
Abstract
Background:
In this proof-of-concept paper, we investigated whether combination treatment with progesterone (P4) and chloroquine (CQ) would reduce ischemic injury more effectively than either agent alone in a transient middle cerebral artery occlusion (tMCAO) model in male rats.
Methods:
P4 (8 mg/kg) and CQ (25 mg/kg) were given alone or in combination beginning at different times during surgery and for 3 days post-occlusion. Locomotor activity and grip strength were evaluated as measures of impairment and recovery. Infarct size was assessed by TTC staining. Markers of autophagy (LC3 and SQSTM1/p62) and apoptosis (Bcl-2 and Bax) were evaluated with western blotting.
Results:
At the doses we employed, the combination was not more effective than either drug given separately on measures of grip strength or locomotor activity. However, combination therapy substantially reduced infarct size, and significantly increased Bcl-2 protein levels and suppressed Bax expression. Progesterone decreased the expression of LC3-II 24 h and SQSTM1/p62 after ischemia.
Conclusions:
Our findings suggest that combination therapy with P4 and CQ is not detrimental and has a small-to-moderate additive neuroprotective effect on ischemic injury in rats without substantively affecting behavioral outcomes. CQ and P4 may help to regulate the expression of both autophagy-related and apoptosis-related proteins.
Introduction
To date, no clinical trial has led to a successful pharmacological, neuroprotective intervention for stroke. The problem may result in part from the fact that stroke causes systemic effects that are not amenable to the modification of a single receptor mechanism or target organ (Stein, 2017; Zhu et al., 2017). A safe and effective therapy that can target multiple pathways to ischemic neuroprotection is urgently needed, even in cases where mechanical clot removal is successful. This is because inflammatory and degenerative processes that accompany the initial injury may not be completely corrected by mechanical clot removal itself. But even a pleiotropic drug is not likely to address all the pathologies following stroke. A recent consensus paper published under the auspices of the NIH proposed that combination therapies may prove more effective in clinical practice than monotherapies (Margulies et al., 2016) and urged that the combinatorial approach needs more attention in pre-clinical investigation.
The neurosteroid progesterone (P4) is a pleiotropic hormone shown to have neuroprotective effects in various models of neuronal injury (Gaignard et al., 2017; Guennoun et al., 2018; Stein, 2017; Zhu et al., 2017). Our laboratory has also published a number of studies demonstrating substantial neuroprotection by P4 in both permanent and transient stroke models (Atif et al., 2013; Ishrat et al., 2010, 2012, 2009; Sayeed et al., 2007; Wali et al., 2016, 2014; Yousuf, Atif et al., 2014; Yousuf, Sayeed et al., 2014). P4 treatment can reduce infarct volume, inflammatory cytokine expression, oxidative damage, apoptosis, and cerebral edema, leading to improved functional outcomes (Ishrat et al., 2012; Ishrat et al., 2009; Sayeed et al., 2007; Wali et al., 2016). However, despite its multiple actions, this does not assure that P4 is a panacea for every mechanism associated with the cascade of cellular injuries that often follow moderate to severe brain injuries, including stroke.
Although a number of agents could be combined with P4 to increase its efficacy, we chose to test a combination of P4 and cholorquine (CQ). We chose CQ for several reasons. A common quinolone derivative, CQ is well known for its clinical use in the treatment of malaria and various cancers (Hall et al., 2018; Monma et al., 2018). It has also been used successfully for decades to treat inflammatory disorders like systemic lupus erythematosus and rheumatoid arthritis (Al-Bari, 2015; Rainsford et al., 2015), but the drug has had more limited use in the experimental treatment of stroke. CQ given prior to insult was reported to inhibit inflammation caused by upregulation of Toll-like receptors (Cui et al., 2013).
CQ is also an inhibitor of autophagy (Kimura et al., 2013) which has been shown to reduce brain damage, decrease infarct size, and improve functional outcome after stroke (Buckley et al., 2014; Cui et al., 2013). There are many papers showing that after a stroke, autophagy is activated and may contribute to ischemic neuronal and/or astrocytic injury in rat models of cerebral ischemia (Descloux et al., 2015; J. Li et al., 2015; Qin et al., 2010; Shao & Liu, 2015; Wen et al., 2008). Autophagy is a lysosome-dependent mechanism by which long-lived proteins, protein aggregates, and damaged organelles are degraded and recycled (Klionsky & Emr, 2000; Levine & Klionsky, 2004) over time. The functional role of autophagy during cerebral ischemia/reperfusion is complex, because its pathophysiological functions depend on the severity and duration of ischemia (hypoxia) (Uberti et al., 2014). Hyperactivation of autophagy can lead to cell death, so autophagy can be either beneficial or detrimental (Jiang et al., 2015), and its role in neuronal survival after transient focal ischemia is incompletely understood.
Both P4 and CQ easily cross the blood-brain-barrier (BBB) and rapidly diffuse throughout the brain (Adelusi & Salako, 1982; Schumacher et al., 2016). P4 and CQ have high safety profiles, act on different pathological injury mechanisms, and are easy to administer and inexpensive. Given the beneficial properties of both agents following cerebral injury, it is reasonable to ask whether, given together with P4, CQ could extend the latter’s neuroprotective effects (or vice versa) and lead to better functional outcomes than either agent given alone. In the current study we investigated whether combinatorial treatment with P4 and CQ could produce better acute stage outcomes than either drug alone after transient cerebral ischemic injury in rats. We evaluated the effects of P4 and CQ on (1) functional recovery (locomotor activity and grip strength); (2) infarct volume (TTC staining); (3) autophagy markers (LC3, SQSTM1/ p62); and (4) apoptosis markers (Bcl-2, Bax).
Materials and methods
Animals and drug administration
Fifty-five adult male Sprague-Dawley rats (320 to 350 gm; Charles River Laboratories, Wilmington, MA, USA) were used according to procedures approved by the Emory University Institutional Animal Care and Use Committee (protocol #2003056). Rats were quarantined for 7 days before the experiment and housed in individual cages with free access to food and water. Room temperature was maintained at 21–25°C, and humidity at 45–50%. The animals were maintained under a 12:12-h reverse light/dark cycle (11:00–23:00 h) so that behavioral testing would occur during their active phase. Baseline behavioral measures were taken 1 day before surgery. Rats were randomly assigned to 5 groups: sham+vehicle (n = 11), transient middle cerebral artery occlusion (tMCAO)+vehicle (n = 11), tMCAO+P4 (n = 11), tMCAO+CQ (n = 11), and tMCAO+P4+CQ (n = 11).
P4 (8 mg/kg, P-0130; Sigma Aldrich, St Louis, MO, USA) was dissolved in 22.5% 2-hydroxypropyl-β cyclodextrin (HBC, Sigma) and administered by intraperitoneal (i.p.) injection for faster absorption at 1 h post-occlusion, and then by subcutaneous (s.c.) injection, for more sustained absorption, at 6, 24 and 48 h post-occlusion. The 8-mg/kg dose and routes of administration for P4 were determined from previous studies in rodents showing that this amount provided the maximal protective effects for stroke (Atif et al., 2013; Sayeed et al., 2006, 2007). Dosing of CQ (25 mg/kg, Sigma) was based on previous reports in the autophagy literature (Cui et al., 2013; P. Li et al., 2018). CQ was dissolved in saline and administered by i.p. injection 10 min before the onset of ischemia, and then by i.p. injection at 24 and 48 h post-occlusion. To keep the study blinded, both sham and tMCAO vehicle groups received saline and HBC.
Transient middle cerebral artery occlusion
Focal cerebral ischemia was induced by the occlusion of the right middle cerebral artery as previously described (Longa et al., 1989). All rats were anesthetized using 5% isoflurane and maintained at 1.5–2% (2:1 nitrous oxide and oxygen) during surgery. A midline incision was made on the ventral surface of the neck, and the right common carotid artery was isolated and ligated with 6.0 silk suture. The internal carotid artery and the pterygopalatine artery were temporarily occluded using a microvascular clip. A 4-0 Doccoltrademark filament (Doccol Corporation, Redlands, CA, USA) was inserted into the right internal carotid artery and advanced approximately 20 mm distal to the carotid bifurcation artery until a gentle resistance was felt. After 120 min of MCAO, the occluding filament was withdrawn back into the common carotid artery to allow reperfusion. Rats were permitted to recover from anesthesia in a pre-warmed cage. Sham-operated rats were subjected only to exposure of the carotid arteries; the monofilament was not inserted.
Physiological parameters were monitored as follows: heart rate was maintained around 300–350 beats per minute and blood oxygen level (SpO2) was kept above 95% (SurgiVetTM model V3304, Waukesha, WI, USA). Core body temperature (37±0.5°C) was maintained with an automated heat lamp (Harvard Apparatus, South Natick, MA, USA) (Wali et al., 2016).
Behavioral testing
Locomotor activity
Digiscantrademark activity-monitoring boxes (Columbus Instruments, Columbus, OH, USA) were used to assay spontaneous motor activity. Each session lasted 5 min and was conducted under red-light conditions in a quiet environment (Wali et al., 2014; Yousuf, Sayeed et al., 2014). Baseline scores were obtained 1 day before surgery and again at 3 days post-tMCAO. The parameters measured were total distance travelled (centimeters) and rest time (seconds), expressed as percent of baseline values.
Grip strength
An electronic digital force gauge grip-strength meter (Columbus Instruments) was used to measure forelimb grip strength as previously described (Ishrat et al., 2010). A digital reading (in Newtons) of three successive trials was obtained for each rat and the best (highest) score was used for data analysis (Wali et al., 2016). Baseline was determined 1 day before surgery. Animals were tested again at 3 days post-tMCAO. The values were expressed as percent of baseline values.
Assessment of infarct size
At 3 days post-stroke, the rats were euthanized and their brains removed immediately and cut into 7 serial coronal sections of 2 mm thickness with a rat brain matrix (Harvard Apparatus). Sliced brain tissues were immersed in a 2% solution of 2, 3, 5-triphenyltetrazolium chloride (TTC, Sigma) in PBS at 37°C for 15 min followed by fixation with 10% formalin neutral buffer solution. Both hemispheres of each stained coronal section were scanned using a high-resolution scanner (Epson Perfection 2400 Photo), and then evaluated by ImageJ software (NIH, Bethesda, MD, USA). Unstained areas (pale color) were defined as ischemic lesions. An edema index was calculated by dividing the total volume of the left hemisphere by the total volume of the right hemisphere. The actual infarct volume adjusted for edema was calculated by dividing the infarct volume by the edema index (Hua et al., 2015; Wali et al., 2012; Won et al., 2014). Infarct volume was expressed as a percentage of the contralateral area for each section.
Western blotting
For western blot analyses, a subset of rats were killed at 24 h after the induction of tMCAO and peri-infarct cortical tissue was processed for protein analysis. Tissues were homogenized in T-per tissue protein extraction (P78510, Pierce Biotechnology, Rockford, IL, USA) containing protease inhibitor cocktail (P8340, Sigma). Homogenates were centrifuged at 10,000× g at 4°C for 10 min and the supernatant was preserved at –80°C. Coomassie blue protein assay (P1856210, Pierce) was performed for protein equalization. Forty-mg protein samples were separated by 4–20% acrylamide Criterion gel (Bio-Rad) at 100 V for 2 h and transferred to a polyvinylidene fluoride membrane at 100 V for 40 min. Nonspecific binding sites of the membrane were blocked with 5% nonfat dry milk in TBS-T (TBS containing 0.05% Tween-20). Membranes were probed with primary antibodies for LC3A/B (mAb #12741, Cell Signaling Technology, Danvers, MA, USA), SQSTM1/p62 (pAb #5114, Cell Signaling), Bcl-2 (sc-7382, Santa Cruz Biotechnology, Santa Cruz, CA, USA), Bax (pAb #2772, Cell Signaling) overnight at 4°C. Membranes were then incubated in horseradish peroxidase-conjugated secondary antibodies (1:5,000; KPL). Beta-actin (1:10,000; Sigma) was probed as a loading control. Blots were developed using a chemiluminescent substrate (Denville Scientific, Holliston, MA, USA) for 3 min. Chemiluminescent bands were detected on Kodak autoradiography film in a dark room, and their densities were measured using ImageJ software (NIH).
Statistics
Results were analyzed with one-way ANOVA followed by Tukey’s post-hoc test. Data are presented as Mean±SE. Calculations were obtained using SPSS 24.0 software. A difference at p < 0.05 was considered significant.
Results
Infarct size reduction by P4 and CQ following cerebral ischemia
ANOVA showed a difference among the treatment groups [F(3,20) = 39.70, P < 0.0001]. Post-hoc analysis showed that treatments with P4 alone, CQ alone and P4+CQ produced a reduction in infarct size (p < 0.05). However, the combination treatment showed a significantly greater reduction in infarct size than either agent given alone (p < 0.05) (Fig. 1).
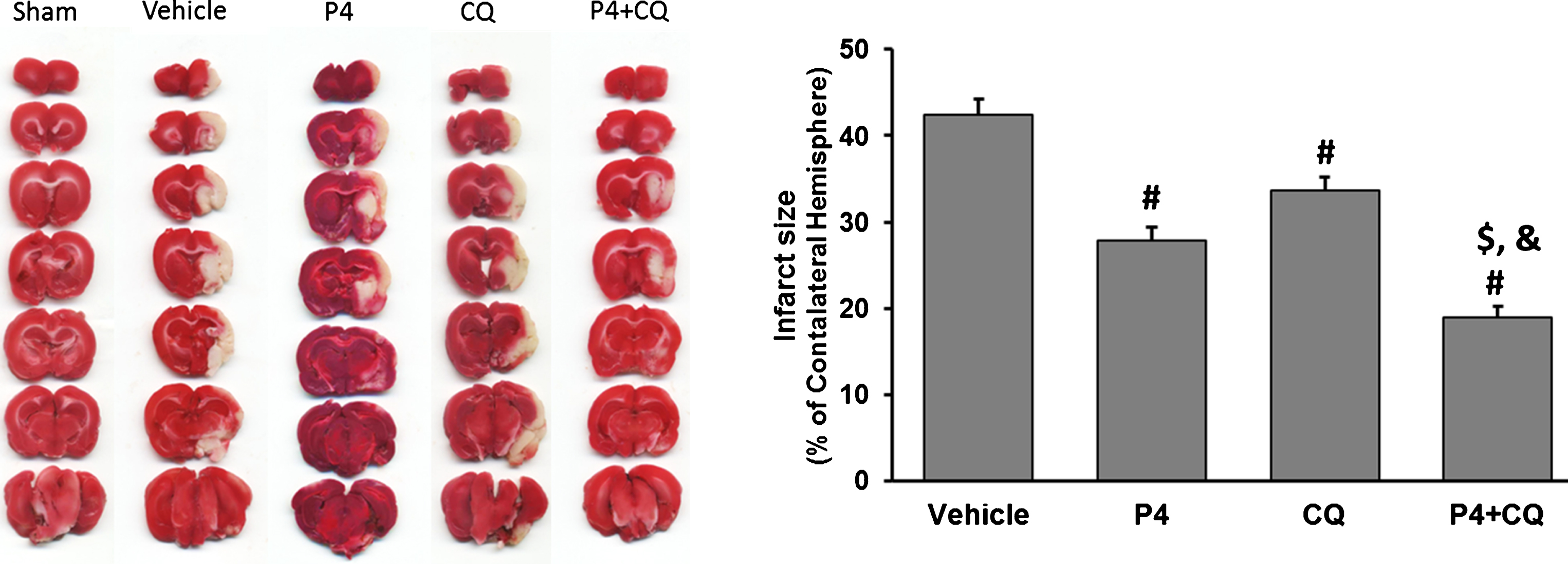
Effect of P4 and CQ alone or in combination on infarct size. Infarct size was reduced by P4 and CQ on Day 3 post-stroke using TTC staining. Values are expressed as Mean±SE. #p < 0.05 compared to vehicle; $p < 0.05 compared to P4; &p < 0.05 compared to CQ. N = 6/per group.
Locomotor activity
Open field activity was measured at 3 days after tMCAO. Total distance travelled showed significant group [F(4,25) = 16.57, P < 0.0001] effects. There was a significant decrease in travel distance in all tMCAO groups compared to shams at 3 days post-stroke (p < 0.05). P4+CQ animals showed significantly greater distance travelled than the vehicle group (Fig. 2A).
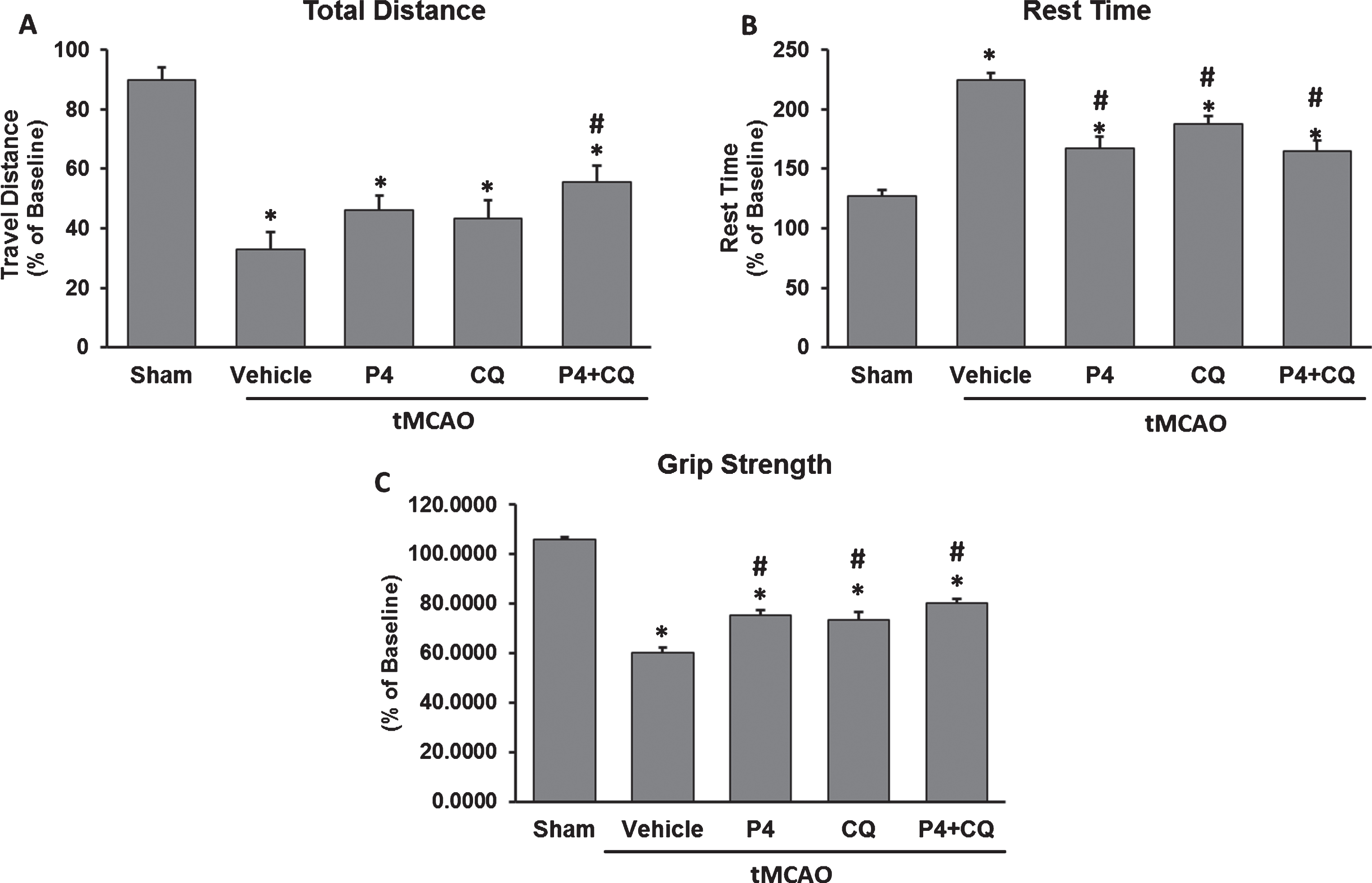
Effect of P4 and CQ alone or in combination on functional outcomes. P4 and CQ improved locomotor activity (A & B) and grip strength (C) on Day 3 post-injury. Values are expressed as Mean±SE. *p < 0.05 compared to sham; #p < 0.05 compared to vehicle. N = 6/per group.
Total rest time showed significant group [F(4,25) = 24.67, P < 0.0001] effects. All of the animals with stroke, regardless of treatment, had longer rest times than the intact animals (p < 0.05). P4 alone, CQ alone and P4+CQ showed increases in locomotor activity compared to injured vehicle-treated animals (p < 0.05), indicating that given alone or in combination, the treatments produced better outcomes on this measure (Fig. 2B).
There was a significant group [F(4,25) = 68.68, P < 0.0001] effect. A significant reduction in grip strength was observed in all injured groups compared to shams at 3 days post-stroke (p < 0.05). P4 alone, CQ alone and P4+CQ showed significantly improved grip strength compared to vehicle (p < 0.05) (Fig. 2C) but the treatment groups were not different from one another.
P4 and CQ effects on autophagy by modulating LC3 and SQSTM1/p62
We observed a significant increase in the LC3-II/LC3-I ratio in the vehicle and CQ groups compared to shams (p < 0.05). P4 alone and CQ alone exerted opposite effects on the LC3-II/LC3-I ratio relative to vehicle (p < 0.05), while the combination produced a significant increase in the ratio compared to P4 alone (p < 0.05) (Fig. 3A). We also observed a decrease in SQSTM1/p62 (sequestosome 1) expression in the vehicle, P4-alone and CQ-alone groups compared to shams (p < 0.05), whereas the combination showed a significant increase in SQSTM1/p62 expression compared to vehicle, P4-alone, and CQ alone (p < 0.05) (Fig. 3B).
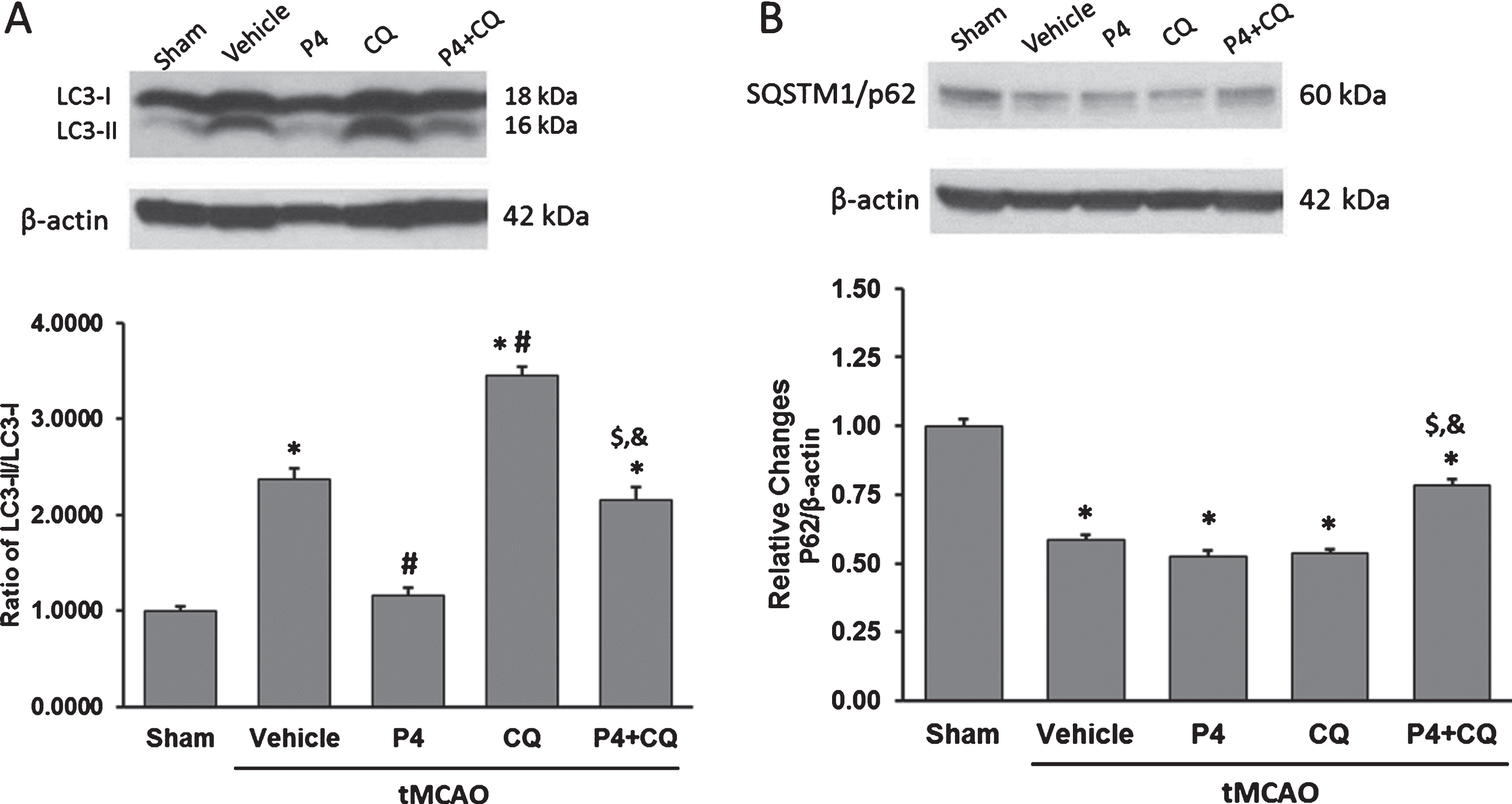
Effect of P4 and CQ alone or in combination on expression of (A) LC3 and (B) SQSTM1/p62 in rat brain 24 h post-surgery. β-actin was used as the loading control. Values are expressed as Mean±SE. *p < 0.05 compared to sham; #p < 0.05 compared to vehicle; $p < 0.05 compared to P4; &p < 0.05 compared to CQ. N = 5/per group.
In the vehicle group, there was a significant decrease in Bcl-2 (Fig. 4A) and a significant increase in Bax (Fig. 4B) expression compared to shams (p < 0.05). P4 alone and CQ alone produced significant but moderate increases in Bcl-2 expression and reductions in Bax expression compared to vehicle (p < 0.05). The combination of P4 and CQ produced a significantly greater increase in Bcl-2 and reduction in Bax expression compared to P4 or CQ given alone (p < 0.05) (Fig. 4A)
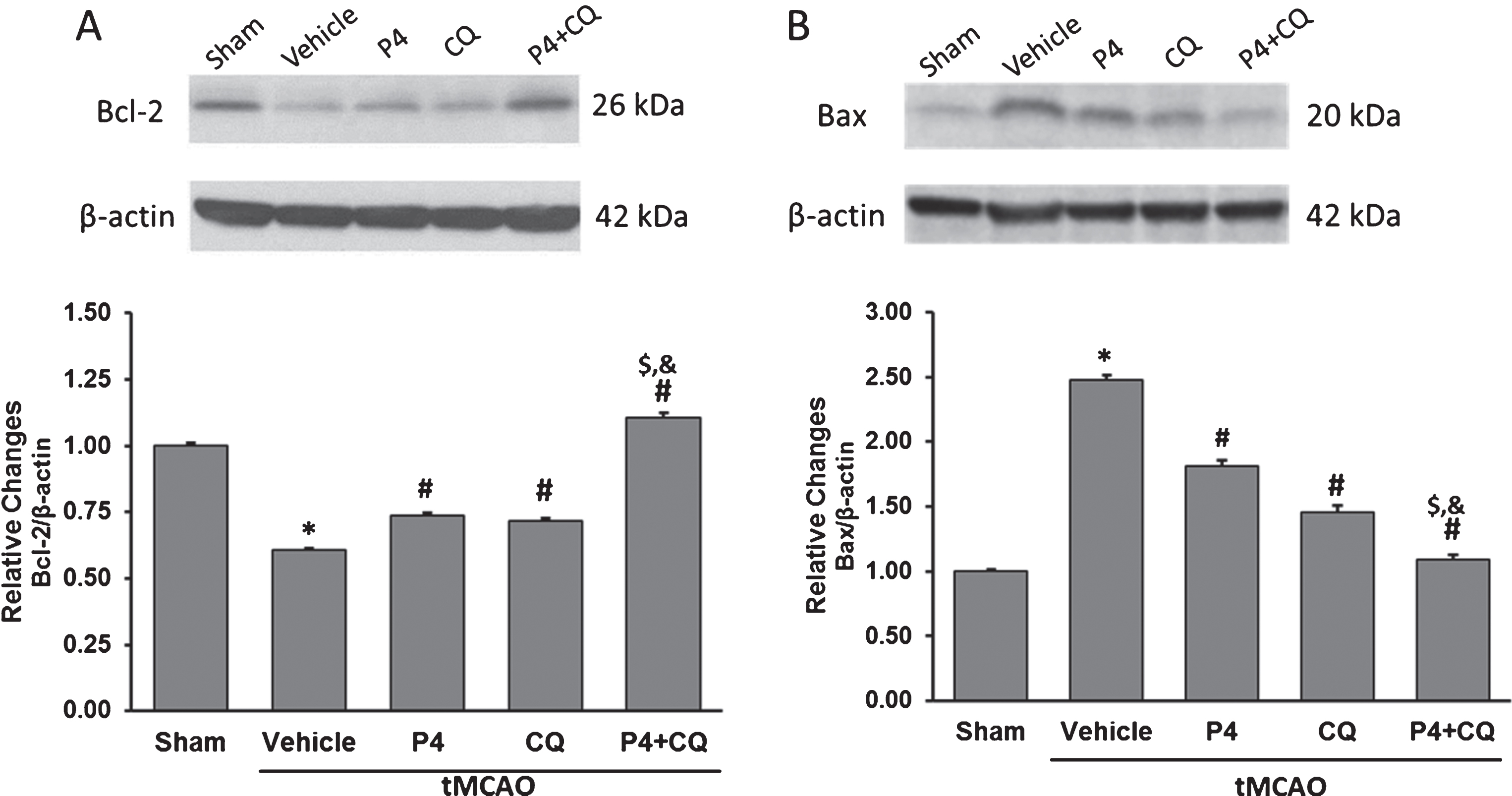
Effect of P4 and CQ on expression of (A) Bcl-2 and (B) Bax in rat brain 24 h post-surgery. β-actin was used as the loading control. Values are expressed as Mean±SE. *p < 0.05 compared to sham; #p < 0.05 compared to vehicle; $p < 0.05 compared to P4; &p < 0.05 compared to CQ. N = 5/per group.
In this proof-of-concept study, we evaluated the potential neuroprotective effects of combining P4 and CQ treatment in male rats following tMCAO to see if better modulation of autophagy would improve the outcome of this injury. Both P4 alone and CQ alone were beneficial in restoring grip strength and locomotion and reducing infarct volume after stroke. P4 combined with CQ resulted in more significant reductions in infarct volume than either drug given alone, but the combination did not lead to better functional outcomes measured at 3 days post-stroke.
The benefit of P4 treatment given before reperfusion in restoring behavioral function and reducing infarct volume following tMCAO was consistent with previous reports (Atif et al., 2013; Yousuf, Atif et al., 2014; Yousuf, Sayeed et al., 2014) from our laboratory. In the present study, treatment with CQ alone also helped to attenuate some of the functional deficits and reduced cortical infarct size, probably through different signaling pathways from P4.
Both P4 and CQ given independently can modulate a number of physiological mechanisms involved in stroke recovery (Atif et al., 2013; Cui et al., 2013; Thome et al., 2013; Yousuf, Sayeed et al., 2014). To explore some of these mechanisms, we investigated the individual and combined effects of P4 and CQ on the expression of autophagy- (LC3, SQSTM1/ p62) and apoptosis- (Bcl-2, Bax) related markers at 24 h post-occlusion.
As a self-digestion process in eukaryotic cells, autophagy is a form of type II programmed cell death (Levine & Klionsky, 2004). Recent studies have reported that, after cerebral ischemia, autophagy is activated and contributes to neuronal/astrocytic cell death (Qin et al., 2010; Wen et al., 2008), and that inhibition of autophagy can attenuate cerebral ischemia-reperfusion injury (Dong et al., 2017; Shui et al., 2016). LC3 is an autophagosomal ortholog of yeast Atg8, and is required for the formation of membranes of autophagosomes (Shui et al., 2016). The cytoplasmic form of LC3, LC3-I, is distributed in the cytoplasm diffusely, but modified and concentrated in autophagosomes during autophagic activation (Kabeya et al., 2000; Mann & Hammarback, 1994; Mizushima, 2004). LC3-II is considered a marker of autophagosomes in mammalian cells (Kabeya et al., 2000), and inhibition of autophagy is associated with the accumulation of p62 protein (Mathew et al., 2009). Autophagic flux can be indirectly measured by examining the LC3-II/I ratio and p62 levels. As shown in Fig. 3A-B, an increase in the ratio of LC3-II/LC3-I and a decline in p62 expression in our vehicle group suggest that at least some autophagy is activated early after tMCAO.
We found that dosing with P4 decreased the expression of LC3-II in the ischemic cortex in the first 24 h after ischemia. CQ, which inhibits the later step of autophagy, leads to accumulation of LC3-II by preventing the degradation of LC3-II-containing autolysosomes (Pestana et al., 2015). In our study, p62 expression was not significantly influenced by P4 or CQ alone but was up-regulated by the combination, suggesting that the combination could have a greater effect on inhibiting autophagy after a stroke, leading to a smaller stroke infarct.
To probe the effect of P4 and CQ on apoptotic pathways, two members of the B-cell lymphoma class proteins were measured: Bcl-2 (anti-apoptotic) and Bax (pro-apoptotic). We observed that in our stroke model P4 up-regulated the expression of Bcl-2 protein and suppressed that of Bax, findings in agreement with other reports that P4 enhances Bcl-2 expression and decreases Bax level in different injury models (Atif et al., 2013; Yao et al., 2005). CQ and P4 had the same influence on Bcl-2 and Bax expression, and combination of P4+CQ strongly inhibited apoptosis by modulating Bcl-2 and Bax expression better than P4 or CQ alone. Although combinational effects were not demonstrated on our measures of autophagy, the data on apoptosis lead us to think that P4 and CQ neuroprotection could act partly through inhibition of Bax and enhancement of pro-survival Bcl-2, and that combining the two drugs could have salutary effects on this process. Ischemic stroke induces multiple injury pathways, including autophagy and apoptosis, leading to loss of brain tissue. On some measures of damage, a combination of P4 and CQ could help to protect the brain from ischemic damage better than vehicle or either drug alone, and this could lead to greater functional recovery in the chronic stage of the disease. Our combination, in the doses employed here, did not have a significant effect on the autophagic reaction to stroke but it did reduce apoptosis by modulating the expression of apoptosis-related proteins, leading to more rapid functional recovery associated with smaller infarct size.
With respect to the functional outcomes we observed at 3 days post-stroke, it is possible that the functional measures used in our experiment were not sensitive enough to discern treatment-induced behavioral differences so soon after a short course of treatment, or after looking at behavior at such a short time after the stroke event. Even more critically, the measures we used may not have been sensitive enough to be altered by the combination therapy, despite the decrease in the size of the infarct. Our results might have been different if we had waited a longer time after injury to see whether the test treatments produced outcomes that were not seen in the vehicle controls. Frankly, we expected to see more salutary early effects of the combination therapy and therefore did not plan for testing more complex cognitive and fine sensory and motor deficits, which might have revealed more subtle changes in behavior in the acute stage of the injury. These uncertainties call for further investigation (Soros et al., 2017). Finally, the concept of very late recovery after brain injury is not a novel notion and has well-documented historical validation. As but one example, in 1974, Norman Geschwind noted that “true neurological changes after apparently fixed lesions, occur at every level of the nervous system in man and the time course of these late changes may extend in some cases over many years” (Geschwind, 1974, p. 472). Geschwind’s chapter on late changes in the nervous system can be considered a classic in describing the long course of recovery, which is very much influenced by individual variation.
Our proof-of-principle study can be interpreted to suggest that therapy with the right combination of P4 and CQ could have a neuroprotective effect on some components of a transient ischemic injury, most notably reduction of infarct size. Since current neuroprotective treatment options for stroke remain very limited, our finding may warrant further exploration for its potential translational therapeutic effects. Future studies should determine the best combinatorial doses given over a longer period of time, examine more complex behavioral and cognitive outcomes measured in both the acute and chronic stages of the injury, and determine whether the beneficial effects are different in males and females, and in females at different times in their hormonal cycling.
Footnotes
Acknowledgments
The first author was sponsored by the Jiangsu Provincial Department of Education. A portion of this research was subsidized by the Marcus Foundation and Allen and Company, USA. The primary support for this research was provided by the University Natural Science Research Project of Jiangsu Province (Grant No. 17KJD310003) and the Research Foundation of Jiangsu Health Vocational College (Grant No. JKA201707). The authors thank Leslie McCann, Fahim Atif, Claudia Espinosa-Garcia, Elena G. Sergeeva, Nefize Turan, Bushra Wali, and Seema Yousuf for their invaluable assistance and advice.