
Abstract
Background:
When nerve transection is performed on adult rodents, a substantial population of neurons survives short-term disconnection from target, and the immune system supports this neuronal survival, however long-term survival remains unknown. Understanding the effects of permanent axotomy on cell body survival is important as target disconnection is the first pathological occurrence in fatal motoneuron diseases such as amyotrophic lateral sclerosis (ALS) and spinal muscular atrophy (SMA).
Objective:
The goal of this study was to determine if facial motoneurons (FMN) could survive permanent target disconnection up to 26 weeks post-operation (wpo) after facial nerve axotomy (FNA). In addition, the potentially additive effects of immunodeficiency and motoneuron disease on post-axotomy FMN survival were examined.
Methods:
This study included three wild type (WT) mouse strains (C57BL/6J, B6SJL, and FVB/NJ) and three experimental models (RAG-2–/–: immunodeficiency; mSOD1: ALS; Smn–/–/SMN2+/+: SMA). All animals received a unilateral FNA, and FMN survival was quantified at early and extended post-operative timepoints.
Results:
In the C57BL/6J WT group, FMN survival significantly decreased at 10 wpo (55±6%), and then remained stable out to 26 wpo (47±6%). In the RAG-2–/– and mSOD1 groups, FMN death occurred much earlier at 4 wpo, and survival plateaued at approximately 50% at 10 wpo. The SMA model and other WT strains also exhibited approximately 50% FMN survival after FNA.
Conclusion:
These results indicate that immunodeficiency and motoneuron disease accelerate axotomy-induced neuron death, but do not increase total neuron death in the context of permanent target disconnection. This consistent finding of a target disconnection-resilient motoneuron population is prevalent in other peripheral nerve injury models and in neurodegenerative disease models as well. Characterization of the distinct populations of vulnerable and resilient motoneurons may reveal new therapeutic approaches for injury and disease.
Keywords
Introduction
Experimental models of peripheral nerve injury have been used for decades to understand cellular responses to injury and to identify new therapeutic approaches for neuroregeneration. Functional recovery from nerve injury requires survival of the neuron cell body, regrowth of its axon, and reconnection to a target, with neuron survival being the first component necessary for success (Hart, Terenghi, & Wiberg, 2008). Study of axotomized motoneurons (MN) is especially important given their exquisite reciprocal relationship with muscle, as both cell populations provide trophic support for each other’s survival, and disruption of this connection can lead to muscle atrophy and MN death.
Factors that impact MN survival after target disconnection include species, age, severity of the injury, and distance of the injury from the cell body (Gu, Spasic, & Wu, 1997; Hart et al., 2008; Moran & Graeber, 2004). In addition, our laboratory has determined that immune status also affects facial motoneuron (FMN) survival after facial nerve axotomy (FNA), in which nerve transection results in permanent target disconnection. Comparing wild type (WT) to the severe combined immunodeficient (scid) mouse model reveals that the immune system supports FMN survival at 4 weeks post-operation (wpo), but this neuroprotective effect is temporary, and FMN survival falls at 10 wpo, correlating with down-regulation of the immune activation (Serpe, Sanders, & Jones, 2000). This transient support only applies to approximately half of the FMN population; the other half of FMN are resilient to target disconnection regardless of immune status.
Target disconnection is also a contributing factor in the MN disease amyotrophic lateral sclerosis (ALS), as loss of neuromuscular junctions is the earliest observed pathology, followed by retrograde axonal die-back that ultimately results in death of the MN cell body (Fischer et al., 2004). In the mutant superoxide dismutase 1 (mSOD1) mouse model of ALS, the disease process first affects lumbar MN, resulting in a total of 50% lumbar MN loss by endstage at 18-19 weeks of age (woa, Chiu et al., 1995). The MN disease process in the mSOD1 mouse ascends rostrally, and FMN loss is first detected at 16 weeks of age (Haulcomb et al., 2014). When FNA is superimposed on mSOD1 mice in the presymptomatic stage of disease at 8 woa, FMN death is observed earlier than WT at 4 wpo and remains stable at 8 wpo, reflecting a similar temporal pattern of target disconnection-induced MN death as observed in immunodeficient animals (Haulcomb et al., 2014). Together, these studies suggest that there exists a MN population that is resistant to both disease- and axotomy-induced MN death in mSOD1 mice.
Spinal muscular atrophy (SMA) is a genetic MN disease caused by a lack of functional survival motoneuron (SMN) protein, and the disease pathology also consists of an axonal die-back process (Udina et al., 2017). The Smn–/–/SMN2+/+ mouse model of SMA recapitulates the human disease phenotype and has a lifespan of 9 – 15 months, and this transgenic mouse was included in this study as a second model of MN disease. At 12 months of age, there is about 60% MN survival in the lumbar spinal cord as a result of the disease process, again demonstrating a disease-resilient MN population (Tsai, Tsai, Lin, Hwu, & Li, 2006). Only one study thus far has examined how an SMA mouse model responds to peripheral nerve injury using the Smn+/– SMA transgenic mouse model, which is the mildest model of SMA disease. Sciatic nerve axotomy of this model revealed no significant difference in axotomy-induced MN death (about 10% loss relative to uninjured control side) in the Smn+/– SMA model at 8 and 16 wpo relative to WT (Udina et al., 2017). There is evidence in the SMA disease model that there are both a disease- and axotomy-resilient MN population, mirroring the findings in the mSOD1 ALS mouse model.
This persistent finding of neurons resilient to target disconnection has been documented in a plethora of publications reporting on other neuronal populations using a variety of peripheral nerve injury models (Hart et al., 2008). This study advances the field by addressing two questions: first, what are the long-term effects of permanent target disconnection on FMN survival, and second, how does immunodeficiency or MN disease further affect FMN survival after axotomy? To accomplish this goal, FMN survival after FNA was assessed in three WT strains: C57BL/6J, B6SJL, and FVB/NJ, and three experimental groups: the recombinase-activating gene-2 knockout model of immunodeficiency (RAG-2–/–) on the C57BL/6J background, the mSOD1 on B6SJL background, and the Smn–/–/SMN2+/+ (SMA) on FVB/NJ background. Our data consistently reveal that approximately 50% of the FMN population is resilient to target disconnection, regardless of mouse strain, immune status, or concurrent MN disease. Further applications of this facial nerve injury model can lead to identification of neuroprotective factors that could be clinically applicable in the contexts of both traumatic nerve injury and neurodegenerative disease.
Materials and methods
Animals and surgical procedure
All animal handling and procedures were in compliance with the National Institutes of Health (NIH) guidelines and were approved by Indiana University School of Medicine’s Institutional Animal Care and Use Committee. The following strains of mice were purchased from The Jackson Laboratory (Bar Harbor, ME): C57BL/6J (C57BL/6J WT, RRID:IMSR_JAX:000664), B6(Cg)-Rag2tm1.1Cgn/J (RAG-2–/–, RRID:IMSR_JAX:008449), B6SJL F1/J (B6SJL WT, RRID:IMSR_JAX:100012), B6SJL-Tg(SOD1*G93A)1Gur/J (mSOD1, RRID:IMSR_JAX:002726). The FVB/NJ and Smn–/–/SMN2+/+ SMA mouse strains were bred in-house and graciously provided by Drs. E.J. Androphy and S.K. Custer. These mice strains are also available through The Jackson Laboratory (FVB/NJ, RRID:IMSR_JAX:001800; FVB.Cg-Smn1tm1Hung Tg(SMN2)2Hung/J, RRID:IMSR_JAX:005058). Normal, non-ovariec-tomized female mice were exclusively used in this study because male mice exhibit significant aggressive behavior post-operatively and require individual housing, which is detrimental to animal welfare.
FNA was performed following previously published methods on 8 week old mice using aseptic technique (Serpe et al., 2000). In brief, the trunk of the facial nerve was exposed at its exit from the stylomastoid foramen and completely transected, and the stumps were separated to prevent reconnection. Right facial paralysis was confirmed by absent vibrissae movement and null eyeblink reflex to air puff stimuli. In all axotomized animals, no recovery of facial motor function was observed throughout the experimental timecourse. The left facial nerve was not operated on, allowing for the left facial motor nucleus to serve as a paired internal control.
Facial motoneuron quantification and statistical analysis
At the appropriate experimental endpoint (4, 6, 10, 18, or 26 wpo; n = 4–8/group/timepoint), animals were euthanized via CO2 inhalation and cervical dislocation, and the brain was removed and flash frozen. Tissue processing, thionin staining, and facial motoneuron counting procedures were executed following previously published protocols (Serpe et al., 2000). One-way ANOVA and Tukey’s multiple comparisons tests, two-way ANOVA and Sidak’s multiple comparisons test, and unpaired t-tests were performed using Graphpad version 7.03.
Results
Long-term survival of axotomized facial motoneurons in C57BL/6J WT and immunodeficient RAG-2–/– mice
To determine the effects of time on axotomized FMN survival, 8 week old C57BL/6J WT mice received FNA, and FMN quantification was performed at 4, 10, 18, and 26 wpo. In the C57BL/6J WT, FMN survival was not significantly decreased relative to the control facial motor nucleus at 4 wpo (89±2%). Significant FMN loss relative to control facial motor nucleus was observed at 10, 18, and 26 wpo (55±6%, 44±5%, 47±6%, respectively, p < 0.001, for each), and no significant differences were observed between 10–26 wpo timepoints (Fig. 1A). Thus, a plateau in the WT group was reached by 10 weeks.
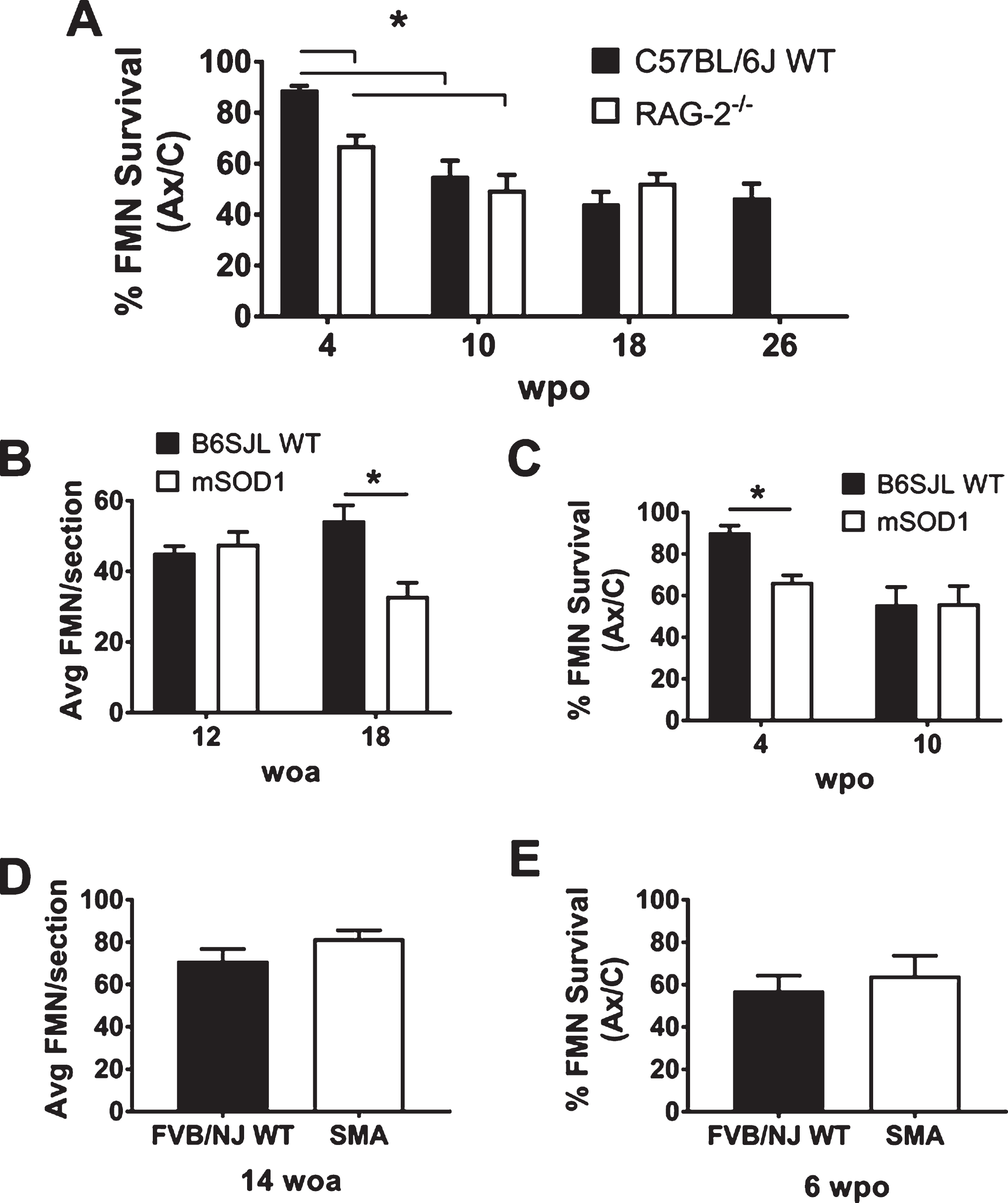
Quantification of FMN survival in WT and experimental groups. A: Average percent FMN survival±SEM comparing the axotomized (Ax) to uninjured control (C) facial motor nucleus at 4, 10, 18, and 26 weeks post-operation (wpo). B: Average FMN counts±SEM per 25μm section of the uninjured control facial motor nucleus at 12 and 18 weeks of age (woa). C: Average percent FMN survival±SEM, comparing the Ax to C facial motor nucleus at 4 and 10 wpo, corresponding with 12 and 18 woa, respectively. D: Average FMN counts±SEM per 25μm section of the uninjured control facial motor nucleus at 14 woa. E: Average percent FMN survival±SEM, comparing the Ax to C facial motor nucleus at 6 wpo, corresponding with 14 woa. Symbols used: *, p < 0.05.
To identify if immunodeficiency had an added impact on long-term FMN survival after FNA, RAG-2–/– mice were next examined. Relative to the uninjured control facial motor nucleus, FMN survival was significantly decreased at 4 wpo (67±4%, p < 0.001) and continued to decrease out to 10 wpo in RAG-2–/– mice (50±6%, p = 0.04). FMN survival levels remained stable at 18 weeks after axotomy (53±4%), with no significant difference relative to 10 wpo (Fig. 1A).
In comparing the two groups, RAG-2–/– mice exhibited significantly earlier FMN loss relative to C57BL/6J WT at 4 wpo (p = 0.007), and there were no differences detected at later timepoints. The findings in the RAG-2–/– group reflect what was observed in a previous study of the Balb/c WT and RAG-2–/– on Balb/c background (Serpe et al., 2000). Prior work on FMN survival after FNA has only gone as far as 10 wpo, in which immunodeficient scid mice exhibit comparable FMN survival relative to the C.B-17 ICR WT strain (Serpe et al., 2000). Together, these findings suggest that there is a population of FMN whose early, transient survival depends on an intact immune system, and a second population of FMN exists for which immune status has no effect.
The impact of concurrent MN disease on long-term FMN survival after FNA was next examined. In the mSOD1 mouse model of ALS, hindlimb motor dysfunction develops at 13 weeks of age (woa), and the mouse is moribund due to paralysis at approximately 18-19 woa (Chiu et al., 1995). In this study, FNA was performed in the pre-symptomatic stage at 8 woa, and animals were euthanized at 4 wpo (12 woa) and 10 wpo (18 woa), just before endstage. The B6SJL WT was included as a strain-matched control.
First, disease-induced MN loss was measured by quantifying FMN in the control facial motor nucleus in both B6SJL WT and mSOD1 groups. At 12 woa, still within the presymptomatic stage, there is no significant loss of FMN in the mSOD1 group relative to B6SJL WT (48±3 vs. 45±2 FMN/section). At 18 woa, near endstage, there is a significant disease-induced loss of FMN in the mSOD1 group relative to B6SJL WT (33±4 vs. 54±4 FMN/section, p = 0.003, Fig 1B).
Second, the effect of superimposing FNA on mSOD1 mice on FMN survival was quantified at early and late timepoints. In the B6SJL WT, FMN survival was not significantly different from the control facial motor nucleus at 4 wpo (90±3%), and a significant drop in FMN survival was observed at 10 wpo (56±8%, p < 0.001; Fig. 1C). In the mSOD1 group, a significant decrease in FMN survival relative to the unoperated control side was observed at 4 and 10 wpo (67±3% and 56±8%, p = 0.004 and 0.0003, respectively), and there was no statistically significant difference between 4 and 10 wpo (Fig. 1C). The mSOD1 group had a significant decrease in post-axotomy FMN survival at 4 wpo relative to B6SJL WT (p = 0.02). There was no difference in FMN survival between these groups at 10 wpo, suggesting that the resilient MN population remains unaffected by concurrent MN disease and target disconnection.
FMN survival in a second model of motoneuron disease
To further explore the effects of MN disease on FMN survival after FNA, the Smn–/–/SMN2+/+ mouse model of SMA was also included in this study. This specific SMA mouse model develops motor symptoms at 36 woa, and lifespan ranges from 36–60 woa, which is a similar lifespan as the strain-matched FVB/NJ WT (Tsai et al., 2006). For this study, FNA was performed within the presymptomatic stage at 8 woa, and the 6 wpo timepoint (14 woa) was selected as an intermediary between 4 and 10 wpo used for previous groups in an effort to conserve animals.
When the effects of SMA disease on FMN survival was quantified at 14 woa, there was no significant difference in the SMA group relative to the FVB/NJ WT (82±4 vs. 71±6, respectively, Fig. 1D). After FNA, the SMA group also had equivalent FMN survival compared to FVB/NJ WT at 6 wpo (64±5% vs. 57±3%, respectively, Fig. 1E). These findings indicate that the SMA MN disease does not augment axotomy-induced FMN death after FNA, similar to the findings observed in the mSOD1 mouse model. This result reinforces the evidence for a MN population that is resilient in settings of both MN disease and axotomy-induced target disconnection.
Discussion
This study examined the long-term effects of axotomy-induced target disconnection on MN survival and determined if immunodeficiency or MN disease further affected survival. The first important finding from this study is that there is a MN population that survives permanent target disconnection for a prolonged length of time. Furthermore, this resilient population is persistent in the contexts of both immunodeficiency and MN disease. This study is also significant because, to our knowledge, this is the first study presenting post-axotomy MN survival data on the Smn–/–/SMN2+/+ mouse model of SMA, with the only other available data being from a sciatic nerve injury on the C57BL/6-Smn+/– model, which also found axotomy- and disease-resilient MN (Udina et al., 2017).
This finding of a long-term resilient population of neurons after axotomy is apparent in other neuronal types as well (Hart et al., 2008). For example, 65% of sensory neurons survive sciatic nerve transection at 24 wpo (McKay Hart, Brannstrom, Wiberg, & Terenghi, 2002), 70% of motoneurons survive C7 spinal nerve transection at 16 wpo (Ma, Novikov, Wiberg, & Kellerth, 2001), and 65% of hypoglossal motor neurons survive axotomy at 52 wpo (Snider & Thanedar, 1989). The only known circumstances that result in near-total neuronal death after axotomy occur with nerve avulsion injuries or when axotomy is imposed on neonatal animals (Catapano et al., 2017; Gu et al., 1997).
The presence of axotomy-vulnerable vs. -resilient FMN challenges the traditional understanding of neuronal physiology, in that trophic factors from target are required for cell body survival. From developmental and functional perspectives, FMN have been historically considered to be homogenous (Moran & Graeber, 2004). However, the results of this study clearly indicate the presence of two populations of FMN with diametrically-opposed responses to target disconnection in both WT and disease mouse models. Future work will focus on the elucidation of underlying mechanisms that define the fate-determining intrinsic steps occurring in the two distinct motoneuron pools. Previous studies determined that axotomy-induced FMN death is asymmetrically distributed throughout the facial motor nucleus, and careful stereotactic analysis may identify individual vulnerable or resilient MN (Canh, Serpe, Sanders, & Jones, 2006). Identification of these different neuronal populations within the seemingly-homogeneous facial motor nucleus may lead to development of biomarkers that can be applied in neuroregenerative research. Extrinsic influences from microglia and astrocytes in the CNS or Schwann cells in the residual nerve will also be evaluated, as these cell responses to nerve injury are also heterogeneous and may differentially promote MN survival (Brushart et al., 2013; Liddelow et al., 2017; Mesnard, Alexander, Sanders, & Jones, 2010; Morganti, Riparip, & Rosi, 2016; Scheib & Hoke, 2013). Identifying and harnessing the key mechanisms behind neuronal resiliency in axotomy and disease will significantly advance the development of therapies for patients.
Footnotes
Acknowledgments
This work was supported by a research grant from the National Institutes of Health (NS40433; KJJ and VMS). We thank Drs. E. J. Androphy and S. K. Custer for generously providing the SMA mice.