
Abstract
Background:
In this study, we evaluate the role of RGMa (Repulsive Guidance Molecule a) during peripheral nerve regeneration using the mouse median nerve model.
Methods:
By real-time PCR and Western Blot analysis, we examined expression changes of RGMa mRNA and RGMa protein in neural tissue after transection and microsurgical repair of the mouse median nerve distal to the transection site. We evaluated histomorphometrical changes in neural tissue distal to the injury site and functional recovery of the grasping force after median nerve transection and repair in wild-type mice and RGMa+/– heterozygous mice.
Results:
RT-PCR revealed a 1,8 fold increase of RGMa mRNA two weeks and a 4,4 fold increase of RGMa mRNA 3 weeks after nerve transection and repair in the nerve segment distal to the injury site. In Western blot analysis, we could show a high increase of RGMa in the nerve segment distal to the injury site at day 14. Histomorphometrical analysis showed significant differences between wild-type animals and heterozygous animals. The absolute number of myelinated fibres was significantly higher in operated heterozygous RGMa+/– animals compared to operated wildtye animals. Using the functional grasping test, we could demonstrate that peripheral nerve regeneration is significantly diminished in heterozygous RGMa+/– mice.
Conclusions:
Employing the mouse median nerve model in transgenic animals, we demonstrate that RGMa plays an important role during peripheral nerve regeneration.
Keywords
Background
RGMa (Repulsive Guidance Molecule A) was originally identified as an axon repellent in the chick retinotectal system (Monnier PP 2002, Stahl 1990). The receptor for RGMa is Neogenin and is widely expressed in embryonic and adult tissues (De Vries 2008, Mueller 2006). By binding to its receptor, RGMa plays an important role in axon guidance, neuronal differentiation and survival during the developement of the central nervous system (CNS) (Matsunaga 2004, Rajagopalan 2004, Wilson 2006). In the CNS RGMa ist upregulated after ischemic stroke and spinal cord injury (Schwab 2005, Schwab 2005). RGMa is also present in multiple sclerosis patients’ active and chronic lesions and in amyloid plaques of patients diagnosed with Alzheimer’s disease (Demicheva 2015, Satoh 2013). Furthermore, RGMa is a strong inhibitor of angiogenesis (Fujita 2017). Recently it has been shown that inhibiton of RGMa promotes regeneration and repair after spinal cord injury in rats (Mothe 2017). Less is known about the role of RGMa in the peripheral nervous system (PNS). In this study, we evaluated the role of RGMa using the mouse median nerve model for assessment of peripheral nerve regeneration.
Methods
Animals and surgical procedure
All surgery was performed using an anesthetic combination of fentanyl (0,05 mg/kg), midazolam (5 mg/kg) and medetomidin (0,5 mg/kg). National guidelines for laboratory animal care and safety were followed.
To assess in vivo responses to peripheral nerve injury, 48 adult male wild-type (WT) C57BL/6 mice weighing approximately 30 g underwent transection of the left median nerve (Tos 2008, Jaminet 2013) directly after the branching of a constant sensible nerve running exactly parallel to the M. pectoralis muscle border. The nerve was subsequently repaired at 40×magnification using two epineural stitches of 12–0 monofilament nylon (Figs. 1 and 2) (Jaminet 2013). Mice were sacrificed after 0, 7, 14 and 21 days postlesion for RT-PCR (n = 6) and Western blot analysis (n = 6).
Intraoperative view of the murine axilla with the three main upper extremity nerves and the pectoralis muscle before transection of the median nerve (15). Intraoperative view showing the transection site of the median nerve (black arrow) and the analyzed nerve segment (dashed arrow). The constant sensible nerve running parallel to the M. pectoralis border has been marked by a white arrow.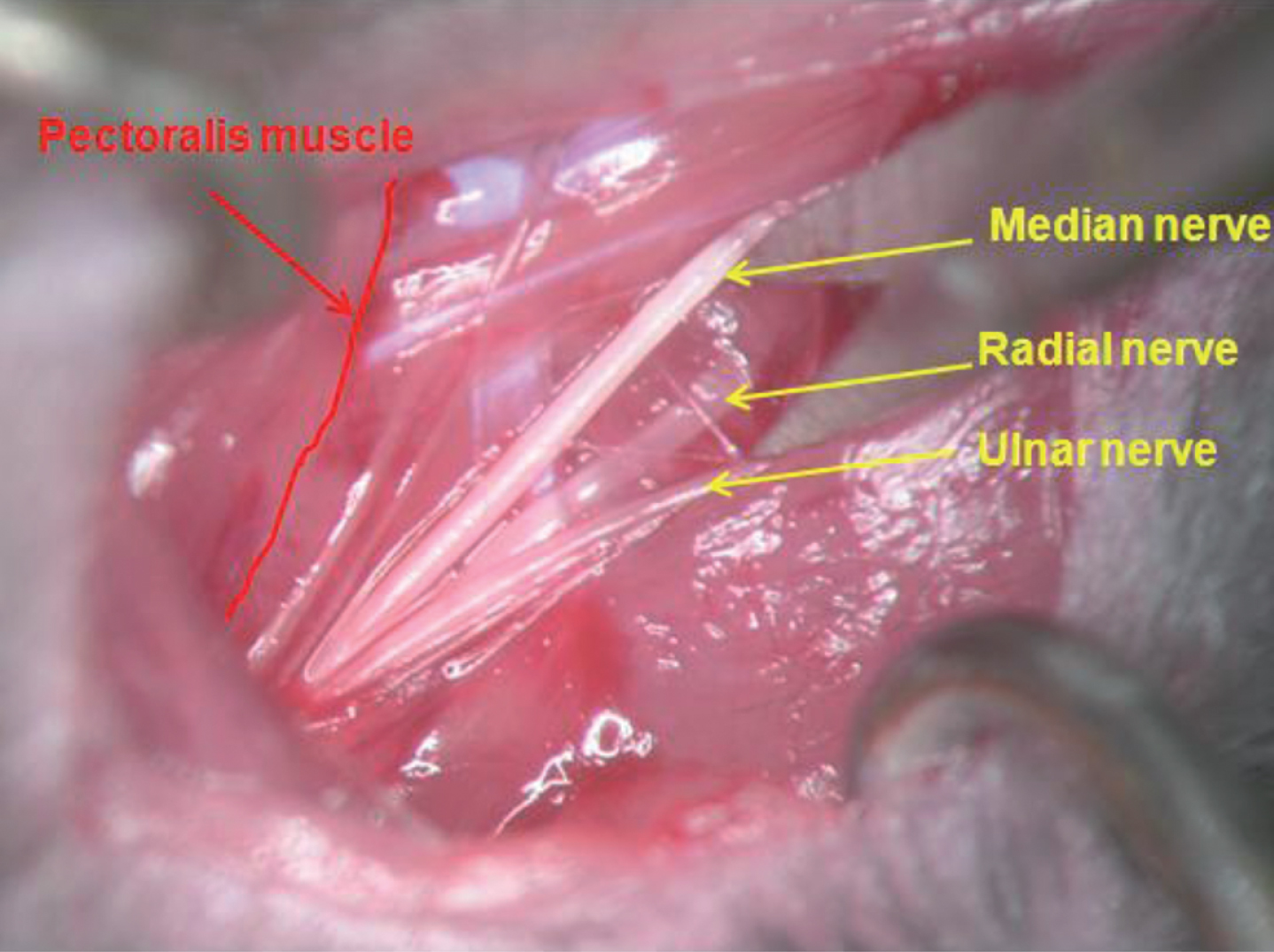

To assess functional recovery after nerve transection and quantitative morphology of nerve fiber regeneration, a total of 48 male mice were used: 24 WT mice and 24 RGMa+/– heterozygous mice (Tessier-Lavigne, Stanford, USA) (Niederkofler 2004). The mice were divided in 4 groups. 12 WT animals and 12 RGMa+/– heterozygous animals underwent transection and repair of the median nerve as described above. 12 WT animals and 12 RGMa+/– heterozygous animals only underwent operative exposure of the median nerve without transection (sham) (Table 1). Similar to the procedure used for rats (Papalia 2003), in order to prevent interferences with the grasping test device during testing due to the use of the contra-lateral forepaw, the contra-lateral median nerve was transected at the middle third of the brachium and its proximal stump was sutured in the pectoralis major muscle to avoid spontaneous reinnervation.
Description of the experimental groups (functional recovery and histomorphometry)
Total RNA was extracted as described by the manufacturer (peqGOLD TriFast, PEQLAB, Erlangen, Germany) from explanted median nerves distal to the transection site. RNA was reverse-transcribed into cDNA in a 20μl reaction (iScripttrademark cDNA Synthesis Kit, BioRad) and subjected to polymerase chain reaction (PCR). Semiquantitative analysis was performed using real-time PCR (RT-PCR) (iCycler; Bio-Rad Laboratories Inc., Hercules, CA, USA) to examine murine RGMa expression levels in median nerve distal to the transection site at 7, 14 and 21 days after transection. Primer sets contained 10 pM of each, sense primer 5′-CAG CAA GCT CAC CAT CAT CT-3′ and antisense primer 5′-CCC GAC ACC TTC TCT GTG AT-3′. The primer set was amplified using increasing numbers of cycles of 94°C for 1 min, 60°C for 2 min, 72°C for 4 min, and a final extension of 72°C for 7 min. Samples were controlled for GAPDH using the following primers: sense 5′-CGG CCT CAC CCC ATT TG-3′, antisense 5′-CGA GAA TGG GAA GCT TGT CAT C-3′.
Western blot analysis
The explanted median nerves distal to the transection site (n = 6) were lyzed and homogenized. The lyzed nerves were pooled and eluted in RIPA buffer (50 mM Tris-HCl [pH 7,4], 150 mM NaCl, 0,5% deoxycholic acid, 0,5% Triton X-100, 1 mM PMSF, 1 mM sodium orthovanadate, 1×protease inhibitor mixture (Roche Molecular Biochemicals)). An equal volume of proteins was loaded on 10% SDS containing polyacrylamide gels for electrophoresis and transferred to a PVDF membrane (Bio-Rad, Hercules, U.S.). The membrane was blocked with 5% nonfat milk, Tris-buffered saline containing 0,05% Tween 20 for 1 hour, then immunoprobed with anti-RGMa antibody (Abcam ab26287, Cambridge, GB) overnight at 4°C. After three washes, the membrane was incubated with HRP-conjugated secondary antibody for 1 h at room temperature. After 3 more washes, the reaction was visualized with the enhanced chemiluminescence reaction (ECL Detection system, Fluka-Luminol 09253, p-Coumeric acid; Sigma-Aldrich, Munich, Germany). As housekeeping protein GAPDH was used and detected by anti-GAPDH antibody (sc25778, Santa Cruz, Santa Cruz, U.S.). Normalized relative intensity was determined using ImageJ V 1.43 Software.
Resin embedding for quantitative morphology of nerve fiber regeneration
At the end-point of regeneration (day 50), operated WT and RGMa+/– heterozygous mice (genetic background C57BL/6) and their respective sham controls (n = 5 per group) were fixed by transcardial perfusion (fixation solution, containing 2.5% purified glutaraldehyde and 0.5% saccarose in 0.1 M Sorensen phosphate buffer) and an 8 mm long segment of the median nerve distal to the site of lesion was removed. A 4/0 stitch was used to mark the proximal stump of the nerve segment. An 8 mm long segment of uninjured median nerve, taken from a corresponding level, was withdrawn from non-operated control animals and used as control. The nerve samples were further fixed and prepared for design-based quantitative morphology of myelinated nerve fibers by immediate immersion in the fixation solution for 3–4 h. They were then washed in a solution containing 1.5% saccarose in 0.1M Sorensen phosphate buffer, post-fixed in 1% osmium tetroxide, dehydrated and embedded in Glauerts’ embedding mixture of resins consisting in equal parts of Araldite M and the Araldite Härter, HY 964 (Merck, Darmstad, Germany), to which was added 1–2% of the accelerator 964, DY 064 (Merck, Darmstad, Germany) and 0,5 % of the plasticizer dibutyl phthalate. Series of 2μm thick semi-thin transverse sections were cut starting from the distal stump of the median nerve segment, using a Ultracut UCT ultramicrotome (Leica Microsystems, Wetzlar, Germany), i.e. 8 mm distal to the transection site. They were stained by toluidine blue for high-resolution light microscopy examination and design-based quantitative morphology that for each nerve specimen was carried out according to the stereological method described by Geuna et al (Geuna 2000, 2001, 2004, 2005). The stereological method, which consists of a two-dimensional variation of the classical disector procedure (two-dimensional disector), and the sampling scheme are verified by comparing estimates with the true density and total number of myelinated fibers in peripheral nerve trunks where true values have been accurately determined by extensive counting. The verification of the 2-D disector method, both of normal and regenerated nerves, showed that estimates of density and total number of myelinated nerve fibers are unbiased.
Functional grasping test
Starting from day 5 postoperative, the grasping test was carried out every 5 days until day 50. Grasping test was performed using a precision balance connected to a rod. The test was performed as described by Tos (Tos 2008). The mouse was held by the tail and put to the metal bar until it is close enough to the device to grasp it. The mouse then pulls on the bar until it looses the grip. Each mouse was tested three times and the average value was recorded. Operated WT mice (n = 12) were compared to operated RGMa+/– mice (genetic background C57BL/6; n = 12). Non-operated WT (sham) (n = 12) and non-operated RGMa+/– (sham) (n = 12) mice served as controls.
Statistical analysis
All data are presented as mean±SD. Statistical analysis was performed using one-way analysis of variance (ANOVA) test for histomorphometrical data. The different time-point assessments of the grasping test were compared by analysis of covariance (ANCOVA). A probability value of less than 5% was considered to be statistically significant.
Results
RGMa mRNA is induced in the injured median nerve distal to the transection site (WT animals)
In order to evaluate the possible role of RGMa during peripheral nerve regeneration, we examined time-dependent mRNA expression after median nerve injury in WT animals. RT-PCR revealed a 1,8 fold increase of RGMa mRNA two weeks after nerve transection and repair and a 4,4 fold increase of RGMa mRNA three weeks after nerve transection and repair (Fig. 3) in the nerve segment distal to the injury site.
RT-PCR revealed a 1,8 fold increase of RGMa mRNA two weeks after nerve transection and repair and a 4,4 fold increase of RGMa mRNA three weeks after nerve transection and repair in WT animals. Data are shown as Mean±SEM, n = 6; ***P < 0.001.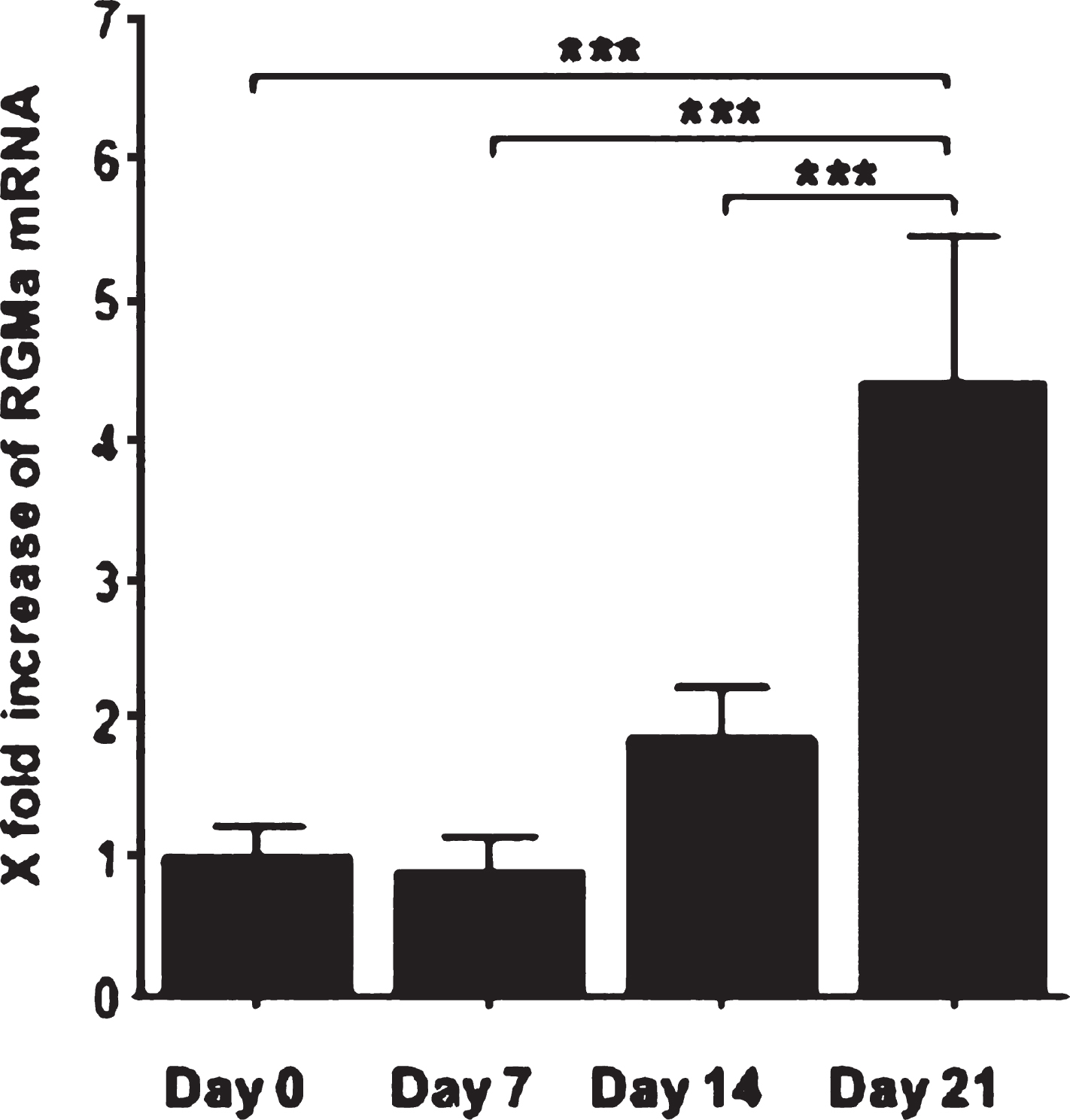
We examined protein expression changes of RGMa after median nerve injury to further elicit the role of RGMa in WT animals. In Western blot analysis, we could show a highly specific increase of RGMa in the nerve segment distal to the injury site at day 14 after peripheral nerve transection and repair in WT animals. Before and after day 14, RGMa is expressed at lower but still high levels (Fig. 4) compared to the uninjured nerve. The normalized relative intensity of protein expression is presented in Fig. 5.

Western Blot: High RGMa protein expression at day 14 distal to the transection site. (RGMa 50 kD; GAPDH 37 kD).

Western Blot: High RGMa protein expression at day 14 distal to the transection site.
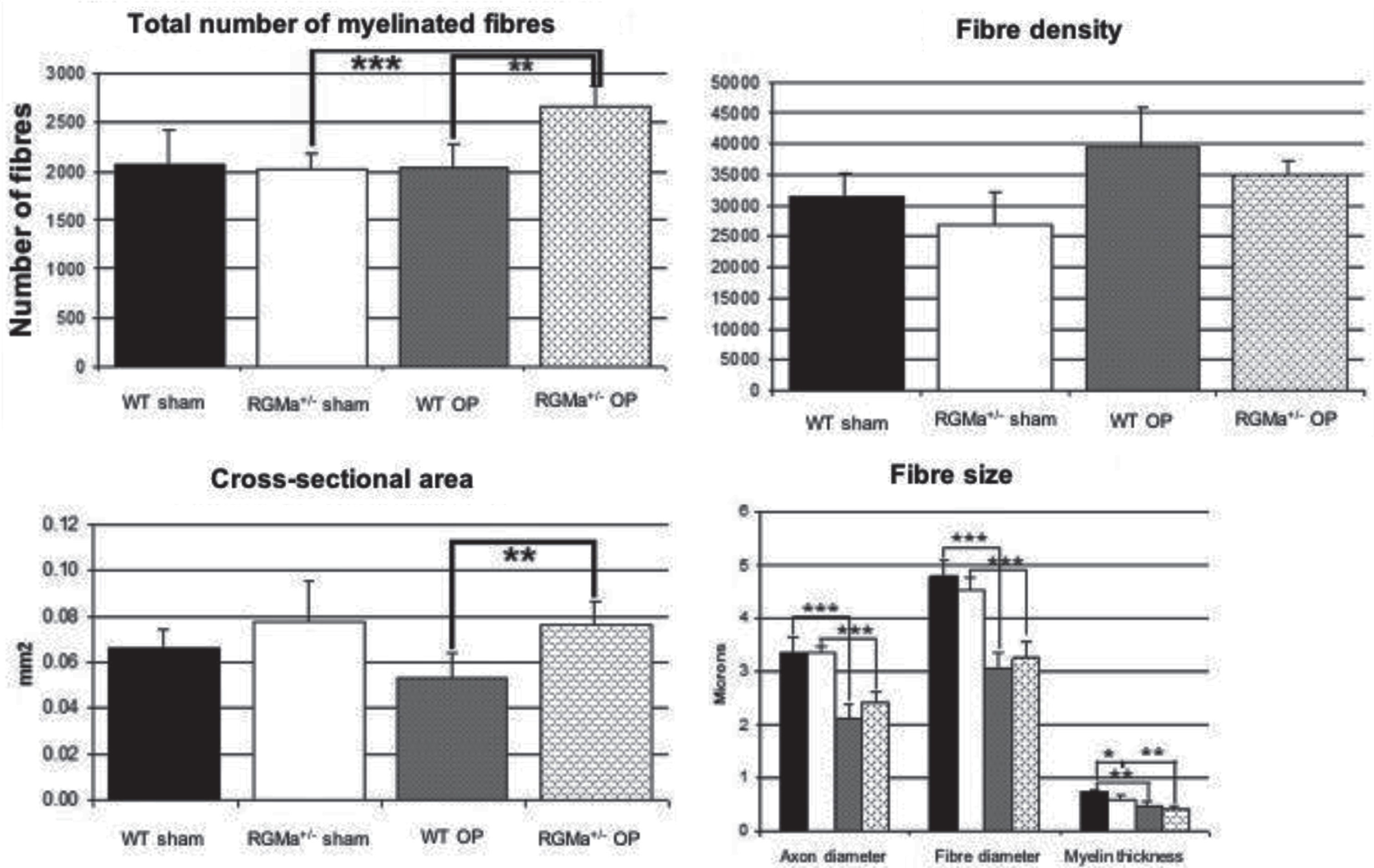
Various morphological predictors of nerve regeneration were assessed by stereology at day 50 after nerve transection. Regarding total number of myelinated fibers, we could show a significant increase after nerve transection and repair in heterozygous RGMa+/– animals with nerve transection and repair. The cross-sectional areas of the nerves also showed a significant increase, comparable to the total number of myelinated fibres in animals with nerve transection and repair. No significant difference could be shown regarding fibre density between all 4 groups. Axon diameter, fiber diameter and myelin thickness were significantly diminished in wildtype animals and transgenic animals after transection and repair without any difference between the two groups (Figs. 6 and 7).
Histomorphometry: significant increase in total number of fibers and cross-sectional area in heterozygous mice compared to WT mice with nerve transection. Histomorphometry: significant increase in total number of fibers and cross-sectional area in heterozygous mice compared to WT mice with nerve transection.
Using the grasping test, we could show that peripheral nerve regeneration is significantly diminished in heterozygous RGMa+/– mice. Whereas there is no difference in grip strength in both control groups (sham WT and sham RGMa+/– mice), we could demonstrate a tremendous drop in total grip strength in operated RGMa+/– animals compared to operated WT animals. On the one hand, grip strength of heterozygous mice only reaches 50% of the strength of WT animals. Furthermore, they never get back their preoperative original strength. On the other hand, the functional recovery of operated RGMa+/– mice moves on by far more slowly compared to WT animals (Fig. 8).
Functional grasping test: grip strength of heterozygous mice only reaches 50% of the strength of WT animals. Functional recovery of operated RGMa+/– mice moves on by far more slowly compared to WT animals, never reaching back the original force.
RGMa was first identified as the cue responsible for mapping temporal retinal axons onto the posterior region of the chick tectum (Monnier 2002). The netrin receptor Neogenin was identified as the guidance receptor mediating RGMa chemorepulsion (Rajagopalan 2004). Neogenin is a bifunctional guidance receptor able to promote RGMa-mediated repulsion as well as chemoattraction in response to Netrin-1 (De Vries 2008, Wilson 2006, Keeling 1997, O’Leary 2013).
In this article, we show that RGMa plays an important role during peripheral nerve regeneration. Findings of our study identify RGMa as an important factor for axonal regeneration after peripheral nerve injury. We demonstrate that RGMa mRNA is highly increased in the nerve segment distal to the injury site during the early phase of peripheral nerve regeneration at weeks two and three. Parallel to these results we show a peak of RGMa protein expression in the nerve segment distal to the injury site at week two. After this timepoint, RGMa protein expression drops down to the level of the uninjured nerve.
Using the grasping test, we showed that RGMa+/– mice recover their grasping force more slowly than WT mice and never get back their initial preoperative force. Histomorphometric results of regenerated nerve fibers showed a significant increase of myelinated fibres and cross-sectional area after nerve transection and repair in heterozygous RGMa+/– animals with nerve transection and repair. Even though the grasping force diminished, the total number of fibres increased, explained by a higher rate of axonal sprouting, promoted by the inhibition of RGMa. In the CNS, blocking of RGMa promotes regeneration and repair (Mothe 2017). In the peripheral nerval system (PNS), unguided sprouting increases the number of fibres, but due to the lack of guidance, these fibres do not reach the target muscle.
Furthermore Schwab et al. (Schwab 2005) showed a high increase of RGMa at the site of injury in the CNS and spinal cord. Invasion of RGMa presenting cells reaches its maximum at day 1–3. Later, it can still be found in the glial scar (Schwab 2005). In the PNS, scar tissue only develops when axon migration is not proceeding (Giger 2010). The lack of scar tissue and the later expression of RGMa (max. 2nd week) show that the expression pattern and function of RGMa in the PNS is completely different from the CNS.
Interestingly, the timepoint of maximal RGMa expression correlates with the peak of macrophage invasion in the distal nerve (Dubovy 2011). These cells could play a key role in peripheral nerve regeneration as strong RGMa presenters. Further studies are necessary to examine this potential interaction.
Summarizing, we explain the reduced grasping force an higher number of nerve fibres in heterozygous RGMa+/– mice by the fact that RGMa deficiency leads to a higher axonal sprouting rate and thus leading to less axons reaching the target muscle (Fig. 9). In WT animals, a subtile balance between its receptor Neogenin directs the growth cone distally to its target organ (Schäufele 2014).
Interpretation of the results: grasping force is reduced and total number of nerve fibres in heterozygous RGMa+/– mice is increased by the fact that RGMa deficiency leads to a higher axonal sprouting rate and thus leading to less axons reaching the target muscle.
Further studies are required to clarify the exact mechanism of action of RGMa during peripheral nerve regeneration. Future research has also to be focused on the distal neuromuscular junction site not analyzed in this study. An additional limitation of this study is the very small nerve size requiring advanced microsurgical skills for performing epineural suturing.