the effect size of NiCBS effect on movement recovery of upper extremity function, balance, walking and motor learning in humans with stroke.
RESULTS:
Calculated effect sizes were moderate to high, offering promise for improving upper extremity, balance and walking outcomes after stroke. We present a conceptual framework that capitalizes on cognitive-motor specialization of the cerebellum to formulate a synergy between NiCBS and behavioral interventions to target specific movement deficits.
CONCLUSION:
NiCBS enhances recovery of upper extremity impairments, balance and walking after stroke. Physiologically-informed synergies between NiCBS and behavioral interventions have the potential to enhance recovery. Finally, we propose future directions in neurophysiological, behavioral, and clinical research to move NiCBS through the translational pipeline and augment motor recovery after stroke.
Introduction
Stroke is one of the leading causes of disability worldwide. Just alone within the United States, nearly 800,000 people suffer a stroke each year and are left with long-standing disability. Post-stroke motor, sensory and cognitive deficits impair functional activities such as upper extremity function, balance and walking, thus forcing stroke survivors to discontinue over 60% of their daily activities. The physical disability and consequent caregiver burden also generates negative psychological and social outcomes and reduces their quality of life.
The ability to regain the capacity for functional movements (i.e., movement recovery) such as walking and upper extremity function are important patient goals for post-stroke rehabilitation. Movement recovery relies on an interaction of spontaneous biological recovery and behavior-driven reorganization/plasticity in the remnant neural networks. Noninvasive Neuromodulation using transcranial magnetic (TMS), direct current (tDCS) or random noise stimulation (tRNS) can influence both recovery and reorganization to improve functional outcomes (Ahmed et al., 2023; Plow et al., 2016). To date, numerous studies have employed noninvasive brain stimulation to the ipsilesional motor cortex and have yielded variable results (Di Pino et al., 2014). This variability arises from the heterogeneity of lesion location and size, small sample sizes and an attempt to stimulate a damaged corticospinal system with limited residual capacity (Sankarasubramanian et al., 2017). An alternate strategy is to modulate undamaged neural substrates and pathways that play a key role in motor recovery (Di Pino et al., 2014; Kantak et al., 2012). Accordingly, contralesional primary motor and premotor stimulation has been investigated with varying levels of success (Sankarasubramanian et al., 2017). Following a supratentorial stroke, cerebellum is a key node within the anatomically intact neural network that has the potential to impact cognitive and sensorimotor mechanisms of recovery and reorganization (Wathen et al., 2018). Recently, invasive deep brain stimulation of cerebellar dentate nucleus has shown promise in improving motor recovery in animal models and early human studies (Baker et al., 2023). Noninvasive brain stimulation methods are more accessible clinically and have allowed modulation of cerebellar excitability to influence motor performance and learning in awake humans. Noninvasive cerebellar stimulation (NiCBS) therefore opens an exciting and clinically feasible possibility to augment and/or accelerate movement recovery by targeting motor, sensory and cognitive mechanisms.
The overall thesis of this paper is to examine the theory and application of NiCBS as an innovative adjuvant to behavioral approaches for enhancing movement recovery after supratentorial stroke. In the first part of this paper, we discuss the potential role of cerebellum in motor, sensory and cognitive recovery following supratentorial stroke. We review current evidence and effect sizes for NiCBS in improving movement outcomes after supratentorial stroke. We then present a theoretical, mechanisms-driven framework to assist the design of hypothesis-driven interventions combining NiCBS and behavioral training (e.g., practice) to promote recovery. Finally, we identify directions of future research that will advance NiCBS as an adjuvant to behavioral approaches in a way that preferentially harnesses the plasticity in distinct cerebellar pathways to improve specific outcomes. While there have been previous calls to align behavioral interventions with ongoing neuroplastic changes during stroke recovery (Zeiler & Krakauer, 2013), we propose that a synergistic interaction between cerebellar neuromodulation and behavioral interventions may help improve specific motor outcomes, thus taking us a step closer to precision medicine.
The complexity of movement recovery and the role of cerebellum following supratentorial stroke
The behavioral phenotype of supratentorial stroke is complex and includes impairments in the motor, sensory and cognitive systems that often interact with each other during functional actions (Sathian et al., 2011). Within the motor system, multiple deficits such as impaired force production and coordination, abnormal tone, abnormal movement synergies (impaired individuation), poor motor planning and execution contribute to impaired task performance. Post-stroke sensory impairments are common and range from impaired ability to sense touch/proprioception to perceptual deficits (e.g., neglect). Finally, cognitive processes such as executive-function, working-memory and ability to dual-task are often impaired in patients with supratentorial stroke, dependent on lesion location. These motor, cognitive and sensory impairments interact to collectively amplify deficits in activity performance, recovery and participation.
Recovery of functional movements after stroke relies on restoration and reorganization within the motor, sensory and cognitive systems. In the context of this review, we define movement recovery as the capacity to complete a functional task successfully in a manner that is consistently accurate and efficient. This movement recovery after stroke is implemented by both restoring the function in the intact neural structures that were initially inactivated after the episode of stroke (e.g., diaschisis) and reorganization in the remnant neural networks that allow for efficient task performance (Fig. 1). While much attention has been focused on cortical reorganization, emerging evidence suggests that restorative and reorganizational processes after supratentorial stroke occur in cortical, subcortical, and spinal networks. Cerebellum, through its connectivity with cerebral cortex, brainstem and spinal networks, is exceptionally poised to contribute to both restoration and reorganization within the sensorimotor and cognitive systems.
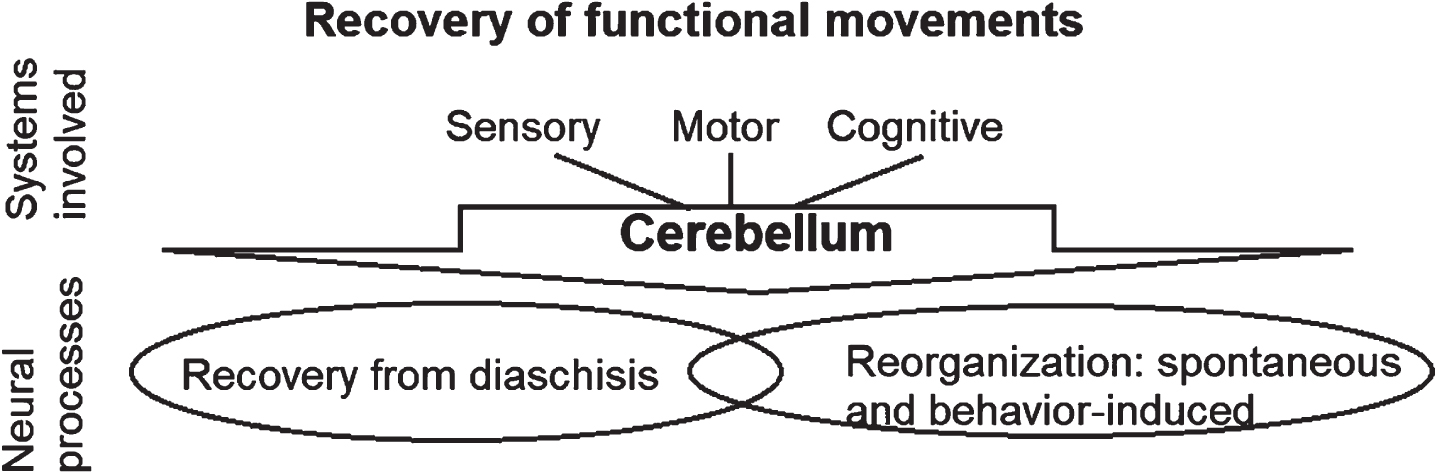
Movement recovery occurs through recovery from diaschisis and reorganization in sensory, motor and cognitive systems. Cerebellum is engaged in both these processes.
Early after supratentorial stroke (∼2–3 days), there is a period of reduced activity in intact neural networks with greatest reduction observed in the perilesional cortex and structures functionally connected to the lesioned brain areas (Rehme et al., 2011; Seitz et al., 1999). For example, there is a reduction in blood flow and metabolism in the contralateral cerebellum putatively due to a functional disconnection of the corticopontocerebellar pathway, a phenomenon described as crossed cerebellar diaschisis (CCD) (Infeld et al., 1995). The extent of early CCD is associated with early motor dysfunction and functional performance in early subacute stage (Takasawa et al., 2002). In a recent study, the volume of lobules IV, VI and VIII-B, all of which share reciprocal connections with frontorparietal and motor circuits, predicted functional independence on a Modified Rankin Scale in supratenotorial stroke survivors (Sadeghihassanabadi et al., 2022). Over the subacute phase (7days- 6months in humans (Bernhardt et al., 2017)), improvements in motor recovery are accompanied by resolution of CCD, although in some cases, diaschisis persists through the chronic phase. Persisting CCD in the chronic phase is thought to be mediated by transsynaptic degeneration of corticocerebellar and cerebellocortical pathways rather than secondary to the decreased perfusion. In chronic stage, the degree of CCD is associated with the severity of neurologic and behavioral deficits (Chang et al., 2017; De Reuck et al., 1997; Serrati et al., 1994). Collectively, these data suggest that cerebellum plays a key role in spontaneous recovery, and that the resolution of CCD is a potent mechanistic target to promote restorative plasticity for recovery.
Concurrently with mechanisms of recovery from diaschisis, reorganization of the remaining neural networks evolves as early as ten days post-stroke and continues into the chronic phases (Grefkes & Ward, 2014). The engagement of the cerebellum in network reorganization is evidenced by functional imaging studies of patients with supratentorial stroke. The cerebellum shows increased activity during goal-directed movements suggesting its engagement in residual motor performance. Longitudinal studies demonstrate increased resting state connectivity between ipsilesional motor cortex and contralesional cerebellum as motor recovery progresses (Park et al., 2011; Tombari et al., 2004; Wang et al., 2010). Finally, experience dependent changes in neural networks with behavioral interventions such as task practice also engage the cerebellum (Puh et al., 2007; Tombari et al., 2004; Várkuti et al., 2013; Ward et al., 2003). Johansen-Berg and colleagues reported a significant correlation between increased fMRI signal change in the cerebellum and improved motor function (Johansen-Berg et al., 2002). Thus, mounting evidence points to cerebellar engagement in reorganization of remnant neural networks to promote movement recovery after stroke. While functional activation of cerebellum associated with motor recovery provides correlational evidence, fundamental research in animal models provides stronger support for the critical role of cerebellum in movement recovery after stroke. In particular, deep brain stimulation and epidural stimulation of the cerebellum after stroke in animal models modulate cortical dynamics and improve motor outcomes (Abbasi et al., 2021; Cooperrider et al., 2014). A detailed review of fundamental research in animal models is beyond the scope of this paper; and can be found elsewhere (Cooperrider et al., 2020).
Given the evidence for cerebellar engagement in spontaneous recovery and reorganization, modulating cerebellar excitability may have therapeutic value. In the subsequent section, we review and summarize the current state of literature focused on noninvasive neuromodulation of cerebellum and its effects on motor performance and recovery after supratentorial stroke.
The growing evidence for the role of cerebellum in motor recovery is strongly supported by recent studies that have applied deep cerebellar stimulation as adjunct to behavioral interventions. A recent non-randomized phase I trial demonstrated that invasive deep brain stimulation to the cerebellar dentate nucleus combined with physical rehabilitation led to clinically significant (median 7 point) change in Fugl-Meyer score in a sample of 12 individuals with stroke who had moderate-to-severe impairment (Baker et al., 2023). These promising results provide further evidence to support the role of cerebellum in post-stroke recovery. Because noninvasive stimulation techniques are more clinically accessible and potentially have lower risks than invasive deep brain stimulation, there have been an increasing number of proof-of-concept studies of NiCBS as an adjunct to behavioral interventions such as motor practice, physical and occupational therapy over the past few years. Here, we present a scoping review to synthesize the current evidence and determine the strength of the effect and parameters of NiCBS used to improve movement recovery in humans with stroke.
Methods
For this paper, we reviewed studies that (a) included individuals over 18 years of age with supratentorial stroke (ischemic or hemorrhagic) regardless of sex, duration and severity of initial impairment, (b) used non-invasive cerebellar stimulation (rTMS, tDCS, TBS or cerebellum-motor cortex PAS), and (c) included motor performance or clinical measures of motor recovery as outcome measures. We excluded studies (a) with individuals with infratentorial stroke (e.g., cerebellar infarction) or other neurological deficits (e.g., Parkinson’s disease/multiple sclerosis), (b) that applied invasive stimulation (e.g.,DBS), (c) that did not include motor function, and (d) animal studies. Four scientific databases were systematically searched: MEDLINE (Medical Literature Analyses and Retrieval System Online; through the PubMed interface), EMBASE (Expert medical Database), Cochrane Central Register of Controlled Trials (CENTRAL), DORIS (Database of Research in Stroke) and PEDro (Physiotherapy Evidence Database). The following keywords were used: stroke, cerebellum, non-invasive brain stimulation, transcranial magnetic stimulation, repetitive transcranial magnetic stimulation, transcranial direct current stimulation, theta burst stimulation, motor learning, motor recovery. The search was restricted to peer-reviewed studies in the English language. Retrieved titles and abstracts of the studies were independently assessed by two authors. The flowchart for selection of studies is illustrated in Fig. 2. After reading full-text articles, the authors selected studies according to the inclusion criteria. Any disagreements between the two authors were resolved through discussion and consensus. All quasi-experimental as well as randomized controlled trials (RCTs) published until August 2023 were included in this scoping review. We qualitatively evaluated the included studies (RCTs, single-arm and within-group studies) with the PEDro Scale.
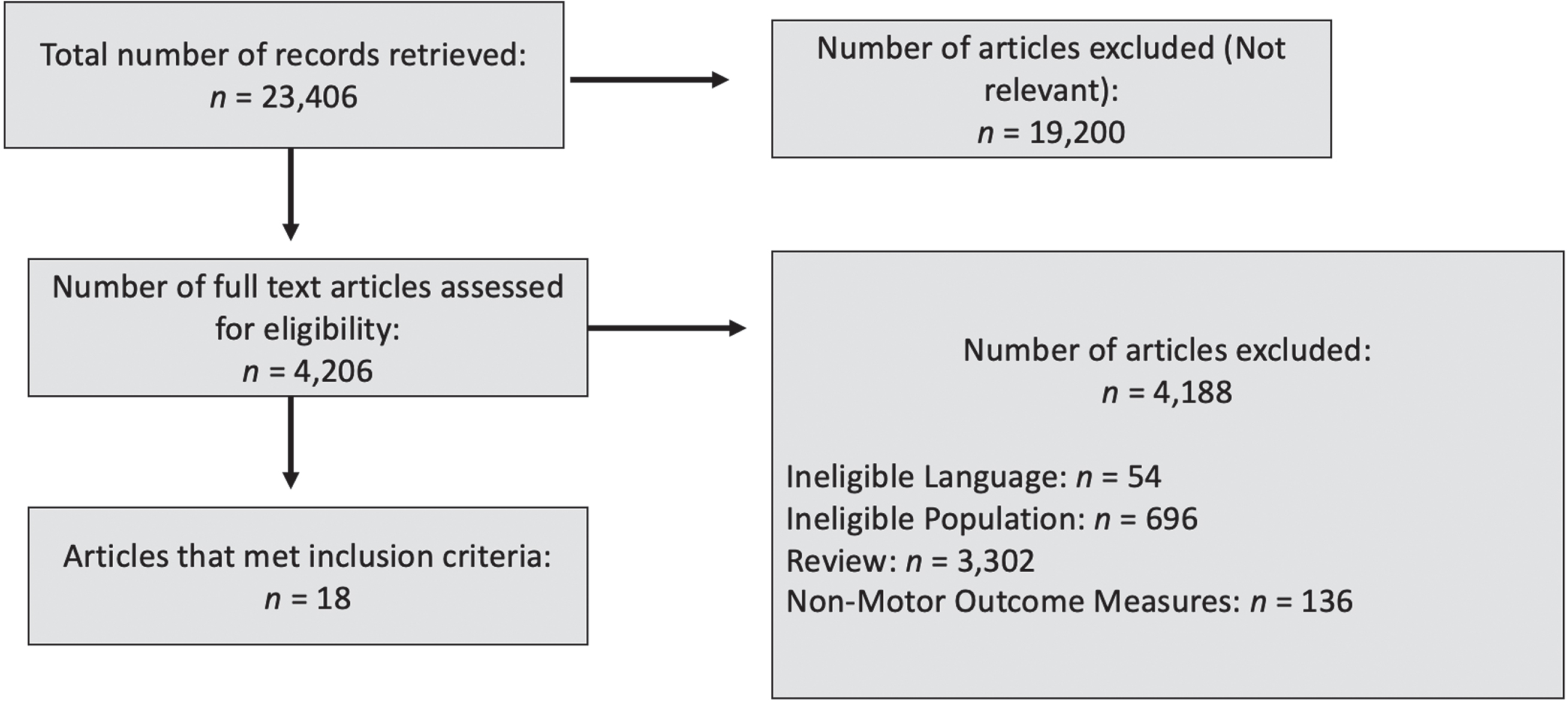
Steps in selection of studies using NiCBS in stroke survivors.

Effect sizes of cerebellar noninvasive brain stimulation and behavioral interventions on upper extremity function, balance and walking outcomes in stroke survivors.
Study characteristics
Each study utilized different NiCBS protocol and two combined stimulation with physical therapy. All studies were designed as RCTs. Outcome measures differed across studies from impairments such as spasticity scores (e.g.,Modified Tardieu Scale) and activity performance (e.g.,Jebsen-Taylor Hand Function Test) to global function (e.g.,Barthel Index).
One study (Chen et al., 2021) used iTBS over the ipsilesional cerebellum, another used PAS over contralesional cerebellum and ipsilesional M1 (Rosso et al., 2022), while the other two (Gong et al., 2023; Wessel et al., 2023) used tDCS 3 cm lateral to the inion, over the cerebellum (ipsilateral to the affected hand, and in the latter over the right cerebellum). Analyses of effect-size for upper extremity impairment and function utilizing iTBS and PAS indicated a large effect size (d = 2.13–2.9) on measures of muscle tone and stiffness (MTS, shear wave velocity) of wrist and elbow flexors. Effect sizes across measures of UE activity performance varied. Two studies found a relatively small effect (JTTr, d = 0.10, movement trajectory, d = 0.02), while a third found moderate to large effects (FM-UE Time 1, d = –0.75, FM-UE Time 2, d = 1.23). The effect on global function- Barthel Index (BI) was moderate (d = 0.47). These analyses indicate that NiCBS likely has a greater effect on impairment with relatively moderate effect on activity performance and participation.
The studies investigating the effects of cerebellar NIBS on posture and balance employed either iTBS or tDCS, and of the seven, three studies combined stimulation with practice/therapy. Four of these studies were RCTs, two were crossover studies, and one was a case-control study.
Of the seven studies, three studies applied iTBS over the contralesional cerebellum and four studies applied tDCS over the cerebellum to study the effects on balance and postural control. Analyses of effect size for posture and balance outcomes indicated a moderate to large effect size (d = 0.92–4.78) on measures of balance using the Berg Balance Score (BBS), BESTest (Koch et al., 2019; Liao et al., 2021; Qurat-ul-ain et al., 2022). Objective measures of CoP movements showed moderate (d = 0.73–0.75 (Ranjan et al., 2021)) to large (d = 1.59 (Xia et al., 2022)) size of NiCBS effects.
Of the seven studies, two applied iTBS over the contralesional cerebellum, one applied anodal stimulation to target dentate nuclei and the lower-limb representations (leg lobules VII-IX); two applied anodes 1–3 cm lateral to the inion, and the final two placed a cathode over the cerebellar hemisphere and an anode over the buccinator muscle of the same side. Analyses of effect size for walking outcomes indicated small effect size (d = 0.43–0.46) on measures of the 6 Minute Walk Test (6MWT), Motricity Index (MI) at a two-week follow up, Step width of the unaffected side, and Cadence (Koch et al., 2019; Picelli et al., 2018). Moderate effect sizes (d = 0.51–0.72) were found on measures such as MI immediately following treatment and at a 4-week follow up, and the Timed up and go test (TUG) scores (Picelli et al., 2018; Qurat-ul-ain et al., 2022).
The scoping review presented thus far provides preliminary evidence that NiCBS may be an effective adjunct to support movement recovery in individuals with stroke. While encouraging, multiple limitations of this scoping review call for critical considerations prior to clinical application. First, most included proof-of-concept studies have small sample sizes; thus, raising the risk of effect size inflation. Given the limited focality of noninvasive stimulation methods and the complexity of functional cerebellar organization, it is unclear which specific structures are stimulated and consequently, which mechanisms are implementing the observed effects. For example, the physiological effects of NiCBS on motor cortical excitability and plasticity after stroke are currently unclear. It is therefore crucial that larger sample studies in individuals with known brain lesions are conducted with concomitant neurophysiological measurements of changes in cerebellar-brain connectivity and cortical plasticity. Women were largely underrepresented in these studies, as is the case in many stroke NIBS trials. Finally, inadequate description of stimulation parameters and accompanying behavioral interventions preclude clear recommendations for integrating NiCBS with behavioral/clinical interventions.
The effectiveness of NiCBS can be further enhanced through an informed approach that combines it with well-defined targeted behavioral interventions. Distinct behavioral interventions rely on distinct mechanisms to implement their beneficial effect on treatment targets. A conceptual framework that considers cerebellar connections with cerebral cortex and brainstem networks may provide a theoretical basis for combining NiCBS and distinct behavioral interventions to target specified mechanisms for movement recovery.
Theory-driven mechanistic framework for noninvasive Neuromodulation of the cerebellum
A theory-driven framework that considers cerebellar specialization and connectivity to influence specific treatment targets underlying movement recovery is needed to inform the choice of NiCBS parameters, concurrently administered behavioral interventions and outcomes in neurorehabilitation research. Figure 4 outlines cerebellar functional regions and emergent cerebello-cortical and cerebellar-brainstem pathways that support mechanisms to improve specific treatment targets during post-stroke recovery. Below, we elaborate on each cerebellar region, and discuss the effects of cerebellar neuromodulation on physiological and/behavioral processes.
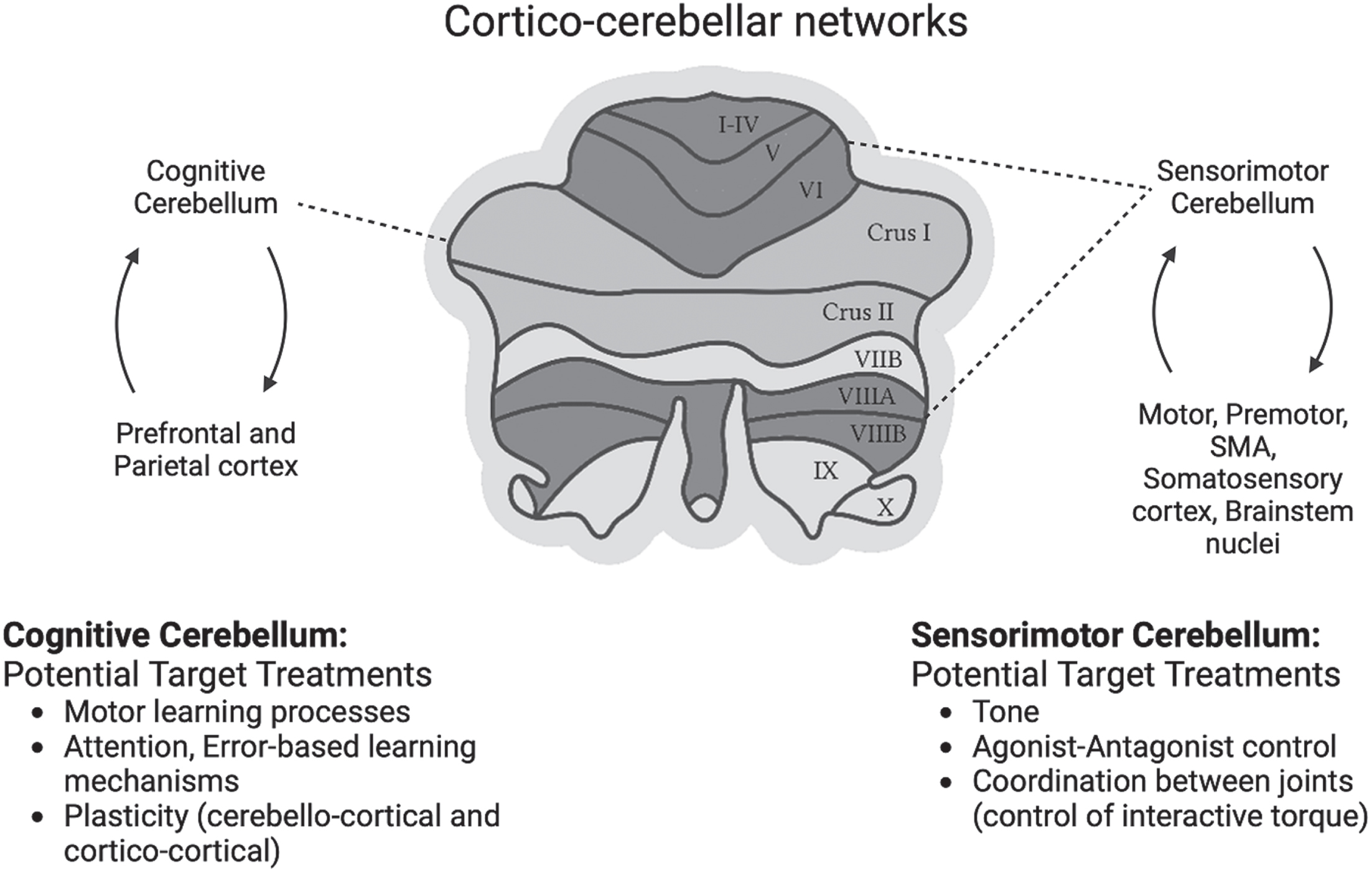
Cerebellum has motor (green) and cognitive (pink) regions. Reciprocal cortico-cerebellar networks to and from motor and cognitive parts of the cerebellum putatively underlying sensorimotor and cognitive processes. Each of these networks form mechanisms for distinct treatment targets. Sensorimotor cerebellum connected to the motor-premotor cortex and brainstem nuclei has the potential to influence tone, agonist-antagonist control and coordination of joints crucial for individuation. Cognitive cerebellum, reciprocally connected to the prefrontal and parietal cortices, may influence cognitive processes such as attention, adaptation and learning.
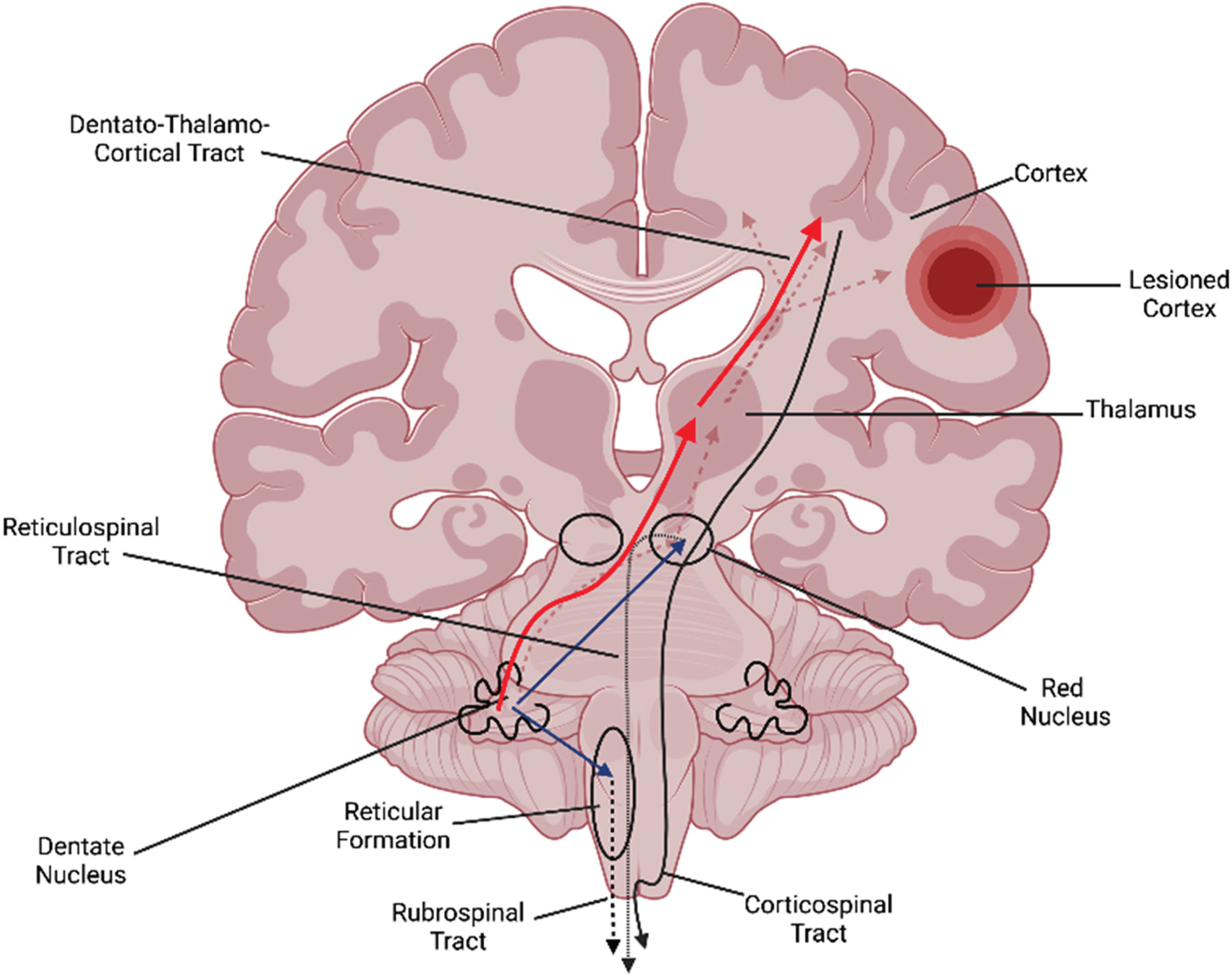
Cerebellar stimulation influences (a) dentato-thalamo-cortical pathway (red) to influence motor cortex and (b) cerebellar-brainstem pathways (blue arrows) to the red nucleus and reticular formation and modulate the rubrospinal and reticulospinal pathways.
A key motor deficit after stroke is the inability to coordinate multiple joints for functional actions such as reaching. Such impaired intersegmental coordination during multi-joint movement arises from deficits in feedforward control that requires planning for the effects of interactive torques (passive torques on a joint due to a movement at an adjacent joint) (Beer et al., 2000). Cerebellum plays a key role in feedforward specification of agonist-antagonist timing as well as coordination between different body segments prior to execution. NiCBS of the sensorimotor cerebellum, via its modulation of motor and premotor cortices, may help improve the ability to modulate muscle activity across multiple joints in anticipation of the mechanical interaction torques generated by one’s own movement and by external forces. NiCBS in neurotypical individuals alters cerebellar brain inhibition to the contralateral motor cortex, as well as influences contralateral corticospinal excitability and intracortical inhibition (Behrangrad et al., 2019; Koch et al., 2008). Whether these physiological effects can be harnessed to improve agonist-antagonist control as well as inter-joint coordination of the paretic arm is open for investigation.
Complex networks are instrumental in driving cognitive-motor learning mechanisms. Through the dentato-thalamo-cortical projections, NiCBS of the contralesional cerebellum alters resting state connectivity in frontal-parietal networks that support remediation of cognitive processes and boost their engagement in recovery. As proposed by Wessel and Hummel, cerebellar stimulation may also influence the cognitive network, and thus be a more effective node to engage a wider network supporting cognitive-motor learning and recovery (Wessel & Hummel, 2018). The influence of cerebellar stimulation on motor skill acquisition may be mediated through multiple interacting mechanisms (Fig. 6). First, NiCBS may directly enhance cerebellar-motor cortical plasticity crucial for specific processes involved in motor learning (Kishore et al., 2014; Koch et al., 2008; Rastogi et al., 2017). Further, NiCBS may prime the motor cortex rendering it more plastic for subsequent peripheral input or behavioral training (Kishore et al., 2021). Second, cerebellar stimulation in conjunction with motor practice may augment the engagement of frontal and parietal areas during motor learning (Casula et al., 2016; Zheng et al., 2023). Third, cerebellar stimulation may also modulate the connectivity between different cortical/subcortical networks (Halko et al., 2014; Rastogi et al., 2017) that implement practice-induced learning improvements. Recent work demonstrating network-wide changes in the perilesional motor, sensory, preSMA, dorsal and ventral premotor areas following deep brain stimulation and behavioral training offers support to these mechanisms (Baker et al., 2023).
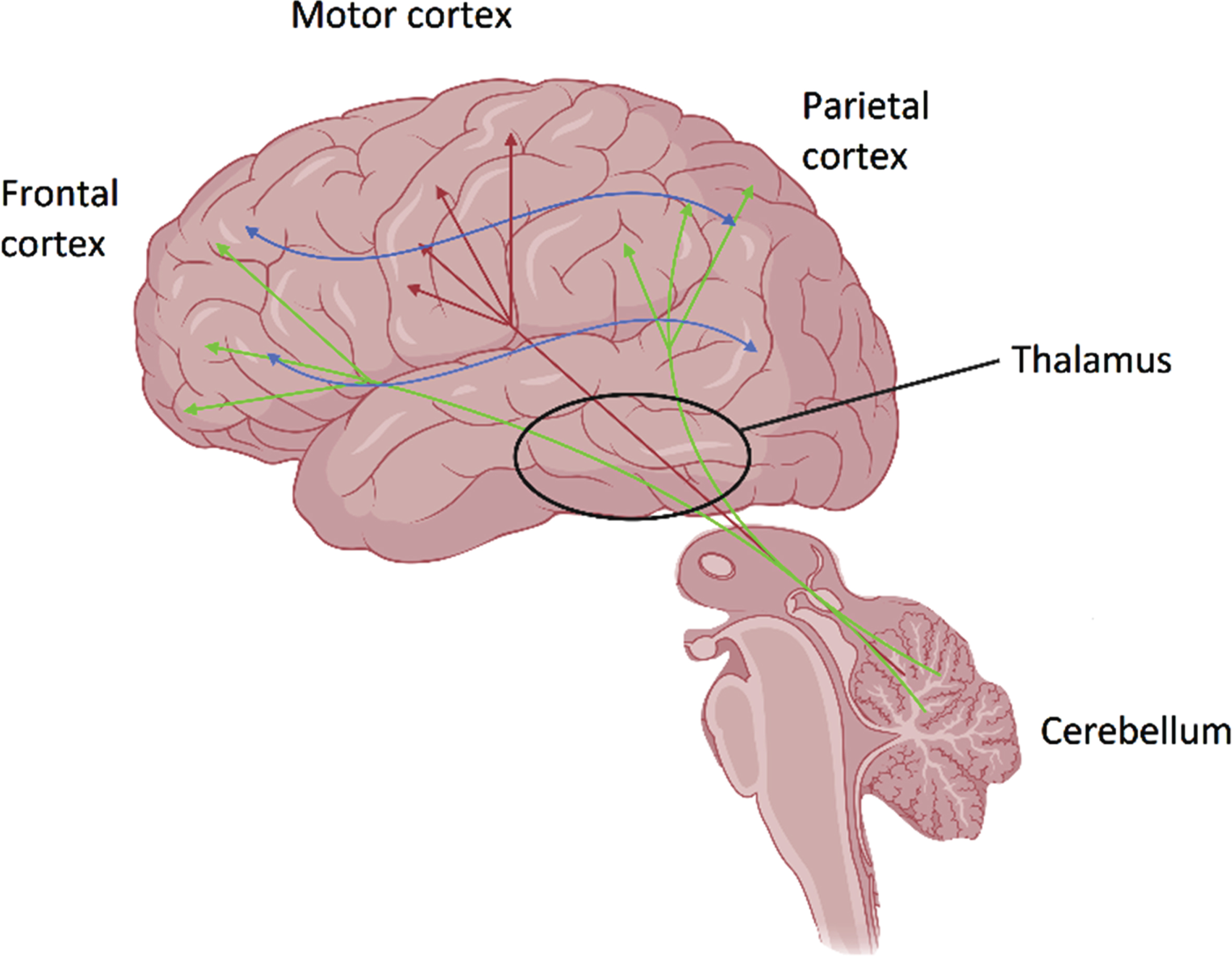
Cerebellar NIBS influences and likely facilitates plasticity within and across multiple contralateral cortical regions: (a) directly via the dentato-thalamo-cortical tract to motor cortex- engaging motor processes (red arrows); (b) directly via the dentato-thalamo-cortical tracts to frontoparietal areas (green arrows)- engaging cognitive processes; (c) indirectly prime connectivity and plasticity between different cortical areas (blue arrows).
Such distinct somatotopy, we suggest, has strong implications for NiCBS use in stroke recovery. Depending on the domain of the treatment target- motor vs. cognitive, it is ideal to tailor the stimulation to specific regions of the cerebellum. Invasive cerebellar stimulation has progressed to provide greater spatial resolution for stimulation; however, more efforts are needed to improve the spatial resolution of NiCBS. A combination of computational modeling and high-definition noninvasive paradigms such as tDCS or distinct coil models for TMS may be needed to improve the spatial resolution of NiCBS. Second, and a broader implication is that behavioral intervention aligned to the treatment target may yield better effects than nonspecific/undefined interventions. For intervention effects to be augmented, NiCBS should prime specific mechanisms/pathways driving the intervention effect for the intended treatment target. For example, if the goal is to improve interjoint coordination (treatment target), then both behavioral and NiCBS interventions must synergistically influence known mechanisms underlying interjoint coordination. While a detailed discussion of mechanisms-driven behavioral interventions for specific treatment targets is beyond the scope of this perspective, we provide a potential pairing of NiCBS and behavioral interventions to improve specific treatment targets in Table 2. These behavioral-NiCBS intervention synergies are conceptually driven, and further research is needed to demonstrate effectiveness of such synergies.
Mapping of functional region of the cerebellum, its connectivity with treatment target and behavioral interventions to improve specific outcomes (i.e., measure of the treatment target)
Thus far, the proposed theoretical framework provides multiple mechanisms through which NiCBS may target sensorimotor and cognitive processes critical to recovery of motor control and learning. Future research will require neurophysiological, behavioral, and clinical studies that fill in crucial gaps needed to refine NiCBS-behavioral intervention combination to target specific impairments, augment motor learning and improve movement recovery.
Current research in NiCBS is at a relatively incipient stage and would benefit from a staged research approach along the translational pipeline driven by a mechanistic framework such as one presented here. Below we outline future research areas in NiCBS to determine key mechanisms, identify and treat appropriate treatment targets, shed light on individual differences, and guide development of synergistic behavioral and neurophysiologic approaches to improve clinical outcomes in individuals with stroke.
Conclusion
There is emerging preliminary evidence for NiCBS as a promising adjunct in movement rehabilitation and recovery after stroke. A careful theory/mechanism-driven staged approach is needed to optimize the efficacy of NiCBS for improving movement outcomes in stroke rehabilitation. Establishing efficacy for combining NiCBS with behavioral interventions will pave way for future investigations to test effectiveness and infuse NiCBS in the clinic to improve patient outcomes.
Footnotes
Acknowledgments
During the time of writing this manuscript, SK’s time was funded by NIH R01-HD092481 and 1R01-HD104637 grants.
Conflict of interest
None of the authors have any conflict of interest to report.
Funding
The project was supported by MossRehab Peer Review Committee grant (FY22-2) to Shailesh Kantak.
{kind=link}
{kind=link}