
Abstract
Objective:
Mesenchymal stem/stromal cells (MSC) promote recovery after spinal cord injury (SCI) using adult bone marrow MSC (BM-MSC). Newborn tissues are a convenient source of MSC that does not involve an invasive procedure for cell collection. In this study the authors tested the effects of rat amnion MSC clone (rAM-MSC) in SCI.
Methods:
We tested intra-parenchymal injection of a GFP+ rat rAM-MSC clone derived from E18.5 rats in rat SCI and measured behavioral recovery (BBB scores), histology and X-ray opacity. Expression of aggrecan was measured in culture after treatment with TGFß.
Results:
Injection of rAM-MSC after SCI did not improve BBB scores compared to control vehicle injections; rather they reduced scores significantly over 6 weeks. Spinal cords injected with rAM-MSC were hard in regions surrounding the SCI site, which was confirmed by X-ray opacity. Whole mount imaging of these cords showed minimal tissue loss in the SCI site that occurred in SCI controls, and persistence of GFP+ rAM-MSC. Mason’s Trichrome staining of tissue sections showed more intense staining for extracellular matrix (ECM) surrounding and extending beyond the SCI site with injections of rAM-MSC but not in controls. In response to TGF-ß treatment in culture, chondrogenic aggrecan was expressed at higher levels in rAM-MSC than in rBM-MSC, suggesting that the upregulation of TGF-ß in SCI sites may promote chondrogenic differentiation.
Conclusion:
Acute injection after SCI of a clonally expanded rAM-MSC resulted in aberrant differentiation towards a chondrocytic phenotype that disrupts the spinal cord and inhibits behavioral recovery after SCI. It will be critical to ensure that injection of extensively expanded neonatal cells do not differentiate aberrantly in traumatic CNS tissue and disrupt recovery.
Introduction
Mesenchymal stem/stromal cells (MSC) have attracted wide interest for potential medical applications because they have beneficial effects in many animal models. They are under investigation for several clinical indications (Figueroa, Carrion, Villanueva, & Khoury, 2012; Singer & Caplan, 2011), and have been approved for graft vs. host disease (GvHD) in Japan (http://www.mesoblast.com/partnerships/overview). MSC are much better tolerated than other cells after non-autologous transplantations, but risks of their use may require additional studies (Bernardo & Fibbe, 2012). MSC have been isolated from several adult tissues, most notably, bone marrow and fat. They also have been obtained from newborn tissues including placenta, amnion, decidua basalis, amniotic fluid, cord blood and Wharton’s jelly (Hass, Kasper, Bohm, & Jacobs, 2011). MSC derived from newborns may be advantageous over adult MSC as they are obtained without invasive procedures and may have greater abilities to proliferate while maintaining multipotentcy (Chen, Yen, Liu, Sytwu, & Yen, 2011).
Allogenic grafts of MSC are rejected minimally because they lack MHC class II antigens. Syngeneic grafts of MSC are rejected to an even lessor extent and autologous MSC should not be rejected at all. However, a high percentage of MSC injected intravenously disappear rapidly as they get trapped in the lungs, liver, spleen and other tissues (Sohni & Verfaillie, 2013), and the limited survival of MSC in various locations is difficult to measure (Figueroa et al., 2012). A major factor responsible for MSC disappearance may be reduced survival in locations that do not provide a supportive niche such as exists in regions where they are normally found (Kfoury & Scadden, 2015). In any case, human MSC have been found to improve outcomes in humans following transplantation, e.g. GvHD (Kurtzberg et al., 2014) and they survive transiently after transplant in sites of disease and injury such as in SCI rats (Osaka et al., 2010).
A major criterion for MSC identity is their ability to differentiate under defined conditions into chondrocytes, osteocytes and fat in vitro (Pittenger et al., 1999). MSC derived from newborn tissues may be more plastic than MSC from adults (Hass et al., 2011) but relatively few studies have injected them into the central nervous system (CNS). Here we studied mesenchymal stem/stromal cells (AM-MSC) derived from rat amnion. We expected that the MSC would promote recovery after SCI but surprisingly syngeneic rAM-MSC exhibited chondrogenic differentiation and had detrimental effects on locomotor recovery.
Materials and Methods
Rat amnion derived stromal cells (rAM-MSC) and bone marrow MSC (rBM-MSC)
rAM-MSCs were isolated from explant cultures of Sprague Dawley rat amniotic membranes (Marcus, Coyne, Rauch, Woodbury, & Black, 2008) and transfected to express GFP (Marcus, Coyne, Black, & Woodbury, 2008). The cells were cultured in MEM supplemented with 2 mM glutamax, 100μg/ml streptomycin, 100 U/ml penicillin, and 15% FCS (complete medium). For induction, cells were treated with 2 mM valproic acid, 15 mM betadine, 2.5 mM taurine, 175μM butylatedhydroxyanisole, 27 nM selenium, 20 nM progesterone, 10μM forskolin, 10 nM K252a, 5 units/ml heparin, 5μg/ml insulin, 1 mM sodium pyruvate, 50μM alpha thioglycerol, 20 nM bathocuproindisulfonic acid in DMEM pH 7.0 for 1 week. The media was supplemented with 10 ng/ml basic fibroblast growth factor at 24 h post-induction. These cells have been characterized as mesenchymal stem cell by FACS analyses (Marcus, Coyne, Rauch, et al., 2008). Rat BM-MSC were isolated from Fisher GFP+ rats (Marano, Sun, Zama, Young, & Uzumcu, 2008) by flushing the contents from femurs and tibia as described (Tokalov et al., 2007). For in vitro experiments, rAM-MSCs and rBM-MSC were cultured for 3 days with or without 10 ng/ml of TGF-β in complete medium, and immunostained with monoclonal antibodies against aggrecan (Santa-Cruz). For transplantation experiments, rAM-MSC were harvested by trypsinization before they reached confluence.
Animal surgery, spinal cord contusion, transplantation and behavior
Female Sprague Dawley rats (77±1 days) were housed under standard conditions (Constantini & Young, 1994; Hasegawa et al., 2005). Rats were anesthetized using intra-peritoneal injection of sodium pentobarbitone (45 mg/kg of body weight), and a dorsal midline incision of 3 cm was made to expose T8 to T11 spinous process. Laminectomy was carried at T9-T10 level and the spinal cord was exposed keeping the dura intact. Contusions were performed using MASCIS impactor by dropping a 10 g weight from a height of 25 mm. Injections were performed within 15 min after contusion using microliter syringes fitted with glass capillaries having a pore diameter of ∼30–50μm into the injury epicenter and 2 mm rostral and caudal to it as described (Hasegawa et al., 2005). Three sites were injected with 2×105 cells in 2μl in the lesion epicenter and 1×105 cells each at sites 2 mm rostral and caudal from the epicenter. Injections were performed slowly over a period of 3 minutes followed by an additional 3 minutes before withdrawing the capillaries to prevent backflow. Transplantation groups included GFP+ rAM-MSC, without or with induction, using DMEM as controls. The injury site was closed in layers and rats were returned to their cages after recovering from anesthesia. Rats received cefazolin (25 mg/kg/day) for 1 week after SCI. Bladders were expressed manually twice a day until control returned. When used, cyclosporine A (CsA) was injected subcutaneously for the entire period of the study at 10 mg/kg/day (Hasegawa et al., 2005). Locomotor recovery was assessed weekly using the 21-point BBB score (Basso, Beattie, & Bresnahan, 1996; Basso, Beattie, Bresnahan, et al., 1996) by two separate BBB scoring teams that were unaware of experimental treatments beginning on the second day and then weekly for up to 6 weeks after SCI. With the 25 mm weight drop used herein, hindlimb exhibited flaccid paralysis and BBB scores within the first 3 days are typically <1 (Basso, Beattie, & Bresnahan, 1996). BBB scores of 1–7 represents hindlimb movements with no weight support; a score of 8 signifies sweeping or plantar placement without weight support, while a score of 9 indicates plantar placement weight support in stance only with no plantar stepping. A score of 10 indicates occasional weight supported plantar steps without forelimb coordination while a score of 11 requires consistent weight-supported plantar stepping with no forelimb-hindlimb coordination. Scores of 12 and 13 represent frequent to consistent weight supported plantar steps with occasional forelimb-hindlimb coordination. Values represent mean±Standard Error of Mean. The Animal Care and Use Committee of Rutgers, The State University of New Jersey approved all rat experimental protocols.
Euthanasia, tissue processing sectioning and immunohistochemistry
Rats were deeply anesthetized and perfused intracardially with saline followed by 4% paraformaldehyde. Spinal cords were extracted and some were visualized for GFP in whole mounts using a Zeiss Stemi SV 11 fluorescence microscope to determine the distribution of the transplanted GFP+ cells. After post-fixation overnight and cryoprotection using 25% sucrose in PBS, 1-cm long horizontal spinal cord 20μm cryosections centered around the lesion epicenter were mounted on super frost slides (Hasegawa et al., 2005). Sections were stained using Masson’s trichrome to detect connective tissue including collagen as a green/blue color using a protocol adopted from (Culling, 1974). Monoclonal antibody CS56 (Sigma-Aldrich, St. Louis, MO), which recognizes chondroitin sulfate on proteoglycans, was used to stain ECM as described (Iseda et al., 2008). Stained sections were imaged using a Zeiss Automated Cell Scan System for Axiovert 200 M, or a Zeiss 510 LSM confocal microscope to obtain a single composite image of a large area centered on the SCI epicenter. Anti-Aggrecan (Santa Cruz, 25674) (1 : 100) was used to stain cells in culture (Hasegawa et al., 2005). Some spinal cords were X-rayed for opacity prior to sectioning.
Results
In preliminary experiments, injection of human AM-MSC (hAM-MSC) acutely in SCI did not improve behavioral recovery in Sprague Dawley rats over a 6-week period (data not shown). This may be due to loss of the human cells in the rat, which was difficult to test. Therefore, we decided to test GFP-expressing rat AM-MSC, which should be more compatible in Sprague Dawley rat SCI than hAM-MSC and could be detected by fluorescence (Marcus, Coyne, Black, et al., 2008). Considering that Sprague Dawley rats are outbred, we treated them with CsA to minimize rejection. The rAM-MSC have been characterized previously, and after neural induction they secreted higher levels of neurotrophic factors (Marcus, Coyne, Black, et al., 2008), which may make them more effective in promoting recovery after SCI. After a severe 25-mm weight drop with the MASCIS Impactor (Basso, Beattie, & Bresnahan, 1996), rAM-MSC with or without induction in culture were injected into the injury epicenter and 2 mm rostral and caudal to it. BBB scores at 2 days after this injury were 0, reflecting the flaccid paralysis due to spinal shock (Basso, Beattie, & Bresnahan, 1996). Surprisingly, the BBB scores of rats injected with rAM-MSC were significantly lower than vehicle injected rats at 2–6 weeks, which was ∼8 (Fig. 1A). The reduction in BBB scores was more pronounced with the induced rAM-MSC compared to the non-induced rAM-MSC.
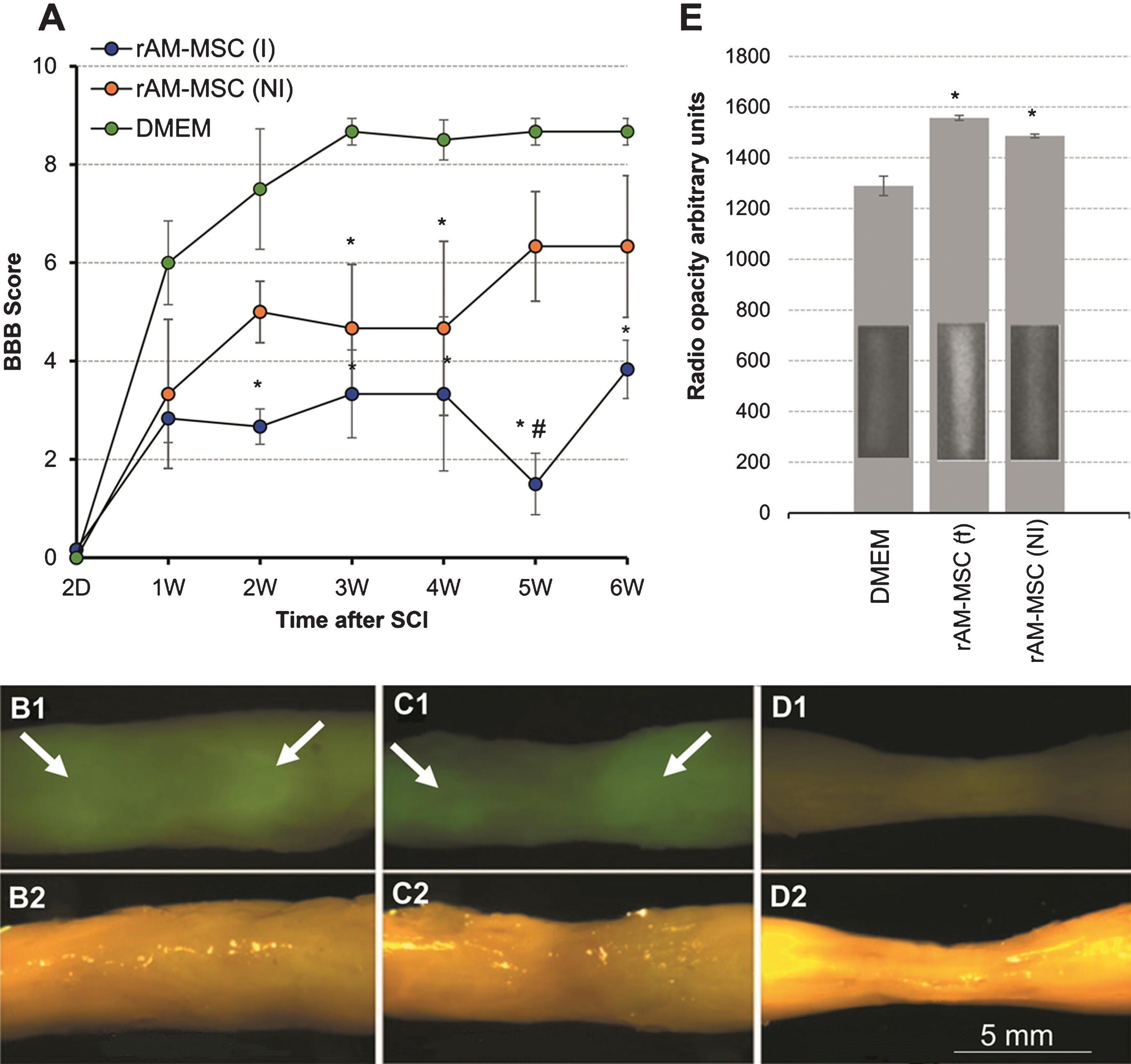
Transplantation of GFP+ rAM-MSC after SCI. BBB scores (A) of SCI rats injected with DMEM as a vehicle, or with GFP+ rAM-MSC that had been induced (I) or not induced (NI) in culture. By comparison to the DMEM SCI controls without cells, the non-induced rAM-MSC (NI) had lower scores, and the induced rAM-MSC (I) had much lower scores. All rats (n = 3/group) received daily injections of CsA. Values are means±Standard Error of Mean, in one-way ANOVA (P < 0.05), * represents significance for DMEM vs. rAM-MSC (I) and rAM-MSC (NI) groups and # for rAM-MSC (I) vs. rAM-MSC (NI). Imaging of whole spinal cords with a stereo zoom fluorescence microscope 6 weeks after SCI and injection with rAM-MSC (I) (B1), rAM-MSC (NI) (C1), and DMEM (D1). Corresponding bright fields images (B2-D2) of the same spinal cords are shown below each fluorescence image. Note tissue shrinkage at the SCI epicenter with control DMEM injection (D2). Radiographs (E) of whole rat spinal cords showed highest average intensities when transplanted with rAM-MSC (I) and progressively lower intensities with rAM-MSC (NI), and saline controls. Representative radiographs of the spinal cords are shown as insets in each bar. Mean intensity values are shown in arbitrary units±Standard Error of Mean.
After 6 weeks, fixed spinal cords dissected from rats injected with rAM-MSC felt much more rigid and appeared more opaque in regions in and around the SCI site than with the SCI controls. Green fluorescent imaging of whole cords showed signals in the rAM-MSC injected cords indicating persistence of GFP+ cells (Fig. 1B1 and C1). Strong GFP fluorescence (arrows) was observed in rostral and caudal regions surrounding the injury epicenter by comparison to the weaker signals in the epicenter, suggesting poorer rAM-MSC survival in the injury epicenter and/or migration of rAM-MSC away from the epicenter. The rAM-MSC injected spinal cords showed little (C1) or no (B1) shrinkage in the injury site, which was obvious with the media controls due to tissue loss.
To analyze spinal cord rigidity by an objective method, we obtained X-ray images of cords to compare their radio opacity. Cords from rats injected with rAM-MSC gave stronger X-ray signals than saline injected cords in and around the SCI epicenter, indicating higher tissue density as found in dense ECM or cartilage (Fig. 1E inserts in the bar graph). Quantitation indicated that the induced rAM-MSC group had the highest X-ray intensities and the non-induced AM-MSC group had lower signals (Fig. 1E), both of which were greater than the vehicle controls. Injections with an immortalized neural stem cell clone into SCI had even lower levels of radio opacity than the SCI control (data not shown) because it suppressed expression of CS56+ ECM by comparison to control SCI (Hasegawa et al., 2005), less ECM results in lower opacity.
To analyze the effects of CsA, we injected the induced rAM-MSC into rat SCI with and without CsA treatment (Fig. 2A). Beginning at 2 weeks after SCI, rats without CsA treatment had an average BBB score of almost 6, which is higher than the CsA-treated group, which plateaued between 3-4. To explore whether these differences are reflected by changes in the tissues, sections prepared 6 weeks after SCI and injections were analyzed with Masson’s Trichrome staining, which shows ECM as blue deposits, cytoplasm as red/pink and nuclei in purple. Injections with media as controls in SCI showed a central lesion cavity, tissue shrinkage, dramatic loss of grey matter, and partial loss of white matter surrounding the injury site (Fig. 2B) similar to that described previously (Hill, Beattie, & Bresnahan, 2001). In spinal cords injected with rAM-MSC, the epicenter was filled with blue stain indicating ECM that extended into rostral and caudal regions of the cord (Fig. 2C and D) but was not observed in control SCI (Fig. 2B).
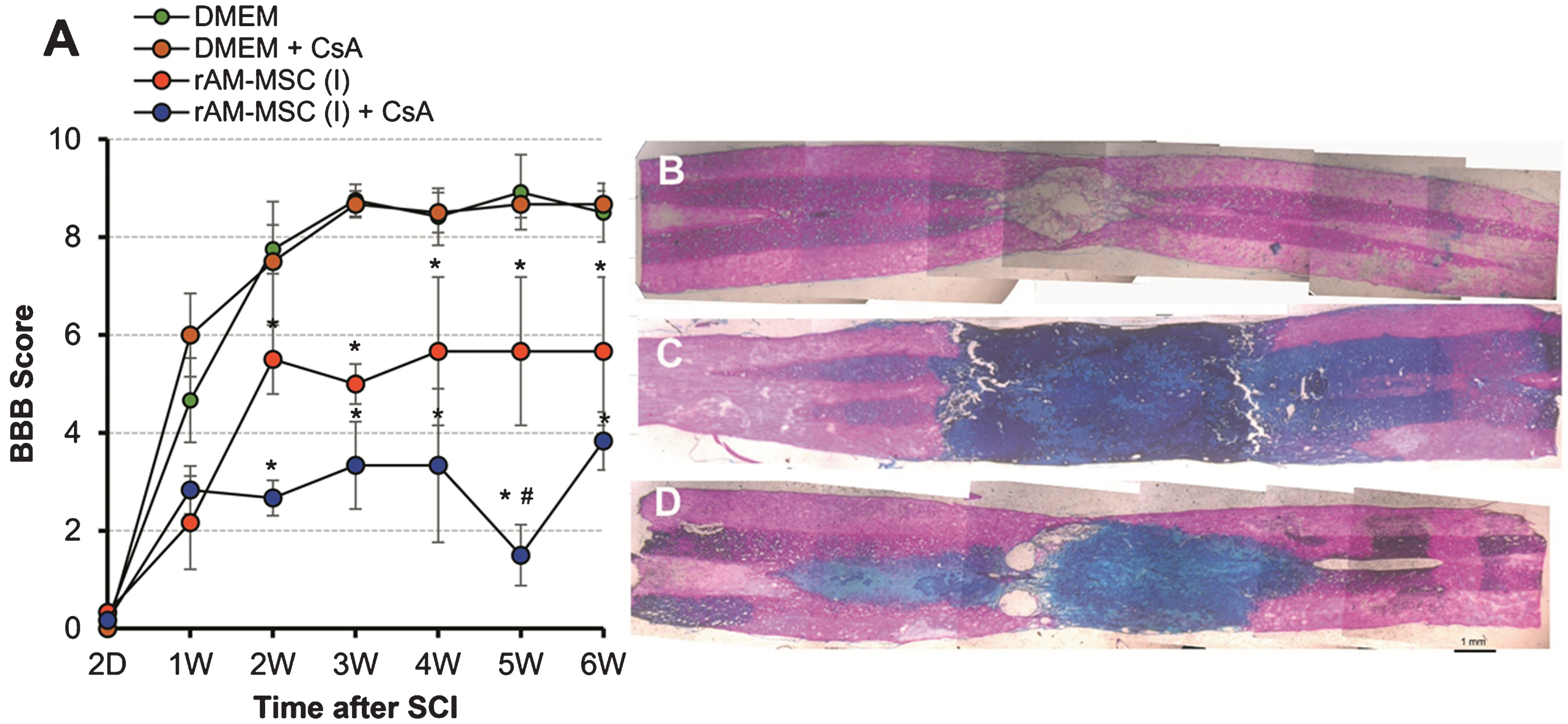
Effect of immunosuppression with CsA on transplantation of rAM-MSC after SCI. Behavioral recovery over 6 weeks (A) using BBB scoring for SCI groups injected with DMEM vehicle alone (green) or with CsA (brown); and injected with rAM-MSC alone (red) or with CsA treatment (blue). Representative tiled microphotographs (B-D) were taken of spinal cord sections centred around the injury site after Masson’s Trichrome staining from rats in panel A injected with DMEM with CsA treatment (B), rAM-MSC without CsA treatment (C) or rAM-MSC with CsA treatment (D). Note minimal ECM (blue) in the SCI epicenter (B) by comparison to the extensive ECM staining along the rostro-caudal axis (C, D) of the cords injected with rAM-MSC, with the most intense ECM staining in rAM-MSC rats that were treated with CsA. Scale bar = 1 mm.

Effects of TGF-β on Expression of Aggrecan in rAM-MSC and rBM-MSC. Induced cultures of rAM-MSC and rBM-MSC were treated with or without TGF-ß and stained by immunofluorescence for aggrecan. The cells were visualized for their expression of GFP (green) and aggressing (red). Note the more intense aggrecan staining of the rAM-MSC by comparison to the weak signal for rBM-MSC.
In the CsA treated rats, ECM completely filled the SCI site with no apparent sparing of white matter (Fig. 2C) and without CsA there was less intense staining for ECM and very small amounts of white matter sparing (Fig. 2D). The rostro-caudal extent of the intense blue stain (Fig. 2C) was approximately the same as that for the fluorescence in the same spinal cord (Fig. 1B1), suggesting that the injected cells deposited ECM. The higher intensity of the ECM stain with CsA treatment may be due to immune protection of the transplanted cells. By comparison to the spared white matter in control SCI (Fig. 2B), progressively less sparing was observed with injections of rAM-MSC without (Fig. 2D) and with CsA (Fig. 2C) treatment. Similarly, by comparison to the BBB scores in control SCI, the scores were lower with injections of rAM-MSC without CsA treatment and lowest with CsA treatment (Fig. 2A). This relationship between the degree of white matter sparing and BBB scores is consistent with previous studies (Basso, Beattie, & Bresnahan, 1996).
The robust ECM deposits observed after transplant or rAM-MSC in outbred Sprague Dawley rat SCI appears to be related to the relative survival of the cells (Figs. 1 and 2). In contrast, survival of syngeneic GFP+ adult BM-MSC decreased in inbred Fisher rat SCI over 6 weeks and was accompanied by dramatically lower levels of CS56+ ECM in SCI sites. Moreover, levels of CS56+ ECM in the adult BM-MSC injected SCI sites were much lower than in rat SCI sites at 4–6 weeks after SCI without cell injections (Iseda et al., 2008) (see Supplemental Figure 1), suggesting that adult BM-MSC express little or no CS56+ ECM and appear to suppress their deposition. Thus there is a major difference between the E18.5 derived rAM-MSC, which deposited large amounts of ECM and the adult BM-MSC, which was associated with lower ECM levels than control SCI.
In an attempt to understand why rAM-MSC (Fig. 2C, D) but not rBM-MSC (Supplemental Figure 1) caused huge ECM deposits in SCI, we compared the effect of TGF-β treatment on these cells in culture. TGF-β is dramatically upregulated after SCI (Carmel et al., 2001; McTigue, Popovich, Morgan, & Stokes, 2000), remains at high levels for at least 8 weeks (Fu et al., 2017), and it has been found to promote chondrocyte differentiation and increased synthesis of ECM including aggrecan in AM-MSC (Mehlhorn et al., 2006; Wei et al., 2009). Immunostaining of MSC cultures with antibodies to aggrecan showed that in response to TGFβ there was more intense aggrecan staining in rAM-MSC than in rBM-MSC. The results suggest that deposition of large amounts of ECM in SCI by rAM-MSC but not by rBM-MSC may be due to differentiation of rAM-MSC along a chondrocytic lineage in vivo.
Here we describe unexpected effects of acute transplantation of neonatal AM-MSC after rat SCI. Previous SCI studies using MSC isolated primarily from adult bone marrow yielded variable degrees of behavioral improvement. However, few studies analyzed survival of the transplanted MSC and those that did found that allografts survived transiently and xenografts survived very poorly (Martinez, Goulart, Ramalho Bdos, Oliveira, & Almeida, 2014; Tetzlaff et al., 2011). In an attempt to maximize survival, we tested newborn GFP+ rAM-MSC allografts with CsA and found robust rAM-MSC survival in the SCI site at 6 weeks. Surprisingly, rAM-MSC transplants resulted in significantly lower BBB scores than in control SCI. The results suggest that after clonal expansion of rAM-MSC they differentiated aberrantly in traumatic sites of SCI into chondrocyte-like phenotypes, which expressed large amounts of ECM.
The first indication of unusual effects of the rAM-MSC was BBB scores lower than controls as early as 2 weeks after SCI and cell transplant. After sacrifice at 6 weeks, the spinal cords around the SCI site were harder and more X-ray opaque than controls. Whole mount imaging revealed a lack of gross tissue loss around SCI sites when transplanted with GFP+ rAM-MSC, and the robust GFP+ fluorescence suggests survival and perhaps expansion of GFP+ rAM-MSC. Histological analyses of Mason’s Trichrome stained tissue sections revealed intense staining for ECM throughout the GFP+ areas. The overlap between the ECM and GFP+ distributions suggests that the GFP+ rAM-MSC transplants produce the ECM but we cannot rule out contributions of ECM from the host spinal cord. In contrast, adult BM-MSC appeared to inhibit ECM deposition from the host spinal cord (Supplemental Figure 1).
Differentiation in culture of the rAM-MSC indicated that they expressed appropriate MSC markers for chondrogenic, osteogenic and adipocitic differentiation (Marcus, Coyne, Rauch, et al., 2008). After in utero injection into brain ventricles the GFP+ rAM-MSC migrated into different brain regions and did not show expansion or unusual differentiation postnatally out to 2.5 months (Marcus, Coyne, Black, et al., 2008). AM-MSC have a greater propensity for chondrogenic differentiation in response to TGF-β in vitro than MSC derived from other tissues. We found that when rBM-MSC and rAM-MSC were treated with TGF-β in culture, aggrecan expression was increased to a greater extent in rAM-MSC than in rBM-MSC. Adult adipose derived MSC had a weaker chondrogenic response to TGF-β than BM-MSC (Hennig et al., 2007).
rAM-MSC transplanted in SCI are exposed to various factors that are upregulated in SCI including TGF-β, which increases dramatically within 6 hours after rat spinal cord contusion (Carmel et al., 2001) and remains elevated for at least 7 days (De Biase et al., 2005). In human SCI, TGF-β is also upregulated as early as 2 days after SCI and it persists for at least 4 months (Buss et al., 2008). Considering that BMPs and TGF-β activate common receptors (Cao & Chen, 2005), it is noteworthy that transplants of hAM-MSC in abdominal muscle differentiated into chondrocytes but only when accompanied by BMP-2 (Wei et al., 2009). Thus chondrogenic differentiation of rAM-MSC in SCI may be due at least in part to upregulation of TGF-β.
The majority of previous studies of MSC in SCI have derived cells from adult bone marrow, which showed transient survival but many studies did not report information on survival (Martinez et al., 2014; Tetzlaff et al., 2011). By comparison to allogenic MSC transplants, there appears to be even weaker survival of xenogenic transplants after injection into rat SCI sites of human BM-MSC (Tetzlaff et al., 2011). Human AM-MSC (Gao et al., 2014; Zhou et al., 2016) and UCBC (Park et al., 2012; Zhilai et al., 2016) also showed weak survival in adult rat SCI sites. This is the first report of robust survival and probable cell expansion of rAM-MSC after transplant in rat SCI even in the absence of immunosuppression by CsA. We discuss two possible explanations for these puzzling results and their potential implications.
One possibility is that there are immune and inflammatory differences between adult BM-MSC and neonatal AM-MSC. Antibodies can be generated against allogeneic adult MSC (Ankrum, Ong, & Karp, 2014), which may play a role in more rapid disappearance of adult BM-MSC than AM-MSC. AM-MSC secrete many anti-inflammatory cytokines in response to inflammation, which may promote their survival, but their responses have yet to be compared with BM-MSC to determine whether they provide a survival advantage in SCI.
A second and perhaps more likely possibility is that neonatal AM-MSC respond more robustly than BM-MSC to certain factors, i.e. TGFß, in promoting chondrogenesis (Syva, Ampon, Lasimbang, & Fatimah, 2017). Thus, the extensive deposition of ECM by the AM-MSC in SCI is consistent with their strong chondrogenic response, in contrast to BM-MSC isolated from bone marrow stroma with little collagen (Bianco, 2014). In response to the rapid and prolonged upregulation of TGFß for at least 8 weeks after SCI (Carmel et al., 2001; Fu et al., 2017), AM-MSC but not BM-MSC are expected to become chondrogenic and assemble ECM. AM-MSC integrate within a 3 dimensional matrix, which serves as a niche where they can survive and continue to differentiate (Bilic et al., 2005). This process could account for what appears to be co-expansion of the AM-MSC within an extensive ECM.
Although this study used cloned rAM-MSC, they are not homogeneous insofar as a sub-population did not survive with CsA treatment. It would be interesting to determine whether other rAM-MSC clones also survive and secrete ECM after transplantation in rat SCI, but that would be extremely laborious and may not provide definitive results. However, we are not aware of any previous reports of neonatal allogenic rAM-MSC transplants in rat SCI sites. Thus, studies are needed to test whether allogenic newborn MSC including those derived from amnion and placenta survive better than adult MSC (e.g. BM-MSC) in SCI sites, and if so, whether they differentiate aberrantly. Because the interpretation of the effects of CsA on the role of immune compatibility of AM-MSC is not straight forward, it would be interesting to test aberrantly differentiating rAM-MSC in euthymic rats but these are difficult to work with for SCI and only one study has reported so far (Jin et al., 2011).
There is concern that extensive expansion of more plastic newborn MSC may allow amplification of sub-populations of cells that can expand and differentiate aberrantly as we observed in SCI, but not in the normal fetal CNS in which they migrated and survived only transiently (Marcus, Coyne, Black, et al., 2008). In this regard it is noteworthy that extensively expanded human neural stem/precursors had reduced efficacy after injection into SCI sites in rodents, and a clinical trial with these cells showed no improvement and was stopped recently (Servick, 2017).
Clinical development requires extensively expanded banks of MSC such as those prepared for allogenic transplants from adult bone marrow (Mesoblast), which is an effective clinical treatment for Graft vs. Host Disease, and from human newborn placenta (Pluristem). Because the altered milieu in SCI sites can affect cell survival and differentiation, it is important to demonstrate safety of cells in SCI models, which has been demonstrated for adult rBM-MSC and serves as a basis for translational studies (Tetzlaff et al., 2011).
Conclusion
This study raises concerns that clonal or highly expanded MSC derived from newborn tissues may yield sub-populations of cells that after transplantation into SCI sites survive, expand and differentiate aberrantly in response to SCI-induced changes in expression of morphogenetic factors. Future studies need to verify the safety and efficacy of injecting allogenic neonatal MSC into traumatic sites of SCI in rodent models before proceeding to the clinic.
Disclosure
There are no conflicts of interest.