
Abstract
Introduction
Cortical damage due to traumatic brain injury (TBI) or stroke often leads to persistent functional impairments if the motor cortex is part of the traumatized or infarcted brain region. The resulting motor impairments are disabling and form a major socio-economic burden (Parker, Wade, & Langton Hewer, 1986; Walker & Pickett, 2007). Thus far, the only clinically proven therapy for patients with motor deficits is physical rehabilitation therapy; still, many patients do not achieve full recovery. To enhance the efficacy of physical rehabilitation therapy, motor cortex stimulation (MCS) has been proposed as a potential therapeutic approach (Brown et al., 2006; Levy et al., 2008). Recently, the results of a multicenter study were reported, in which stroke patients suffering from hemiplegia received six weeks of MCS via implanted epidural electrodes concurrent with physical rehabilitation therapy (Levy et al., 2015). The authors reported a promising recovery course of the patients that was still present six months after cessation of the therapy (Levy et al., 2015). MCS has also been applied in rodent and non-human primate models of ischemic infarcts and resulted in improved limb function, again when being combined with physical rehabilitation therapy (Adkins et al., 2006; Baba et al., 2009; Plautz et al., 2003). However, limited data are available investigating the effect of MCS on its own without an additional intervention.
One of the mechanisms explaining the therapeutic effect of MCS on motor recovery is neuroplasticity. In a previous study, we found that MCS applied to naïve rats increased cell proliferation in the subventricular zone (SVZ) compared to non-stimulated controls (Jahanshahi et al., 2013a). Furthermore, a higher number of neural stem and progenitor cells (NSPCs) and mature neurons were detected in the motor cortex underneath the electrode (Jahanshahi et al., 2013a). This finding could be explained by a process called electrotaxis, where cell migration is induced by an electrical field (Jahanshahi et al., 2013b). In other studies using different forms of electrical stimulation, this increase of NSPCs at the side of stimulation was further corroborated, strengthening the hypothesis of electrotaxis in vivo (Morimoto et al., 2011; Rueger et al., 2012).
Animal models can be used to mimic clinical symptoms in a standardized way. With a controlled cortical impact (CCI), a cortical lesion can be created in rats that results in long-lasting functional deficits (Schönfeld et al., 2016). Similar to humans, rats possess a topographic organization of the motor cortex, where distinct cortical areas control the function of specific body parts (Nishibe et al., 2010; Starkey et al., 2012) and a CCI lesion in the forelimb area of the motor cortex can cause deficits in motor functions specific to the contralateral forelimb (Schönfeld et al., 2016).
In the present study, we tested whether MCS as a standalone treatment is able to achieve functional recovery in a rat model of severe CCI in the forelimb area of the motor cortex. To document functional recovery, we measured the effect of MCS on forelimb function and metabolic brain activity.
Materials & methods
Subjects
All animal experiments were conducted according to the directive 2010/63/EU on the protection of animals used for scientific purposes and had been approved by the local ethical committee for animal experiments at Maastricht University. Forty male Sprague-Dawley rats (Charles River, France), ten weeks old and weighing approximately 400 g at the time of surgery, were housed in pairs under a reversed 12 h light/dark cycle. Housing and testing facilities were kept at a constant temperature of 22° C and a humidity of 40–60%. Animals received standard laboratory chow (Sniff, Germany) and acidified water (pH 2.3–2.7) ad libitum, if not specified otherwise. Each behavioral assessment took place during the dark phase of the reversed night-day cycle (between 7 am and 7 pm), which is the active period of therats.
CCI induction and electrode implantation
Induction of CCI was performed as previously described in detail (Schönfeld et al., 2016). Shortly, a craniotomy was made above the forelimb area of the motor cortex (coordinates AP 0–3.5 mm anterior to bregma, ML 0.5–4 mm lateral to bregma) contralateral to the dominant paw, as determined by baseline performance in the Montoya staircase test. All rats received a CCI using an electromagnetically driven impactor device (Leica Impact One, Leica Biosystems, USA) with an impactor tip of 3 mm diameter, an impact depth of 5 mm and a velocity of 3 m/s (Fig. 1c). Polyurethane-isolated flat electrodes (3.4×3 mm; Medtronic, USA) with six exposed monopolar platinum/iridium contact points, were positioned on top of the CCI lesion (Fig. 1a-b) and a reference wire was anchored to the contralateral skull (coordinates AP 1.75 mm anterior to bregma and ML 2.25 mm lateral to bregma). Both the electrode and the reference wire were fixed with dental cement (Paladur, Heraeus, Germany) exposing two electrode contact pins (Multi-Contact, Switzerland) on the back of the animal’s head to allow connection to an external electrical stimulator. Animals from the control group received non-functional dummy electrodes of the same size and material. After surgery, rats were left to recover for two weeks.

Location of the CCI and electrode placement. A schematic representation of the somatotopic organization of the rat motor cortex is shown, modified from Fonoff et al. (2016; a). The CCI targeted large parts of the forelimb area (FL; CCI area represented by a large circle), followed by electrode placement on top of the lesion (square). Small circles represent the monopolar electrode contacts that delivered the current. On the right, a scaled-down picture of the electrode lead is shown (real size 3.4×3 mm; b). CCI on the motor cortex damaged large parts of the cortex and the dorsal striatum (c; image shown at 1.7 mm anterior to bregma). CCI: controlled cortical impact; W: whiskers; N: neck muscles; FL: forelimb; HL: hindlimb; T: tail; E: eyes; M1: primary motor cortex; M2: secondary motor cortex; CG1: cingulate cortex 1; CG2: cingulate cortex 2; CC: corpus callosum; CPu: caudate putamen (striatum).
MCS was applied daily for 2 h during a period of 31 consecutive days. Stimulation parameters were chosen based on previous studies (Baba et al., 2009; Jahanshahi et al., 2013a; Teskey et al., 2003) and consisted of a frequency of 30 Hz, 1 ms pulse width and biphasic constant current set at 50% of the current that evoked a motor response. MCS was delivered by an external digital stimulator (DS8000, World Precision Instruments, Germany) while rats stayed individually in stimulation chambers (width 28 cm, depth 50 cm, height 47.8 cm) and were allowed to move around freely under conditions similar to their home cage. Control animals underwent the same procedure without any current being delivered.
Behavioral testing
Reaching and grasping abilities of both forelimbs were assessed with the Montoya staircase test. In short, rats have to retrieve sucrose pellets (Test Diet, USA) lying on each step of a staircase located on the left and on the right hand side of a platform inside a narrow translucent box (Montoya et al., 1991). All steps, except the highest one, were baited with three sucrose pellets. Rats were habituated to the staircase boxes and then trained daily until they retrieved a minimum of 55% of the pellets from at least one of the staircases (Windle et al., 2006). Testing sessions lasted 15 minutes and took place on two consecutive days, twice daily with a minimal inter-trial interval of three hours resulting in a total of four testing sessions per time point of behavioral assessment. During the training and testing period, rats were food deprived to 85–90% of their free-feeding weight to increase their motivation for pellet reaching. Data are presented as the total number of eaten pellets for the impaired and healthy forelimb separately, as well as using a difference score defined as the score of the healthy forelimb subtracted from the score of the impaired forelimb (pelletsimp – pelletsheal). Behavioral testing in the Montoya staircase test was performed at three time points: Before CCI, two weeks after CCI and after four weeks of (sham) MCS.
Paw use during vertical exploration was measured by the cylinder test as described previously (Schönfeld et al., 2016). Rats were transferred to Perspex cylinders on an illuminated platform and recorded from above during 10 minutes (GoPro Hero 4, GoPro, USA), while they explored the cylinder by rearing and leaning against the wall. Based on the video footage, the first twenty wall contacts were scored and used for analysis. Wall contacts were made using both paws individually (‘impaired’, ‘healthy’) or using both paws at the same time (‘both’). Data are presented as the percentage of the wall contacts with either the impaired or the healthy forelimb relative to the total twenty wall contacts (contactsimp/20 * 100 and contactsheal/20 * 100, respectively). In addition, the difference score of the percentages is shown to visualize asymmetry between both forelimbs (% contactsimp –% contactsheal). Behavioral testing using the cylinder test was performed at four time points: Before CCI, two weeks after CCI, two weeks after initiation of MCS or sham stimulation and after four weeks of MCS or sham stimulation.
Functional imaging
Distribution of 18F-fluorodesoxyglucose as an indirect indicator of glucose-related metabolic activity in the central nervous system was visualized in vivo using a μPET scanner (μPET Focus, Siemens, the Netherlands). Rats were anesthetized with Isoflurane and received 10–20 mBq 18F-fluorodesoxyglucose (FDG; GE Healthcare, the Netherlands) intravenously, immediately followed by scanning the entire brain for 30 minutes. Thereafter, a static image was reconstructed using OSEM2D. Each animal underwent μPET twice; on the first day rats from the MCS group were scanned with stimulation off, whereas on the second day stimulation was switched on 10 minutes before as well as throughout the entire duration of scanning in order to visualize potential acute effects of MCS by using autoradiography. Control animals were scanned twice under the same conditions, thus without any stimulation being delivered. The lesion volume was calculated by delineating the virtual CCI area, identified as the cortical area without any visual presence of FDG (Fig. 3a). The CCI lesion was delineated throughout its full length in static images of sequential brain slices using pmod image analysis software (PMOD 2.9, pmod Technologies, Switzerland).
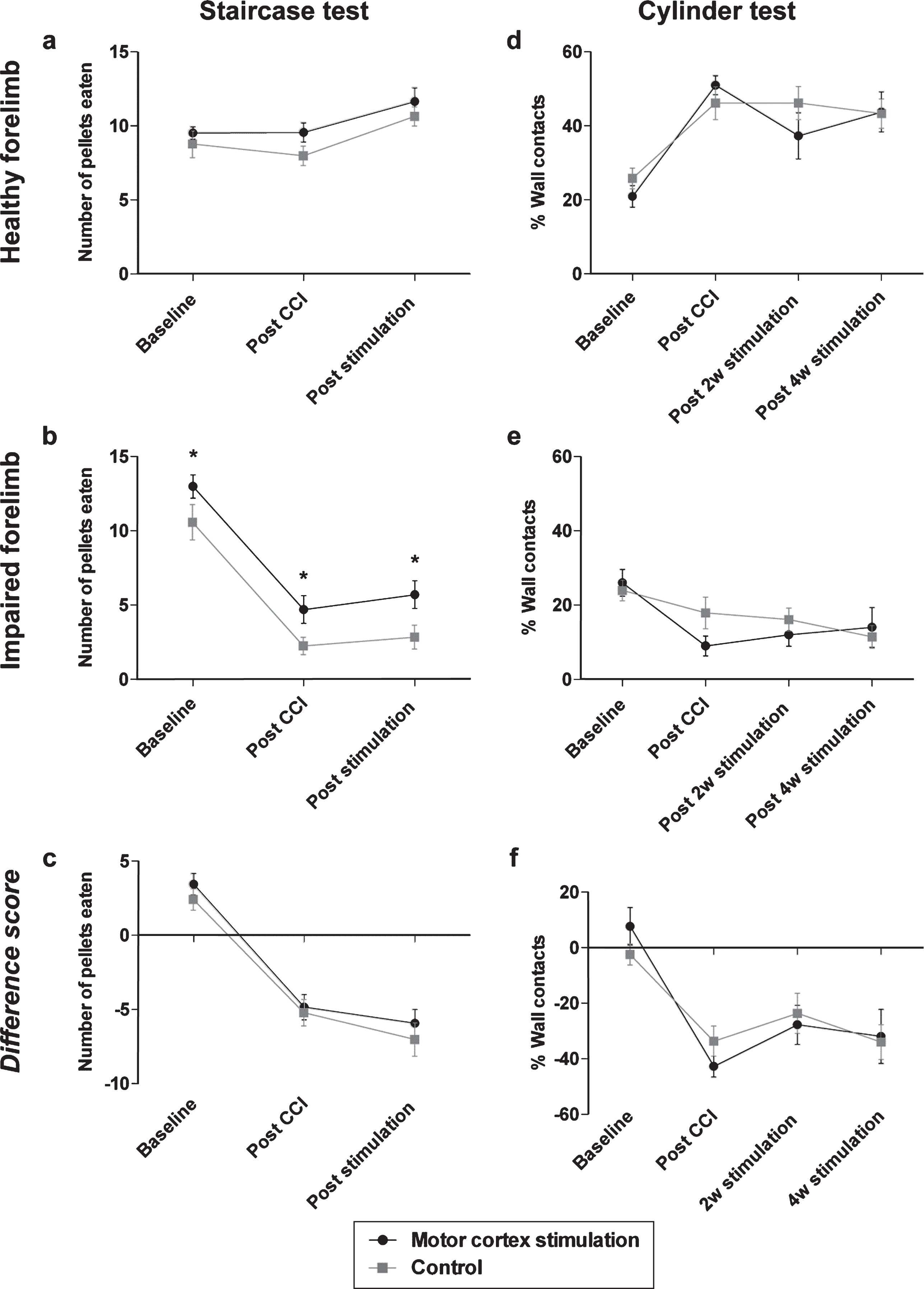
Chronic motor cortex stimulation after a unilateral CCI neither recovered reaching and grasping skills nor paw use during vertical exploration. CCI to the forelimb area of the motor cortex did not affect pellet retrieval with the healthy paw (a) whereas it resulted in significantly less pellet retrieval with the impaired paw (b), as measured in the Montoya staircase test. This behavioral impairment was unaltered by MCS. Concerning the impaired paw, an overall significant difference between groups at all time points was detected, which was unrelated to the application of MCS. The difference score also reflected the tendency of less pellets eaten with the impaired paw (c). Use of the cylinder test showed that after a unilateral CCI rats increased the use of their ipsilateral paw to lean against the cylinder walls (d), while neglecting the paw contralateral to the CCI lesion (e). The difference score also show a decreased use of the impaired forelimb to lean against the cylinder wall (f). This effect of CCI on motor impairment was not restored by MCS. CCI = controlled cortical impact; MCS = motor cortex stimulation; *p < 0.05.

Neither lesion volume nor glucose distribution was affected by chronic motor cortex stimulation. A representative μPET image is shown in a horizontal plane, used to delineate the lesion area (white line, a). Chronic MCS of the lesion area did not significantly change the size of the CCI lesion (b). Representative autoradiography images are shown of a control rat (c) and a rat that received motor cortex stimulation (d). The radioactive signal intensity (arbitrary units, a.u.) for the healthy (e) and lesioned hemisphere (f) separately did not differ between control rats and rats that were stimulated. Also no difference between groups was present in the ratio of the signal intensity between the lesioned hemisphere and the healthy hemisphere. CCI: controlled cortical impact; M: medial; L: lateral; D: dorsal; V: ventral.
Autoradiography was performed after the second μPET scan to visualize FDG distribution at a high spatial resolution. After transcardial perfusion with 4% paraformaldehyde, brains were frozen and cut into 50 μm thick sections. Autoradiography phosphor plates (GE Healthcare, the Netherlands) were exposed to the frozen brain sections during approximately 2 hours and read for each animal with a Typhoon FLA7000IP scanner (GE Healthcare, the Netherlands). For each animal, the entire lesioned and healthy hemispheres of three sections at a comparable bregma level were delineated separately and the intensity of the FDG signal for the individual hemispheres was measured using ImageQuant TL software (GE Healthcare, the Netherlands). Signal intensity (arbitrary units, a.u.) depended on the amount of radioactive counts present in the delineated area and was corrected for the injected amount of MBq, animal weight, the time from injection until exposure to the autoradiography plate and the duration of exposure on the autoradiography plate. To correct for inter-individual fluctuations, intensity values were expressed by dividing the intensity values measured within the lesioned hemisphere by the intensity values measured within the healthy hemisphere for each individual animal. In addition, intensity values for each hemisphere are shown separately.
Data are presented as mean±standard error of the mean and were analyzed with repeated-measures ANOVA (SPSS 20, IBM, US) with time (baseline, post CCI and post stimulation) as within-subjects factor and group (MCS and control) as between-subjects factor. Imaging data were analyzed with one-way ANOVA and p-values below 0.05 were considered significant. Values below or above 1.5 interquartile ranges were identified as outliers by SPSS and excluded. In addition, animals that lost their electrodes during the course of the experiment and therefore could not undergo stimulation during 31 days were excluded from the analysis.
Figures were created using GraphPad Prism 5 (GraphPad Software, US), Adobe Illustrator CS6 (Adobe, US) and Microsoft PowerPoint 2011 (Microsoft, US).
Results
Chronic motor cortex stimulation failed to recover grasping skills and paw asymmetry during vertical exploration behavior
The Montoya staircase test was used to assess the recovery of reaching and grasping skills that were impaired by severe CCI. Repeated measures ANOVA revealed that the number of eaten pellets with the healthy paw increased after CCI compared to the number of eaten pellets before CCI for both groups [F(2, 44) = 14.27, p < 0.001; Fig. 2a] without any significant difference between rats that received MCS and rats that received sham stimulation. The number of eaten pellets with the impaired paw decreased after CCI [F(2, 46) = 85.18, p < 0.001; Fig. 2b] and a significant difference was detected between the groups at all time points [F(1, 23) = 6,18, p < 0.05]. This difference between the groups, however, was constant at all timepoints (2.41, 2.45 and 2.87 pellets, representing the mean number of pellets eaten before CCI, after CCI and after four weeks of MCS or sham stimulation respectively), which indicates a lack of functional improvement caused by MCS. The difference score was calculated by subtracting the number of pellets eaten with the healthy paw from the number of pellets eaten with the impaired paw. Using the difference score, a decline over time was shown implying worse pellet retrieval with the impaired paw in both groups [F(2, 46) = 71.44, p < 0.001; Fig. 2c]. The analysis of the difference score also revealed that MCS treatment did not affect the number of pellets eaten after CCI. These results indicate that MCS did not have any effect on grasping and reaching behavior after CCI.
The cylinder test was performed to measure vertical exploration behavior with the individual forelimbs. After CCI, all animals showed an increased reliance on the healthy paw to lean against the cylinder wall [F(3, 69) = 19.30, p < 0.001; Fig. 2d] at the expense of using their impaired paw [F(3, 66) = 6.77, p < 0.001; Fig. 2e]. However, MCS treatment did not restore usage of the impaired paw. Analysis of the difference score showed a stronger asymmetry in paw use after CCI compared to paw use before CCI [F(1.8, 66) = 19.73, p < 0.001; Fig. 2f]. Yet, treatment with MCS could not resolve this asymmetry in wall contacts between both forelimbs. Taken together, these results indicate that treatment with MCS could not restore forelimb use for vertical exploration behavior.
Lesion volume and glucose metabolism in the lesioned hemisphere did not change despite motor cortex stimulation
Delineation of the lesion area in the reconstructed μPET images was performed to estimate the amount of histological damage that was present after chronic application of MCS (Fig. 3a). The lesion volume of animals that received chronic MCS was not significantly different from the lesion volume of non-stimulated controls [F(1,8) = 2.98, p > 0.05; Fig. 3b], which indicated a lack in overt tissue recovery.
Imaging the distribution of FDG in brain slices was performed by means of autoradiography to measure functional recovery that may have occurred at the tissue level (Fig. 3c-d). The ratio of intensity values between the healthy and the lesioned hemisphere did not differ between rats that received MCS and control rats that received sham stimulation [F(1,7) = 0.12, p > 0.05; Fig. 3g]. When comparing the intensity values of the delineated lesioned and healthy hemisphere separately, no difference was detected between the animal groups, either [lesioned hemisphere: F(1,7) = 0.64, p > 0.05; healthy hemisphere: F(1,7) = 0.61, p > 0.05; Fig. 3e-f], showing again that MCS treatment did not influence FDG distribution.
Discussion
Motor impairments are among the most debilitating consequences of stroke or TBI and have a strong impact on a patient’s day-to-day activities. Electrical stimulation of the motor cortex has been shown to cause functional improvements in animal models of ischemic stroke and the ability of MCS to improve motor recovery in humans is currently under investigation (Levy et al., 2015; Shin et al., 2014). In rodent models of stroke, functional recovery was measurable as improved limb placement in response to a sensory cue, grasping or balance (Cheng et al., 2012; Moon et al., 2009). An increased formation of new blood vessels and dendritic sprouting has been found in addition to more NeuN-positive cells in the ischemic cortex (Cheng et al., 2012; Kang et al.,2013).
Although stroke and TBI have a different cause, both result in strikingly similar effects at the cellular level, such as excitotoxicity, oxidative stress and inflammatory responses (Lopez et al., 2015); therefore treatments effective in animal models of stroke might also be applicable to animal models of TBI. However, compared to stroke, endogenous plasticity processes are more limited after CCI lesions and more diverse behavioral rehabilitation training is necessary to induce morphological and functional rehabilitation (Combs et al., 2016; Jones et al., 2012). In the specific case of MCS, stimulation parameters that induced robust recovery in stroke models were less effective in a TBI model with comparable motor impairments (Jefferson et al., 2015).
In the current study the potential of chronic MCS to achieve functional recovery after a cortical lesion was assessed. To induce the cortical lesion, rats received a CCI in the forelimb area of the motor cortex contralateral to the preferred limb. MCS was applied to freely moving rats during a period of 31 consecutive days and different aspects of forelimb function were assessed before and after the stimulation period. We measured fine motor skills with the Montoya staircase test and the cylinder test and both tests are sensitive ways to detect asymmetrical paw use after a cortical lesion (MacLellan et al., 2013; Schönfeld et al., 2016; Windle et al., 2006). Furthermore, comparable behavioral tests were used in studies where a regenerative effect of MCS was detected (Morimoto et al., 2011; Zhou et al., 2010). In addition, we visualized the distribution of a radioactive glucose analog, FDG, to detect potential changes in brain metabolism after chronic MCS.
In line with previous research, we found that severe CCI created long-lasting motor impairments specific to the contralateral forelimb (Schönfeld et al., 2016), which could be detected for up to eight weeks after the insult. Motor impairments observed in patients with a cortical lesion usually have a chronic course; therefore modeling long-lasting motor impairments in animals is essential and can be achieved by severeCCI.
In the current study, we wanted to test the therapeutic potential of MCS as a standalone treatment administered in a home cage-environment. This procedure was different compared to previous research, where MCS was always administered together with physical rehabilitation training. In the current study, MCS as an independent treatment did not cause any improvement of motor impairments. After CCI, reaching and grasping skills with the impaired limb were equally affected in rats that received MCS compared to sham-stimulated control rats. Also, after CCI rats predominantly used their healthy forelimb during vertical exploration whereas usage of the other, impaired, forelimb was not restored after chronic MCS.
In line with these findings, no changes in either lesion size or FDG distribution were detected after the application of MCS. Changes in FDG distribution can be used as an indicator of neural activity (Gobel et al., 2013); therefore increased FDG distribution after MCS would have been an indirect measure of tissue restoration, whereas the absence of any MCS-induced change suggests a lack of treatment effect on energy metabolism.
A few studies on TBI in rats have reported a therapeutic effect of MCS (Jefferson et al., 2015; Yoon et al., 2015; Yoon et al., 2012). In those studies, MCS was co-administered with behavioral rehabilitation training during a period ranging from two (Yoon et al., 2015; Yoon et al., 2012) to nine weeks (Jefferson et al., 2015). In all studies an improvement of forelimb function at the end of the stimulation period was reported (Jefferson et al., 2015; Yoon et al., 2015; Yoon et al., 2012), together with an increase in size of the cortical area responsible for wrist movement, as assessed by intracortical microstimulation mapping (Jefferson et al., 2015). However, the TBI lesions created in these studies were considerably smaller compared to the lesion we applied, which may have increased the likelihood of regeneration. Also, in these studies the implanted electrodes did not only cover the damaged cortex, but also stimulated spared cortical regions, which could facilitate re-mapping of lost functions onto the surrounding cortex. Lastly, MCS was always administered together with behavioral training, which is representative of the clinical situation, but does not provide any information about the therapeutic effect of MCS on its own. Three factors might explain why MCS did not induce functional regeneration in our study. First, the present CCI lesion was very severe and damaged the corpus callosum and parts of the striatum in addition to the entire forelimb area of the motor cortex. We chose for inducing such a severe CCI since previously we have been able to measure long-lasting behavioral impairments after this specific form of CCI (Schönfeld et al., 2016). In studies using milder CCI lesions in the motor cortex, spontaneous recovery of motor functions was measured, starting already at 2 weeks after the lesion (Goffus et al., 2010; Nishibe et al., 2010; Shijo et al., 2015). Taken together, behavioral improvement might be more likely after a milder lesion since the area to regenerate is smaller and surrounding cortical regions are spared which might allow functional re-mapping of the lost area.
Second, in numerous studies, in which MCS caused functional improvements, it was not chosen to stimulate the damaged brain area directly; instead stimulation electrodes were implanted on top of the lesion penumbra (Adkins et al., 2006; Boychuk et al., 2011; Jefferson et al., 2015; Moon et al., 2009; O’Bryant et al., 2014; Plautz et al., 2003; Zhou et al., 2010). Electrical stimulation of the spared surrounding cortex may induce plasticity processes in contrast to the stimulation of a damaged brain region that during the course of several weeks develops into a large morphological cavity (Schönfeld et al., 2016). In a number of studies the therapeutic effect of MCS was not explained by tissue restoration at the lesion side, but by remapping of the lost functions onto the spared cortex around the lesion (Boychuk et al., 2011; Jefferson et al., 2015; Teskey et al., 2003). In the current study, the lesion area was stimulated directly to clarify whether the influence of MCS on NSPCs found in an earlier experiment (Jahanshahi et al., 2013a) could rescue the damaged cortical tissue. In a previous study we showed an increased amount of NSPCs at the stimulated cortex of naïve rats, which presumably had migrated from the SVZ (Jahanshahi et al., 2013a). Under in vitro conditions, electrical fields can induce migration of cells towards the current source, a process known as electrotaxis (Babona-Pilipos et al., 2011; Babona-Pilipos, Pritchard-Oh, Popovic, & Morshead, 2015; Feng et al., 2012; Liu et al., 2015; McCaig et al., 2005), and in vitro electrotaxis is a widely proven phenomenon occurring in different cells types, including NSPCs (Babona-Pilipos et al., 2015; Feng et al., 2012; Liu et al., 2015). Also, stimulation of the striatal penumbra after ischemic stroke in rats has been shown to increase the number of proliferating cells in the vicinity of the electrode and this finding co-occurred with a decreased lesion size and behavioral improvement (Morimoto et al., 2011). However, contrary findings have been reported as well. In a different study, a non-invasive form of cortical stimulation was applied in naïve rats and did not lead to directional migration of labeled neural stem cells in response to an electrical field (Keuters et al., 2015). The authors concluded that accumulation of neural stem cells at the stimulated cortical area is rather due to local cell proliferation and not to cell migration from neurogenic regions (Keuters et al., 2015). These contrary findings indicate, that in vivo electrotaxis first needs to be reliably demonstrated, before its role in stimulation-induced motor recovery can be investigated.
Third, in previous studies MCS has been delivered while the animals underwent rehabilitative therapy in the form of repetitive reaching with the impaired forelimb (Boychuk et al., 2011; Jefferson et al., 2015; Teskey et al., 2003). Probably an additional behavioral stimulus, that is stronger and more specific to the impaired forelimb than mere locomotion in a home cage, is necessary to cause improvement through MCS. Pairing of MCS with rehabilitation therapy may result in a synergistic effect and there are only a few reports showing a therapeutic effect of MCS without any additional intervention (Adkins-Muir & Jones, 2003; Adkins et al., 2006; Zhou et al.,2010).
In conclusion, the use of MCS as a standalone treatment did not improve motor impairments in a rat model of severe cortical damage using our specific treatment modalities.
Disclosure statement
No competing financial interests exist.
Footnotes
Acknowledgments
This work was supported in part by a grant from Hasselt University (BOF11603BOFMO) to SH and a grant from the Hersenstichting Nederland and Medtronic Europe (A1196117) to YT and AJ.